
Potential Value of Wood Tar as a Natural Fungicide against Valsa mali

Research and Development Center of Biorational Pesticides, Northwest A & F University, Yangling 712100, China
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(5), 1531; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051531
Submission received: 22 January 2022
/
Revised: 12 February 2022
/
Accepted: 14 February 2022
/
Published: 24 February 2022
(This article belongs to the Topic Bioactives and Ingredients from Agri-Food Wastes)
Abstract
:The Valsa canker caused by Valsa mali seriously harmed the production of East Asian apples and caused very significant economic losses. Considering the chemical residues and the improvement of people’s awareness of environmental protection, there is a need for screening new green pesticides for the control of Valsa canker. Therefore, we conducted systematic evaluations on the antifungal activity of wood tar. In this research, the effective concentration (EC50) of six strains of V. mali to wood tar was determined, and the EC50 ranged from 69.54 to 92.81 μg/mL. After treatment with wood tar, the hyphae of V. mali broke, swelled, and deformed; the permeability of the cell membrane increased; and the activity of pectinase reduced. Moreover, the expression levels of five genes related to pectinase also decreased significantly. In addition, the activities of phenylalanine ammonia lyase (PAL) and peroxidase (POD) of apple leaves treated with wood tar also increased. On detached apple branches, wood tar also showed therapeutic and protective activities. In the 2016–2019 field experiments, wood tar also showed good efficacy against Valsa canker and promoted the formation of callus. (In the experiments from 2016 to 2019, it can be seen that the control effect of 50% wood tar and 100% wood tar in the field is above 75% and promoted the formation of callus.) This study is the first to report the bidirectional efficacy of wood tar against Valsa mali and for trunk wound healing. The above results evidenced that wood tar has great potential to be developed as a natural alternative to commercial fungicides for the management of apple Valsa canker.
1. Introduction
Apple Valsa canker is a devastating disease caused by V. mali. In recent years, V. mali has seriously hindered the production of apples, with an average incidence of more than 50% in the Shaanxi Province of China, and the affected area is between 30% and 90%. V. mali infects branches, trunks, and fruit, causing necrosis of the diseased part, spreading to the entire trunk, and even causing the tree to the death in severe cases. It has caused great harm to apple production in East Asia, which is the main area of apple production, causing great economic losses to farmers. This pathogen can easily infect, especially when the tree is damaged or frostbite. Because the disease is occult and spreads rapidly, it is difficult to control the disease effectively [1].
In the past few decades, fungicides have been the main disease prevention and control agents. The asomate used on apple trees in the early years has been banned by the Ministry of Agriculture of China due to its high toxicity and high residue. According to recent reports, methionine acetic acid is recommended for the treatment of rot diseases, but it has not been registered [2]. Chemical agents have a limited ability to inhibit lesion expansion [3]. The risk of resistance also increases due to the high and excessive use of chemical fungicides [4]. When the fungicide is used for a long time, target pathogens easily develop resistance, and the iteration of the fungicide can not achieve the desired effect, so it is necessary to look for new alternatives in controlling the Valsa canker.
Wood tar is the byproduct that is discharged when wood is carbonized in a charcoal kiln or distilled in a distillation furnace. The application of wood tar is mostly limited to the scope of deep processing and mainly in the chemical industry, such as refining plasticizers and compounds used in chemical plants, which causes most of the content to be wasted. Although the antifungal activity of wood tar has been reported [5], its activity against V. mali is little known. The stickiness of wood tar is a fairly individual characteristic that can adhere to the lesion for a long time. Phenols are the main components of wood tar. Most of the phenols contained in it have bactericidal properties, such as phenol, 2-methoxyphenol, 2-methoxy-4-methylphenol, 2,6-dimethoxyphenol, and other reported antibacterial substances [6]. The synergistic action of various antibacterial compounds contained in wood tar can effectively slow down the occurrence of resistance. In addition, wood tar was natural and environmentally friendly.
Therefore, the purpose of this study is to: (a) evaluate the susceptibility of different samples of wood tar to V. mali, (b) explore the effect of wood tar on the morphological and physiological characteristics of V. mali, (c) estimate the protective and therapeutic activities of several tars on apple tree branches, and (d) assess the field efficacy of wood tar in controlling Valsa canker. These experiments will systematically evaluate the potential value of wood tar as a natural fungicide against V. mali and provide new information on understanding the mechanism of wood tar against V. mali.
2. Results
2.1. Sensitivity to Wood Tar
The EC50 values of wood tar inhibiting the mycelia growth of the six V. mali strains ranged from 69.54~92.81 μg/mL, with an average EC50 value of 81.16 ± 3.24 μg/mL. The EC50 value of carbendazim inhibiting mycelial growth ranged from 0.08 to 0.21 μg/mL, with an average EC50 value of 0.152 ± 0.019 μg/mL (Table 1).
2.2. Effect of Wood Tar on Mycelial Morphology
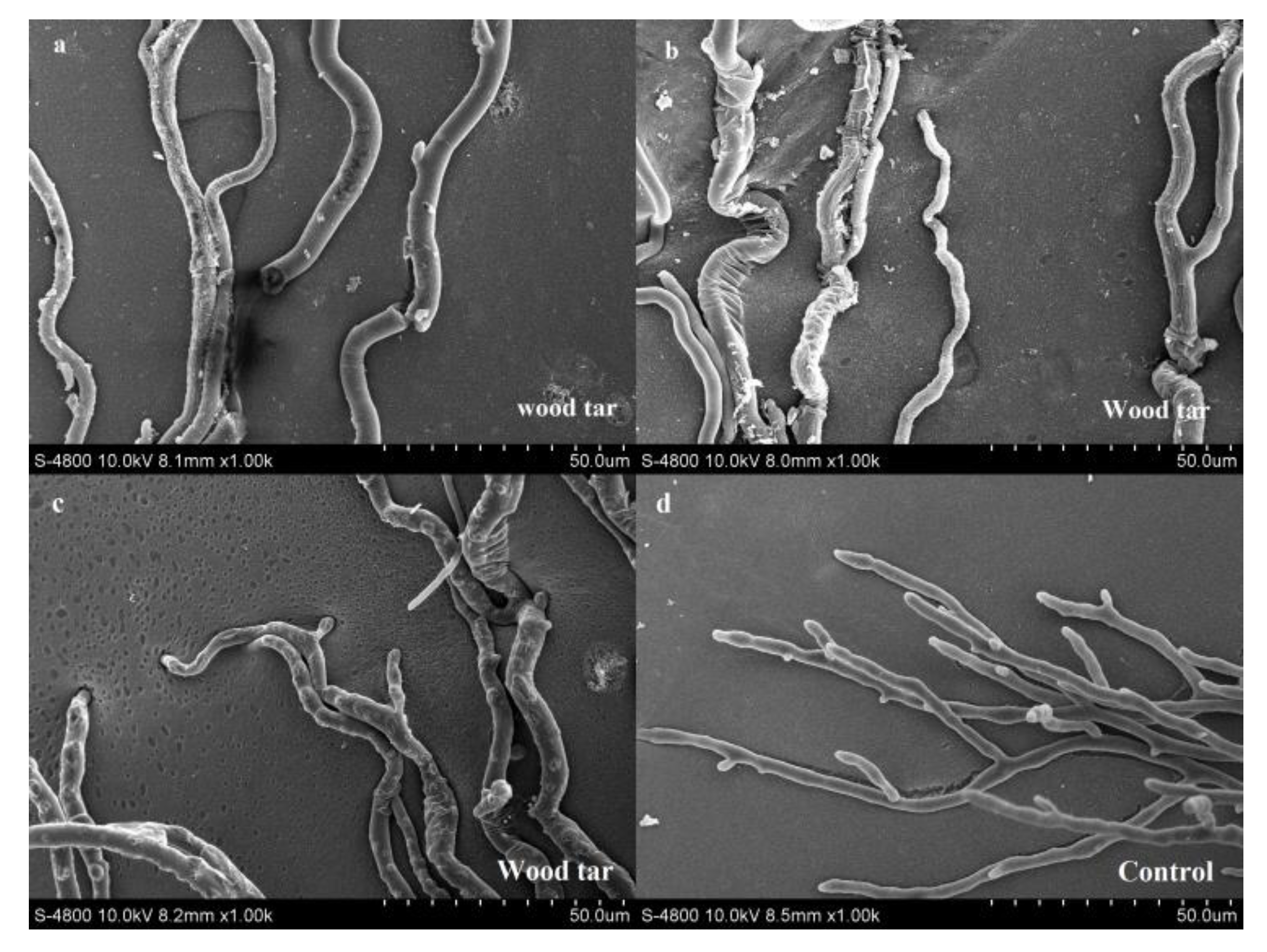
A scanning electron microscope was used to observe the mycelial morphology of V. mali treated or untreated with wood tar. After treatment with wood tar with a concentration of EC50, the mycelia were broken, swollen, wrinkled and twisted (Figure 1a–c). The mycelia in the control group were smooth and grew normally (Figure 1d).
2.3. Effect of Wood Tar on the Permeability of Cells
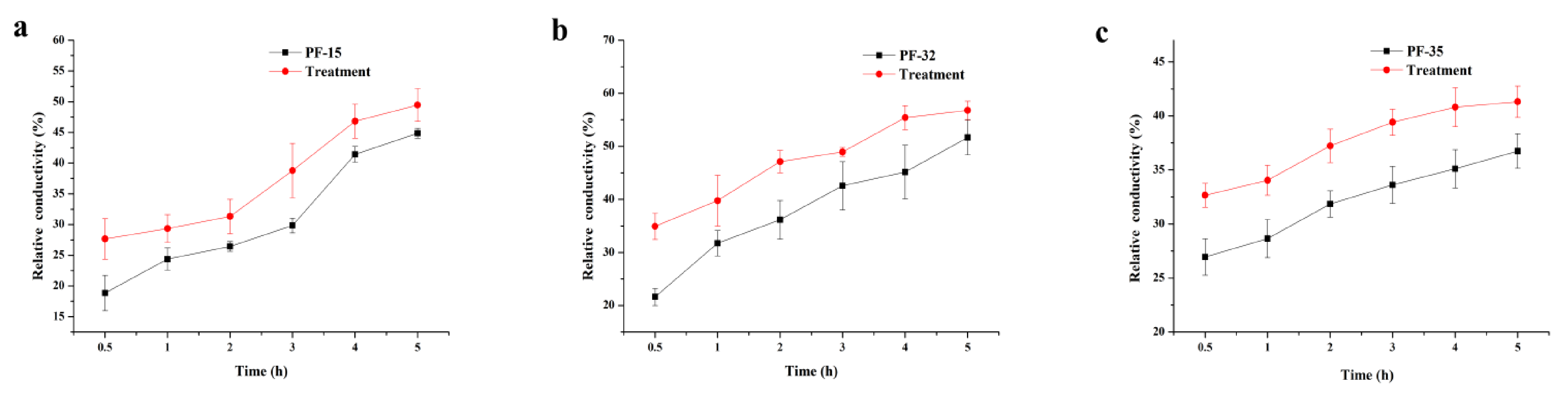
Whether with wood tar treatment or not, the relative conductivity of the strains increased over time. After wood tar treatment, the relative conductivity of the three strains was always higher than the corresponding untreated control (Figure 2).
2.4. Effect of Wood Tar on Pectinase Activity and Oxalic Acid Content
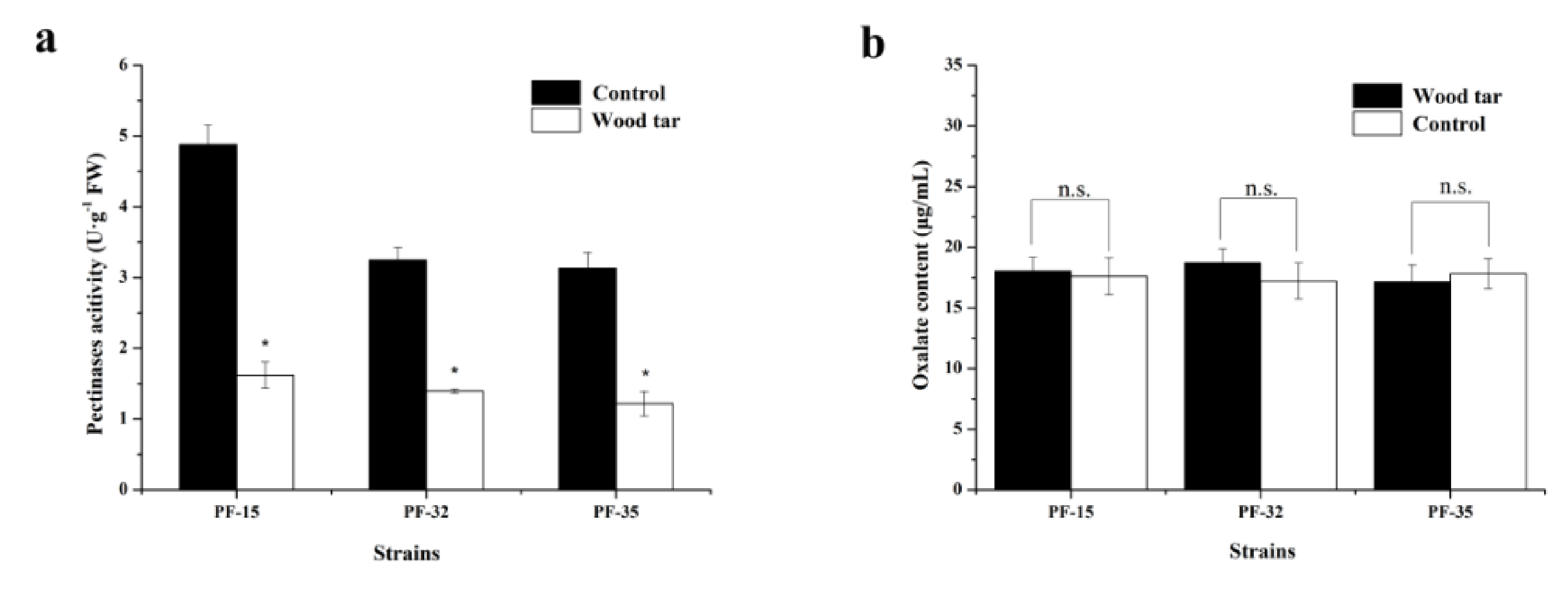
The activity of pectinase in the three strains differed from each other. After treatment with wood tar, the activity of pectinase significantly declined compared to the control (Figure 3a). However, there was no significant difference in the content of oxalic acid when treated with wood tar or not (Figure 3b).
2.5. The Expression of Pectinase Genes
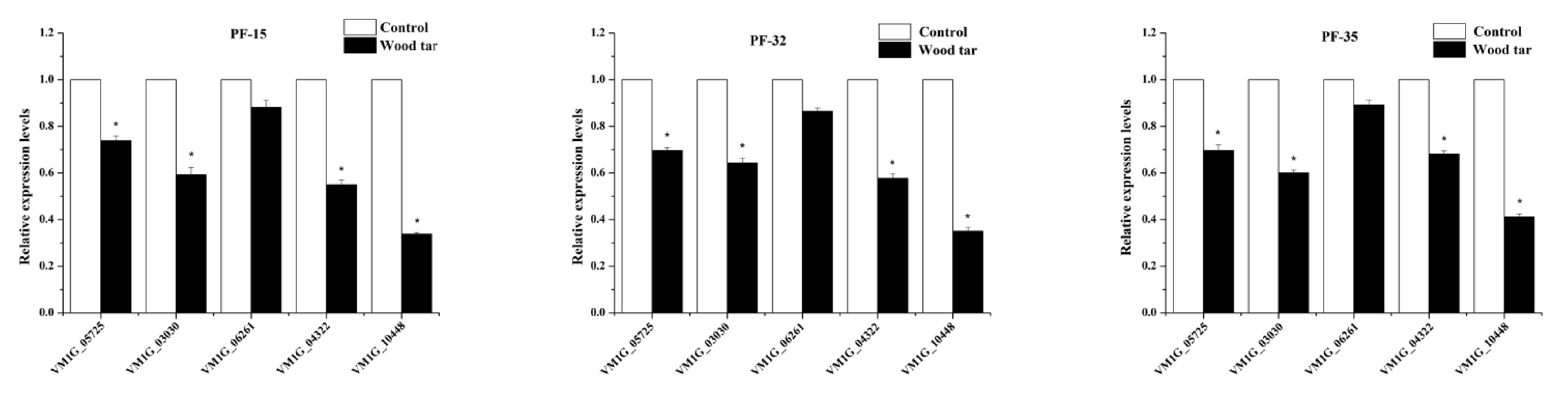
In this study, we examined the expression levels of five pectinase genes by q-RT-PCR [7]. As shown in Figure 4, four pectinase genes (VM1 G_05725, VM1 G_03030, VM1 G_04322, VM1 G_10448) were significantly down-regulated after wood tar treatment, while the gene (VM1 G_06261) had a slightly lower, but not significant, expression level after wood tar treatment (Figure 4).
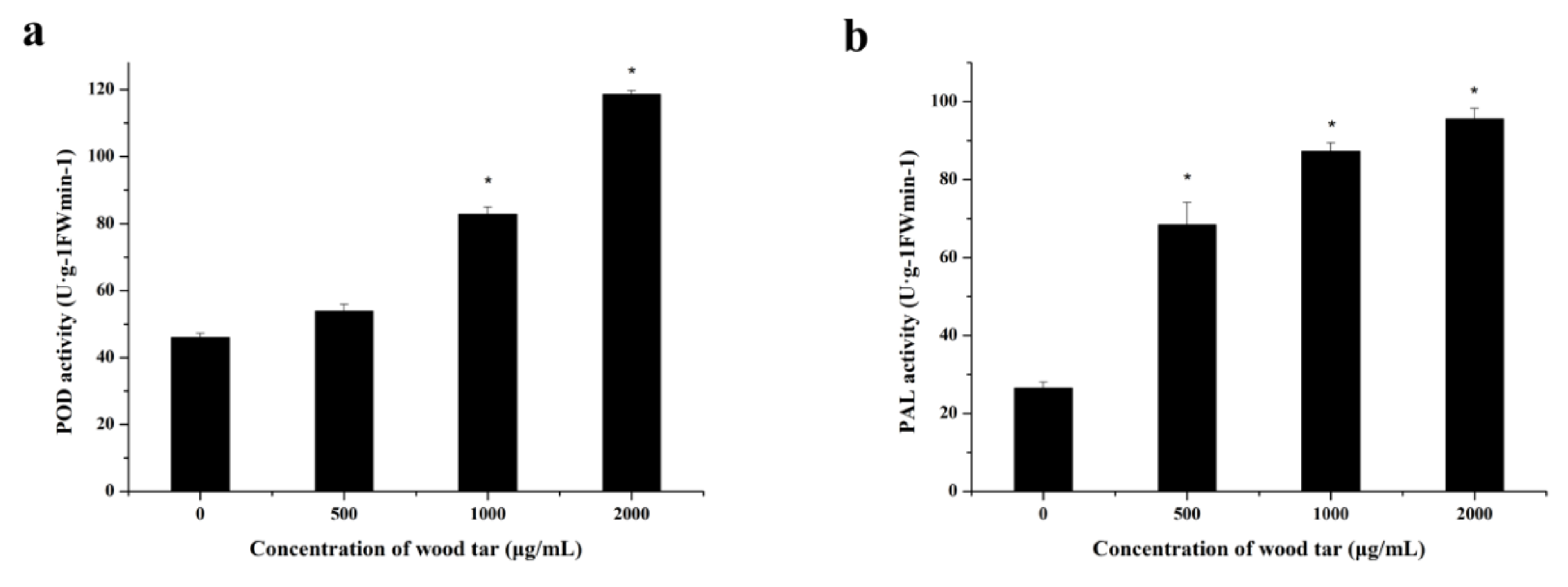
2.6. Activity of PAL and POD in Apple Leaves
As shown in Figure 5, the POD and PAL activities of apple leaves gradually enhanced with the increased concentration of wood tar.
2.7. Protective and Therapeutic Activity of Wood Tar on V. mali
After inoculation for 3 d on detached apple branches, a larger area of lesions was produced by the strain PF-32 and treated with water. When treated with 2000 μg/mL wood tar before inoculation, the protective efficacy reached 77.35%, which was comparable to that of carbendazim at 400 μg/mL (Table 2). When treated with 2000 μg/mL wood tar after inoculation, the curative effect was 67.89%. In addition, 1000 μg/mL wood tar had more than 45% protective and therapeutic activity against Valsa canker. These results showed that wood tar has both protective and therapeutic activity (Figure 6).
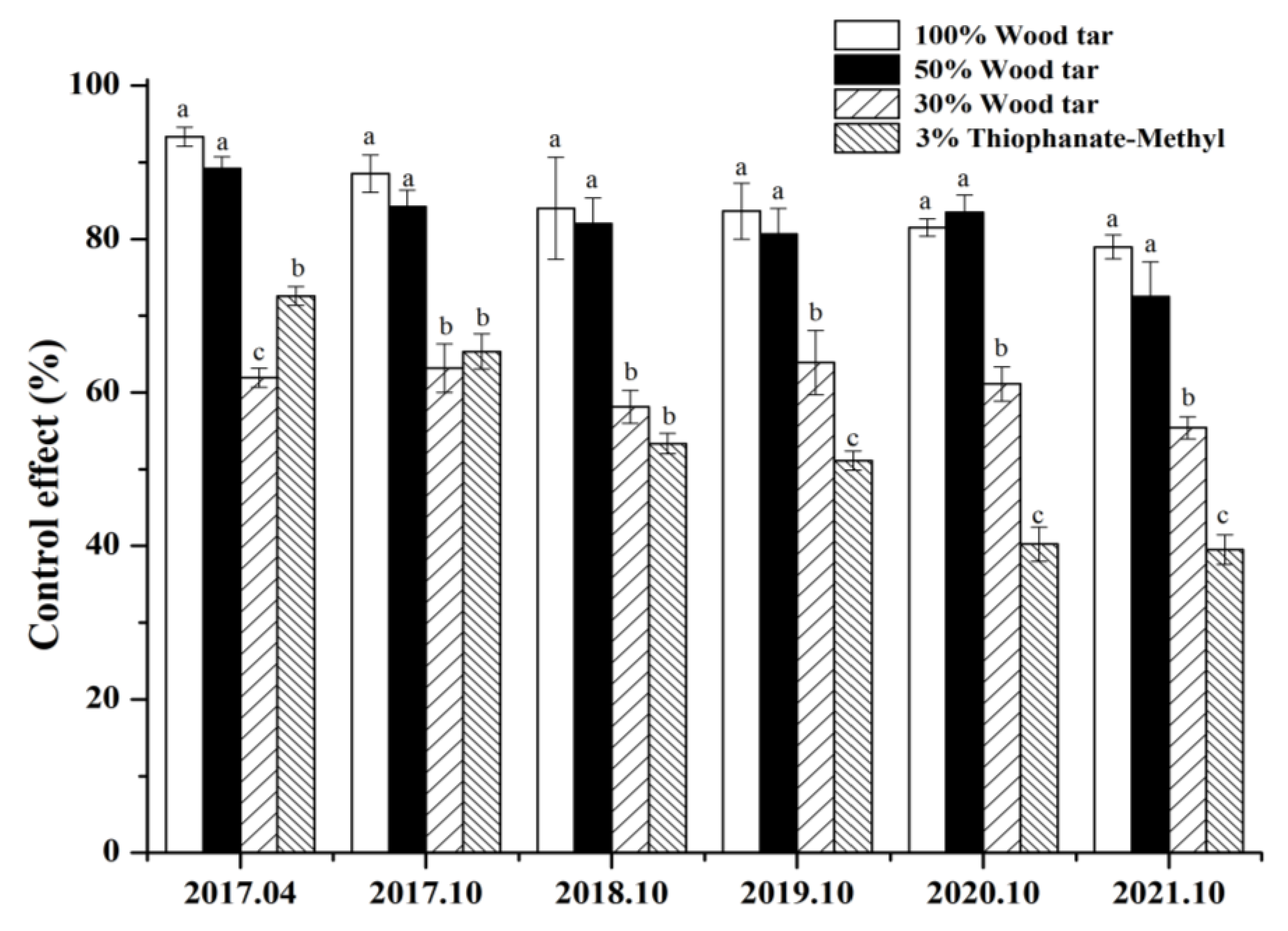
2.8. Field Test of Wood Tar on Valsa Canker
After being treated with 50% concentration, 30% wood tar, and 3% Thiophanate-methyl, the 50% concentration of wood tar is significantly more effective than 30% wood tar and 3% Thiophanate-methyl in preventing Valsa canker in four years (Figure 7). From Figure 8, it can be seen that the control results of wood tar on the diseased part of Valsa canker are more significant compared with the control, and the effect of promoting healing is also very obvious.
3. Discussion
Apple Valsa canker is still a serious disease threatening the main apple-producing areas. The commonly used fungicides have not been able to achieve the ideal control effect, and the problems of residue and drug resistance became increasingly prominent [8,9]. The selection of substances that are environmentally friendly, stable in physical properties, and have bactericidal activity has become a point of attention [5,10,11,12]. Wood tar has a long history of use, and it was used for wood preservation in ancient times due to its ability to penetrate the xylem [5].
In this study, the sensitivity of wood tar to six different strains of V. mali was determined. The results suggested that wood tar exhibited sufficient antifungal activity against V. mali. In this study, wood tar treatment increased the cell membrane permeability, indicating that wood tar may disrupt the cell membrane structure of V. mali, leading to cell membrane rupture and leakage. Previous studies have shown that the virulence and the content of oxalic acid are closely related. Oxalic acid is necessary for pathogenicity [13]. In addition, pectinase plays a key role in the infection of apple bark and is one of the cell wall hydrolases secreted by V. mali [7]. In this study, the level of pectinase activity decreased after wood tar treatment, indicating that wood tar inhibited the ability of V. mali to infect apple trees. In this study, wood tar had both good protective and therapeutic activity against V. mali. Moreover, the protective activity was always better than that of curative activity, which indicated that wood tar should be used as a protective agent to better prevent the occurrence of the disease.
There is evidence that POD and PAL are key enzymes involved in plant defense [14,15]. In the presence of hydrogen peroxide, POD can oxidize phenol to quinine. PAL participates in the synthesis and accumulation of plant secondary disease resistance substances (phytoalexin, lignin, and phenolic substances) [16,17]. The formation of lignin can increase the thickness of cell walls and the degree of tissue lignification and form a mechanical barrier for pathogen invasion. After wood tar treatment, the activities of POD and PAL increased. This is consistent with the results in Figure 6 that the callus after treating the lesion with wood tar is thicker than the untreated tissue. It is of important research significance to study whether wood tar has the function of plant resistance induction. Due to winter pruning and scraping of diseased spots, V. mali invaded. The application of wood tar can not only protect the wound but also inhibit the expansion of V. mali.
The results of the field tests from 2017 to 2020 show that wood tar has a significant effect in controlling the annual infection of V. mali, and the control effect in four years has obtained unexpected results in the field. The results show that 100% and 50% wood tar demonstrated a better controlling effect, and there was no significant difference between their efficacy. Importantly, different types of wood tar showed lasting efficacy. In the first two years after application, the efficacy of the reference fungicide was slightly higher than the treatment group, while in the latter three years, the effect was reversed. The results of field experiments are affected by multiple factors. In the future, we will continue to conduct more field trials to further clarify the use method and dose of the wood tar.
Pectinase is one of the important cell wall hydrolase enzymes in V. mali infecting process. Pectinase invades and colonizes apple tissue by hydrolyzing apple cell walls. According to previous studies, after knocking out pectinase-related genes, the pathogenicity of V. mali was significantly reduced [18]. In this study, the activity of pectinase was significantly decreased after treatment with wood tar, and the levels of five genes related to pectinase were also down-regulated to varying degrees. The above results prove that wood tar can reduce the virulence of V. mali infections in apples by inhibiting the activity of pectinase and the expression level of related genes.
It is considered one of the environmentally friendly methods to find new ways to use from discarded plant resources. Because of its stable and abundant carbon source, wood tar is also a new type of pesticide that can provide nutrients to plants. Although wood tar has a complex composition, it has research prospects due to its excellent antibacterial activity and induced resistance.
In summary, this study confirmed that wood tar has antifungal activity, can inhibit the onset of plants for a long time, has a good healing effect on wounds, and can be developed as a new plant fungicide. As far as we know, this is the first report of the antifungal activity and biochemical reaction of wood tar to V. mali, which will provide new research ideas for the mechanism of wood tar. The mechanism of wood tar against V. mali was complex. It can be seen from the results that wood tar inhibits the activity of the pathogen by affecting the activity of pectinase, a key enzyme in V. mali infecting apples. This is also corroborated by the down-regulation of pectinase-related genes in V. mali. Further studies are still needed to explore the mechanism of wood tar against V. mali.
4. Materials and Methods
4.1. Fungicides, Medium, and Strains
Wood tar (100%) was purchased from Shaanxi Yixin Bioenergy Technology Development Co. Ltd. and dissolved in 10 mL ethanol to 100 mg/mL. Carbendazim (98%) provided by Shenyang Institute of Chemical Industry (Shenyang, China) was dissolved in 0.1 mol/L hydrochloric acid (hydrochloric acid).
Potato dextrose agar (PDA) was prepared with 200 g of potato, 18 g of agar, and 20 g of glucose per liter of distilled water. Six pure monospore strains collected from Shaanxi Province, China, were provided by the College of Plant Protection of Northwest A&F University and kept at 4 °C on PDA. The six V. mali strains in this study were collected from different apple branches in Shaanxi Province. The surface of apple branches with Valsa canker symptoms was cut and sterilized with 1% NaClO for 1 min, then treated with sterile water for 30 s and air-dried. The sterilized apple branches were placed on PDA plates and soaked 3 times with 100 μg/mL NaClO at 25 °C. These pure cultured V. mali strains were isolated and identified based on previous studies [19]. All these strains were maintained on the PDA slide at 4 °C prior to sensitivity testing.
4.2. Sensitivity of V. mali to Wood Tar
The sensitivity of V. mali to wood tar was tested as follows. Mycelia plugs (diameter 4 mm) taken from the colony of V. mali were placed on PDA plates containing 0, 6.25, 12.5, 25, 50, 100, and 200 μg/mL of wood tar. The plates were placed in an incubator at 25 °C. After 5 days of inoculation, the diameter of the colony was measured, and the EC50 value was calculated [20]. There are three PDA plates for each treatment. In this test, carbendazim was used as the reference fungicide, and the concentration was as follows: 0, 0.0625, 0.125, 0.25, 0.5, 1, and 2 μg/mL.
4.3. Effect of Wood Tar on Mycelial Morphology
The mycelia plugs extracted from the edge of the vigorously growing colony of the PF-15 strain were transferred to the PDA plate containing the EC50 value of wood tar. Take PDA without wood tar as a control. After 4 days, in a 25 °C incubator, the edge of the area (10 mm x 10 mm) was cut, specimens were fixed in 4% glutaraldehyde diluted in 0.1 mol/L phosphate buffer (pH 6.8) for 6–8 h at 4 °C, washed with the same buffer for 1 h, dehydrated with graded ethanol concentrations (30, 50, 70, 80, 90, 90, and 100%) for 20 min, critical point dried, mounted on stubs, and sputter-coated with a platinum-plated palladium target sand [19,21]. The hypha morphology of V. mali was observed with a scanning electron microscope (SEM, JSM-6360 LV, Japan) [22].
4.4. The Effect of Tar on the Cell Membrane Permeability
For the three strains PF-15, PF-32, and PF-35, 10 mycelia plugs were transferred to a 250 mL bottle containing 100 mL of PDB. After shaking at 180 rpm for 72 h, the EC50 value of some flasks was corrected with cuminic acid at 25 °C for 72 h. A flask without cumin acid was used as a control. After shaking the flask for 24 h, the hyphae were collected. Then, 0.5 g of fresh mycelium per sample was suspended in 25 mL of distilled water. We measured the conductivity of distilled water after 0.5, 1, 2, 3, 4, and 5 h with a conductivity meter (CON510 Eutech/Oakton, Singapore). After 5 h, the mycelium was boiled for 5 min, and the final conductivity was measured. There are three replicates for each treatment, and the experiment is three times in total. We used the following equation to calculate the relative conductivity of V. mali [23]:
Relative conductivity at different times = conductivity × 100%
4.5. Pectinases Activity and Oxalic Acid Content
Pectinases activity was determined according to Wang et al. [23]. Briefly, different volumes of D-galacturonic acid solution (0, 0.2, 0.4, 0.6, 0.8, or 1.0 mL) added with double steam water to 1 mL. Then, we added 3.0 mL of 3, 5-dinitrosalicylic acid and 0.5 mL of citric acid buffer (pH 4.8) and boiled it for 5 min. After natural cooling, a constant volume was reached by double steaming water to 5 mL. The absorbance at 540 nm was measured with a microplate reader, and the standard curve was drawn.
In order to determine the pectinase activity, the above filtrate was centrifuged at 4 °C at 12,000 rpm for 5 min. Then, the activity of pectinase in the supernatant was determined according to the standard curve. Oxalic acid content in the supernatant above was determined according to [24]. Three replicates per treatment were used, and the experiment was conducted three times.
4.6. Effect of Wood Tar on the Expression of Genes Involved in Pectinase
Total RNA was extracted with a kit (Omega Biotechnology, New York, NY, USA). Real-time quantitative polymerase chain reaction (qRT-PCR) was performed on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Richmond, CA, USA). The expression of the pectinase gene was normalized to the reference gene G6 PDH [18]. Calculations were performed using the 2-ΔΔCt method [25]. qRT-PCR primers are in Table 3. Each treatment was repeated 3 times, and the experiment was performed 3 times.
4.7. Effect of Wood Tar on the Activity of POD and PAL
Apple leaves cut from a three-year-old Fuji tree were sprayed with 1000 and 2000 mg/L of wood tar until there was runoff. After air drying, the leaves were ground in liquid nitrogen, and the activity of POD and PAL was determined using a commercial kit according to previous studies [26,27]. Ten leaves were used for each treatment, and the experiment was repeated three times.
4.8. Protective and Therapeutic Activity of Wood Tar against V. mali
The length of the detached branches from apple trees (10-year-old “Fuji trees”) was 10 cm. For protective activity, the branches were sprayed with water (control), wood tar at 1000 and 2000 μg/mL, and carbendazim at 400 μg/mL until runoff. After 24 h, mycelia plugs taken from the margin of the colony of V. mali strain 06−5 (selected at random) were placed on the branches and maintained at 25 °C. For therapeutic activity, fungicides were sprayed after 24 h of inoculation. After the branches were placed in a plastic box at 25 °C and 80% relative humidity for 72 h, the colony diameters in two vertical directions were measured, and the lesion area was calculated [28,29,30,31]. There are three plants with nine twigs for each treatment (three twigs are cut from one plant), and the experiment was repeated twice.
Lesion area (cm2) = long lesion × short lesion length × ¼
4.9. Field Test of Wood Tar against V. mali
This experiment was conducted in Dali County, Weinan City, Shaanxi Province. A total of five treatments were designed, including wood tar (30%, 50%, and 100%), 3% thiophanate-methyl ointment, and a blank control (treated with water). Fifty disease lesions of similar sizes were selected for each treatment. First, we used a scalpel to extend a 0.5 cm circle around the diseased spot and then a knife to thoroughly clean the diseased tissue and healthy skin inside the circle (the knife must be disinfected) to reach the xylem. We applied the drug evenly with a toothbrush around the spot of the diseased tissue (thickness about 2 mm) [32,33,34,35,36]. The experiment was conducted in the autumn of 2016, and the results were first investigated in the spring of the following year. After that, the results were investigated in October each year from 2017 to 2021.
4.10. Data Analysis
All the data were subjected to an analysis of variance (ANOVA) using SPSS 19.0 (SPSS Inc., Chicago, IL, USA). Duncan’s multiple range test was used to analyze the significant differences. The EC50 values were calculated using the Data Processing System (DPS, version 7.05) by plotting the percentage of mycelia growth inhibition against the log10 fungicide concentrations in three independent measurements.
Author Contributions
Y.C. and M.L. conducted the experiments and wrote the manuscript; J.Z. and K.H. provided materials and analyzed the data; Y.S. and J.F. conceived the whole experiment. All authors have read and agreed to the published version of the manuscript.
Funding
This work was sponsored by the Key R&D projects of Shaanxi Province (2020 ZDLNY-07-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
This manuscript does not contain any conflict of interest.
Sample Availability
Not applicable.
References
- Wang, L.; Gao, Z.; Huang, L.l.; Wei, J.; Zang, R.; Kang, Z.S. Screening fungicide for pathogen inhibition and disease control of apple tree Valsa canker. Acta Phytopathol. Sin. 2009, 39, 549–554. [Google Scholar]
- Byrde, R.; Crowdy, S.H.; Roach, F.A.; Byrde, R.; Crowdy, S.H.; Roach, F.A. Observations on apple canker: V. eradicant spraying and canker control. Ann. Appl. Biol. 1952, 39, 581–587. [Google Scholar] [CrossRef]
- Tamura, O.; Saito, I. Histopathological changes of apple bark infected by Valsa ceratosperma (tode ex fr.) maire during dormant and growing periods. Jpn. J. Phytopathol. 1982, 48, 490–498. [Google Scholar] [CrossRef]
- Cao, K.Q.; Guo, L.Y.; Li, B.H.; Sun, G.Y.; Chen, H.J. Investigations on the occurrence and control of apple canker in China. Plant Prot. 2009, 35, 114–117. [Google Scholar]
- Mazela, B. Fungicidal value of wood tar from pyrolysis of treated wood. Waste Manag. 2007, 27, 461–465. [Google Scholar] [CrossRef]
- Li, Y.X.; Lin, H.W. Gas chromatographic analysis of organic components in hardwood bark wood vinegar and wood tar. Chin. J. Anal. Sci. 2012, 28, 58–62. [Google Scholar]
- Yin, Z.Y.; Liu, H.Q.; Li, Z.P.; Ke, X.W.; Dou, D.L.; Gao, X.N.; Song, N.; Dai, Q.Q.; Wu, Y.X.; Xu, J.R.; et al. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N.K. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol. Technol. 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Liao, M.; Ren, X.X.; Gao, Q.; Liu, N.N.; Tang, F.; Wang, G.; Cao, H.Q. Anti-fungal activity of moso bamboo (Phyllostachys pubescens) leaf extract and its development into a botanical fungicide to control pepper phytophthora blight. Sci. Rep. 2021, 11, 4146. [Google Scholar] [CrossRef]
- Marino, G.; Gaggia, F.; Saiano, F.; Biavati, B.; Marangoni, B. Elimination of in vitro bacterial contaminants in shoot cultures of ‘MRS 2/5’ plum hybrid by the use of Melia azedarach extracts. Eur. J. Plant Pathol. 2009, 123, 195–205. [Google Scholar] [CrossRef]
- Pereira, F.; Sancha, S.; Feliciano, D.; Luo, X.; Mulhovo, S.; Duarte, A.; Madureira, A.M.; Ferreira, M. Evaluation of the antibacterial activity of some African medicinal plants. Planta Med. 2015, 81, 108. [Google Scholar] [CrossRef]
- Mangalagiri, N.P.; Panditi, S.K.; Jeevigunta, N. Antimicrobial activity of essential plant oils and their major components. Heliyon 2021, 7, e06835. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Huang, J.S.; Jenkins, S.F. Relationship of mycelial growth and production of oxalic acid and cell wall degrading enzymes to virulence in Sclerotium rolfsii. Can. J. Plant Pathol. 1985, 7, 109–117. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Wen, P.F.; Chen, J.Y.; Kong, W.F.; Pan, Q.H.; Wan, S.B.; Huang, W.D. Salicylic acid induced the expression of phenylalanine ammonia-lyase gene in grape berry. Plant Sci. 2005, 169, 928–934. [Google Scholar] [CrossRef]
- Leyva, A. Low temperature induces the accumulation of Phenylalanine Ammonia-Lyase and Chalcone Synthase mRNAs of Arabidopsis thaliana in a light-dependent manner. Plant Physiol. 1995, 108, 39–46. [Google Scholar] [CrossRef]
- Subramaniam, R.; Reinold, S.; Molitor, E.K.; Douglas, C.J. Structure, inheritance, and expression of hybrid poplar (Populus trichocarpa × Populus deltoides) phenylalanine ammonia-lyase genes. Plant Physiol. 1993, 102, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.Y.; Ke, X.W.; Huang, D.X.; Gao, X.N.; Voegele, R.T.; Kang, Z.S.; Huang, L.L. Validation of reference genes for gene expression analysis in Valsa mali var. mali using real-time quantitative PCR. World J. Microb. Biot. 2013, 29, 1563–1571. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Y.; Han, L.R.; Zhang, X.; Feng, J.T. Potential use of cuminic acid as a botanical fungicide against Valsa mali. Microb. Pthhogenesis 2017, 106, 9–15. [Google Scholar] [CrossRef]
- Lira, R.H.; Hernandez, M.; Pineda, G. Bactericidal and fungicidal activity of plant extracts from endemic plants of the Chihuahuan Desert of northern Mexico. Planta Med. 2006, 72, 213. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Y.; Zhang, Y.; Zhang, X.; Feng, J.T. Antifungal activity and biochemical response of cuminic acid against Phytophthora apsica leonian. Molecules 2016, 21, 756. [Google Scholar] [CrossRef] [Green Version]
- Jiao, H.; Fan, Y.Y.; Gao, X.N.; Huang, L.L.; Wang, X.B.; Wang, L. Control Efficacy of Eight Fungicides on Apple Valsa Canker. J. Henan Agric. Sci. 2015, 44, 95–99. [Google Scholar]
- Wang, Y.; Jiang, L.; Wang, M.M.; Feng, J.T. Baseline sensitivity and action mechanism of the sterol demethylation inhibitor flusilazole to Valsa Mali. Pestic. Biochem. Physiol. 2020, 171, 104722. [Google Scholar] [CrossRef]
- Duan, Y.B.; Ge, C.Y.; Liu, S.M.; Chen, C.J.; Zhou, M.G. Effect of phenylpyrrole fungicide fludioxonil on morphological and physiological characteristics of Sclerotinia sclerotiorum. Pestic. Biochem. Phys. 2013, 106, 61–67. [Google Scholar] [CrossRef]
- Wang, X.L.; Zang, R.; Yin, Z.Y.; Kang, Z.S.; Huang, L.L. Delimiting cryptic pathogen species causing apple Valsa canker with multilocus data. Ecol. Evol. 2014, 4, 1369–1380. [Google Scholar] [CrossRef]
- Fukasawa-Akada, T.; Kung, S.D.; Watson, J.C. Phenylalanine ammonia-lyase gene structure, expression, and evolution in Nicotiana. Plant Mol. Biol. 1996, 30, 711–722. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S.; Bulgakov, P.V. Phenylalanine ammonia-lyase and stilbene synthase gene expression in rolB transgenic cell cultures of Vitis amurensis. Appl. Microbiol. Biotechnol. 2009, 82, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Bessho, H.; Tsuchiya, S.; Soejima, J. Screening methods of apple trees for resistance to Valsa canker. Euphytica 1994, 77, 15–18. [Google Scholar] [CrossRef]
- Chen, C.; Liang, W.X.; Li, B.X.; Wang, C.X.; Dong, X.L. Effects of Temperature, Humidity, and Wound Age on Valsa mali Infection of Apple Shoot Pruning Wounds. Plant Dis. 2016, 100, 2394–2401. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.L.; Huang, L.L.; Gao, Z.P.; Ke, X.W.; Kang, Z.S. Laboratory evaluation methods of apple Valsa canker disease caused by Valsa ceratosperma sensu Kobayashi. Acta Phytopathol. Sin. 2010, 40, 14–20. [Google Scholar]
- Ma, Y.Q.; Li, J.P.; Wang, L.; Li, J.J.; Hui, N.N.; Zhou, T.W. Inhibitory effects of five fungicides on apple tree valsa canker. Gansu Agric. Sci. Technol. 2012, 6, 20–22. [Google Scholar]
- O’Neill, T.M.; PYE, D.; Locke, T. The effect of fungicides, irrigation and plant density on the development of Peronospora sparsa, the cause of downy mildew in rose and blackberry. Ann. Appl. Biol. 2002, 140, 207–214. [Google Scholar] [CrossRef]
- Sistani, F.; Ramezanpour, S.S.; Nasrollanejad, S. Field evaluation of different fungicides application to control olive leaf spot. Aust. J. Basic Appl. Sci. 2009, 3, 3341–3345. [Google Scholar]
- Large, E.C.; Beer, W.J. Field trials of copper fungicides for the control of potato blight iii. Low-copper fungicides. Ann. Appl. Biol. 2010, 33, 406–413. [Google Scholar] [CrossRef]
- Okada, K.; Furukawa, M. Occurrence and countermeasure of fungicide-resistant pathogens in vegetable field of Osaka prefecture. J. Pestic. Sci. 2008, 33, 326–329. [Google Scholar] [CrossRef] [Green Version]
- Nazanin, Z.N.; Jessica, K. Effects of host plant resistance and fungicide application on phoma stem canker, growth parameters and yield of winter oilseed rape. Crop. Prot. 2018, 112, 313–321. [Google Scholar]
Figure 1.
Effect of wood tar on the mycelial morphology of V. mali: (a–c) plates treated with EC50 concentration; (d): untreated plates.
Figure 1.
Effect of wood tar on the mycelial morphology of V. mali: (a–c) plates treated with EC50 concentration; (d): untreated plates.

Figure 2.
Effect of wood tar on the relative conductivity of the three strains (a) PF-15, (b) PF-32, and (c) PF-35. Bars denote the stand errors of three experiments.
Figure 2.
Effect of wood tar on the relative conductivity of the three strains (a) PF-15, (b) PF-32, and (c) PF-35. Bars denote the stand errors of three experiments.

Figure 3.
Effect of wood tar on the pectinase activity (a) and oxalic acid content (b). Bars denote the stand errors of three experiments. * denotes significant difference according to Fisher’s protected LSD test (p = 0.05).
Figure 3.
Effect of wood tar on the pectinase activity (a) and oxalic acid content (b). Bars denote the stand errors of three experiments. * denotes significant difference according to Fisher’s protected LSD test (p = 0.05).

Figure 4.
Relative expression levels of these five genes in three wild-type V. mali strains: PF-15, PF-32, and PF-35. The mean and standard error of three independent experiments is presented numerically. Asterisks indicate significant differences from controls according to Student’s t-test: * p < 0.05.
Figure 4.
Relative expression levels of these five genes in three wild-type V. mali strains: PF-15, PF-32, and PF-35. The mean and standard error of three independent experiments is presented numerically. Asterisks indicate significant differences from controls according to Student’s t-test: * p < 0.05.

Figure 5.
Effect of wood tar on the POD (a) and PAL (b) activity in apple leaves. Bars denote the stand errors of three experiments. Asterisk denotes significantly different from the control according to Student’s t-test: * p < 0.05.
Figure 5.
Effect of wood tar on the POD (a) and PAL (b) activity in apple leaves. Bars denote the stand errors of three experiments. Asterisk denotes significantly different from the control according to Student’s t-test: * p < 0.05.

Figure 6.
Protective and curative activity of wood tar against Valsa canker.

Figure 7.
Field efficacy of wood tar against Valsa canker. Bars denote the stand errors. Different letters above the columns indicate significant differences according to Duncan’s multiple range test (p < 0.05).
Figure 7.
Field efficacy of wood tar against Valsa canker. Bars denote the stand errors. Different letters above the columns indicate significant differences according to Duncan’s multiple range test (p < 0.05).

Figure 8.
Field efficacy of wood tar on Valsa canker: (a–f) pictures of the diseased parts of the trunk in 2021; (g,h) field pictures of control in 2021.
Figure 8.
Field efficacy of wood tar on Valsa canker: (a–f) pictures of the diseased parts of the trunk in 2021; (g,h) field pictures of control in 2021.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The sensitivity of V. mali to wood tar and carbendazim.
| Strains | EC50 (μg/mL) for | |
|---|---|---|
| Wood Tar | Carbendazim | |
| PF-15 | 76.73 bc a | 0.15 b |
| PF-18 | 83.45 b | 0.13 bc |
| PF-24 | 85.20 b | 0.19 c |
| PF-28 | 92.81 a | 0.21 c |
| PF-32 | 69.54 c | 0.08 a |
| PF-35 | 79.22 b | 0.15 b |
a Mean values followed by the same letter within the same column were not significantly different in LSD (least significant difference) tests at p = 0.05.
Table 2.
Protective and curative activity of wood tar in controlling V. mali.
| Sample | Protective Activity | Curative Activity | ||
|---|---|---|---|---|
| Lesion Area (cm2) | Control Efficacy (%) | Lesion Area (cm2) | Control Efficacy b (%) | |
| Wood Tar (2000 μg/mL) | 1.17 a a | 77.35 a | 1.813 a | 67.89 b |
| Wood Tar (1000 μg/mL) | 2.055 b | 60.22 b | 2.968 b | 47.43 c |
| Carbendazime (400 μg/mL) | 1.33 a | 74.25 a | 1.426 a | 74.74 a |
| Control | 5.166 c | - | 5.646 c | - |
a Values followed by the same letter within the same column were not different according to Fisher’s least significant difference (LSD) (p = 0.05). b Control efficacy = [(Lesion area of control -Lesion area of treated group)/(Lesion area of control)] × 100%.
Table 3.
qRT-PCR primers used in this study.
| Primer | Sequence (5′–3′) | Use |
|---|---|---|
| P1 | CTCGCCCATGTACTATGTCTTC | The qRT-PCR primer was |
| P2 | GTATCCCAGCCATCCGTATTC | for the VM1 G_05725 gene |
| P3 | CCCGCACTACTTCTTCTTTGA | The qRT-PCR primer was |
| P4 | ACGTCGCTTCCTTGGATTT | for the VM1 G_03030 gene |
| P5 | GTGACCATCTCTAACAGCCATATC | The qRT-PCR primer was |
| P6 | CTGGTCATCAGATCCCGTAAAG | for the VM1 G_06261 gene |
| P7 | GGAGGGAATAAGGTGAGGATCTA | The qRT-PCR primer was |
| P8 | CAGGCCTCAAGTACGCATTAT | for the VM1 G_04322 gene |
| P9 | CCAAAGTTCTTCAAGGCCAATC | The qRT-PCR primer was |
| P10 | GCAGGTTGTTGATGTAGTTGATG | for the VM1 G_10448 gene |
| P11 | TCAGAACAAGTTCGAGGGCGACAA | The qRT-PCR primer was |
| P12 | TGAGGGCAATAGAGGGCTTGTTCA | for the reference gene (G6 PDH gene) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, Y.; Lv, M.; Zhou, J.; Huang, K.; Sun, Y.; Feng, J. Potential Value of Wood Tar as a Natural Fungicide against Valsa mali. Molecules 2022, 27, 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051531
AMA Style
Chen Y, Lv M, Zhou J, Huang K, Sun Y, Feng J. Potential Value of Wood Tar as a Natural Fungicide against Valsa mali. Molecules. 2022; 27(5):1531. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051531
Chicago/Turabian StyleChen, Yue, Mengjing Lv, Juan Zhou, Ke Huang, Yubo Sun, and Juntao Feng. 2022. "Potential Value of Wood Tar as a Natural Fungicide against Valsa mali" Molecules 27, no. 5: 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051531