
Exploring Peptaibol’s Profile, Antifungal, and Antitumor Activity of Emericellipsin A of Emericellopsis Species from Soda and Saline Soils

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Biosynthesis of Emericellipsin A and Its Isoform by Different Emericellopsis Species
2.2. Isolation and Characterization of Emericellispsin A and DE Hydroxyl Form
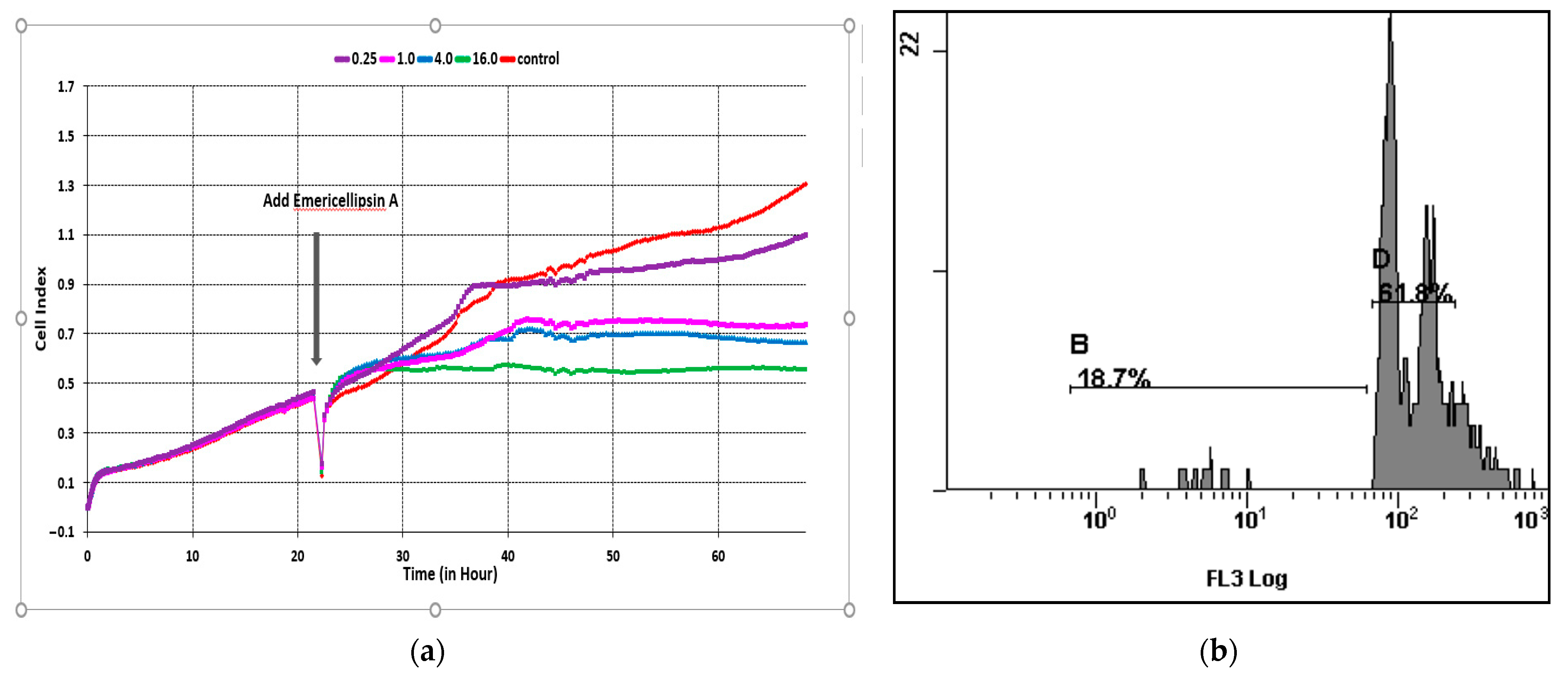
2.3. Effects of Emericellipsin A Peptide on Colorectal Carcinoma (HCT116) Cells in Real-Time Systems
3. Discussion
4. Materials and Methods
4.1. Fungal Strain Identification and Phylogenetic Analysis
4.2. Cultivation of the Fungi and Extraction of Emericellipsins
4.3. Purification and Identification of Emericellipsin A and Dehydrate Isoform
4.3.1. HPLC analysis of Prepared Samples
4.3.2. Calibration and Quantitation of Emericellipsin A
4.3.3. Sample Preparation and MALDI-TOF MS/MS Analysis
4.3.4. EmiA and dEmiA Form Detection Assay
4.4. Biological Assays
4.4.1. Antifungal Activity
4.4.2. Real-Time Cell Analyzer Test
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Debets, A.J.; Bilanenko, E.N. Are alkalitolerant fungi of the Emericellopsis lineage (Bionectriaceae) of marine origin? IMA Fungus 2013, 4, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Summerbell, R.C.; Gams, W.; Schroers, H.J.; Mitchell, J.I. A new Acremonium species associated with Fucus spp., and its affinity with a phylogenetically distinct marine Emericellopsis clade. Stud. Mycol. 2004, 50, 283–297. [Google Scholar]
- Hagestad, O.L.; Hou, L.; Andersen, J.H.; Hansen, E.H.; Altermark, B.; Li, C.; Kuhnert, E.; Cox, R.J.; Crous, P.W.; Spatafora, J.W.; et al. Genomic characterization of three marine fungi, including Emericellopsis atlantica sp. nov. with signatures of a generalist lifestyle and marine biomass degradation. IMA Fungus 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Baranova, A.A.; Georgieva, M.L.; Bilanenko, E.N.; Andreev, Y.A.; Rogozhin, E.A.; Sadykova, V.S. Antimicrobial potential of alkalophilic micromycetes Emericellopsis alkalina. Appl. Biochem. Microbiol. 2017, 53, 703–710. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Vicente, T.F.; Esteves, A.C.; Alves, A. Novel halotolerant species of Emericellopsis and Parasarocladium associated with macroalgae in an estuarine environment. Mycologia 2020, 112, 154–171. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/ (accessed on 13 February 2022).
- Ishiyama, D.; Satou, T.; Senda, H.; Fujimaki, T.; Honda, R.; Kanazawa, S. Heptaibin, a novel antifungal peptaibol antibiotic from Emericellopsis sp. BAUA8289. J. Antibiot. 2000, 53, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kulko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A Novel Lipopeptaibol Emericellipsin A with Antimicrobial and Antitumor Activity Produced by the Extremophilic Fungus Emericellopsis alkalina. Molecules. 2018, 23, 2785. [Google Scholar] [CrossRef] [Green Version]
- Sadykova, V.S.; Gavryushina, I.A.; Kuvarina, A.E.; Markelova, N.N.; Sedykhd, N.G.; Georgieva, M.L.; Barashkova, A.S.; Rogozhin, E.A. Antimicrobic Activity of the Lipopeptide Emericellipsin A Isolated from Emericellopsis alkalina against Biofilm-Forming Bacteria. Appl. Biochem. Microbiol. 2020, 56, 292–297. [Google Scholar] [CrossRef]
- Li, T.; Li, L.; Du, F.; Sun, L.; Shi, J.; Long, M.; Chen, Z. Activity and Mechanism of Action of Antifungal Peptides from Microorganisms: A Review. Molecules 2021, 26, 3438. [Google Scholar] [CrossRef]
- Flissi, A.; Ricart, E.; Campart, C.; Chevalier, M.; Dufresne, Y.; Michalik, J.; Jacques, P.; Flahaut, C.; Lisacek, F.; Leclère, V.; et al. Norine: Update of the nonribosomal peptide resource. Nucleic Acids Res. 2020, 48, D465–D469. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Paço, A.; Escada, L.F.; Albuquerque, M.S.F.; Pinto, C.A.; Saraiva, J.A.; Duarte, A.S.; Rocha-Santos, T.A.P.; Esteves, A.C.; Alves, A. Unveiling biological activities of marine fungi: The effect of sea salt. Appl. Sci. 2021, 11, 6008. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Hilário, S.; Van de Peer, Y.; Esteves, A.C.; Alves, A. Genomic and Metabolomic Analyses of the Marine Fungus Emericellopsis cladophorae: Insights into Saltwater Adaptability Mechanisms and Its Biosynthetic Potential. J. Fungi 2022, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Kuvarina, A.E.; Gavryushina, I.A.; Kulko, A.B.; Ivanov, I.A.; Rogozhin, E.A.; Georgieva, M.L.; Sadykova, V.S. The Emericellipsins A-E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multi-drug-Resistant Pathogenic Fungi. J. Fungi 2021, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Stefanowics-Hajduk, J.S.; Ochocka, R. Real-time cell analysis system in cytotoxicity applications: Usefulness and comparison with tetrazolium salt assays. Toxicol. Rep. 2020, 7, 335–344. [Google Scholar] [CrossRef]
- Hazekawa, M.; Nishinakagawa, T.; Kawakubo-Yasukochi, T.; Nakashima, M. Evaluation of IC50 levels immediately after treatment with anticancer reagents using a real-time cell monitoring device. Exp. Ther. Med. 2019, 18, 3197–3205. [Google Scholar] [CrossRef] [Green Version]
- Revie, N.M.; Iyer, K.R.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Evolution, mechanisms and impact. Curr. Opin. Microbiol. 2018, 45, 70–76. [Google Scholar] [CrossRef]
- Shadrivova, O.; Gusev, D.; Vashukova, M.; Lobzin, D.; Gusarov, V.; Zamyatin, M.; Zavrazhnov, A.; Mitichkin, M.; Borzova, Y.; Kozlova, O.; et al. COVID-19-Associated Pulmonary Aspergillosis in Russia. J. Fungi 2021, 7, 1059. [Google Scholar] [CrossRef]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 453. [Google Scholar] [CrossRef]
- Lau, J.L.; Dunn, M.K. Therapeutic Peptides: Historical Perspectives, Current Development Trends, and Future Directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Sarkar, T.; Chetia, M.; Chatterjee, S. Antimicrobial Peptides and Proteins: From Nature’s Reservoir to the Laboratory and Beyond. Front. Chem. 2021, 9, 432. [Google Scholar] [CrossRef]
- Degenkolb, T.; Fog, K.; Dieckmann, N.D.; Rocha, F.B.; Chaverri, P.; Samuels, G.J.; Thrane, U.; von Dohren, H.; Vilcinskas, A.; Bruckner, H. Peptaibol, Secondary-Metabolite, and Hydrophobin Pattern of Commercial Biocontrol Agents Formulated with Species of the Trichoderma harzianum Complex. Chem. Biodivers. 2015, 12, 662–684. [Google Scholar] [CrossRef] [PubMed]
- Shenkarev, Z.O.; Paramonov, A.S.; Nadezhdin, K.D.; Bocharov, E.V. Antiamoebin I in Methanol Solution: Rapid Exchange between Right-Handed and Left-Handed 310-Helical Conformations. Chem. Biodivers. 2007, 4, 1219–1242. [Google Scholar] [CrossRef]
- Singh, V.P.; Pathania, A.S.; Kushwaha, M.; Singh, S.; Sharma, V.; Malik, F.A.; Khan, I.; Kumar, A.; Singh, D.; Vishwakarma, R.A. 14-Residue peptaibol velutibol A from Trichoderma velutinum: Its structural and cytotoxic evaluation. RSC Adv. 2020, 10, 31233–31242. [Google Scholar] [CrossRef]
- Gavryushina, I.A.; Georgieva, M.L.; Kuvarina, A.E.; Sadykova, V.S. Peptaibols as Potential Antifungal and Anticancer Antibiotics: Current and Foreseeable Development (Review). Appl. Biochem. Microbiol. 2021, 5, 556–563. [Google Scholar] [CrossRef]
- Lee, J.W.; Collins, J.E.; Wendt, K.L.; Chakrabarti, D.; Cichewicz, R.H. Leveraging Peptaibol Biosynthetic Promiscuity for Next-Generation Antiplasmodial Therapeutics. J. Nat. Prod. 2021, 84, 503–517. [Google Scholar] [CrossRef]
- Perlatti, B.; Nichols, C.N.; Alspaugh, J.A.; James, B.; Gloer, J.B.; Bills, G.F. Sphaerostilbellins, New Antimicrobial Aminolipopeptide Peptaibiotics from Sphaerostilbella toxica. Biomolecules 2020, 10, 1371. [Google Scholar] [CrossRef]
- Berg, A.; Schlegel, B.; Ihn, W.; Demuth, U.; Gräfe, U.J. Isolation and Structural Elucidation of New Peptaibols, Bergofungins B, C and D, from Emericellopsis donezkii HKI 0059. J. Antibiot. 1999, 52, 666–669. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Balázs, D.; Urbán, P.; Szepesi, A.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; et al. Structural Diversity and Bioactivities of Peptaibol Compounds From the Longibrachiatum Clade of the Filamentous Fungal Genus Trichoderma. Front. Microbiol. 2019, 10, 1434. [Google Scholar] [CrossRef] [Green Version]
- Otto, A.; Laub, A.; Porzel, A.; Schmidt, J.; Wessjohann, L.; Westermann, B.; Arnold, N. Isolation and Total Synthesis of Albupeptins A-D: 11-Residue Peptaibols from the Fungus Gliocladium album. Eur. J. Org. Chem. 2015, 34, 7449–7459. [Google Scholar] [CrossRef]
- Casagrande, N.; Borghese, C.; Gabbatore, L.; Morbiato, L.; De Zotti, M.; Aldinucci, D. Analogs of a Natural Peptaibol Exert Anticancer Activity in Both Cisplatin- and Doxorubicin-Resistant Cells and in Multicellular Tumor Spheroids. Int. J. Mol. Sci. 2021, 22, 8362. [Google Scholar] [CrossRef]
- Dalzini, A.; Bergamini, C.; Biondi, B.; De Zotti, M.; Panighel, G.; Fato, R.; Peggion, C.; Bortolus, M.; Maniero, A.L. The Rational Search for Selective Anticancer Derivatives of the Peptide Trichogin GA IV: A Multi-Technique Biophysical Approach. Sci. Rep. 2016, 6, 24000. [Google Scholar] [CrossRef] [Green Version]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef] [PubMed]
- Luong, H.X.; Thanh, T.T.; Tran, T.H. Antimicrobial Peptides—Advances in Development of Therapeutic Applications. Life Sci. 2020, 260, 118407. [Google Scholar] [CrossRef] [PubMed]
- Hegedüs, N.; Marx, F. Antifungal Proteins: More than Antimicrobials? Fungal Biol. Rev. 2013, 26, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, G.; Wang, S.W.; Chiang, Y.R.; Chi, W.C.; Kuo, Y.H.; Phong, D.; Chen, C.Y.; Lee, T.H. Anti-inflammatory effects of peptides from a marine algicolous fungus Acremonium sp. NTU492 in BV-2 microglial cells. J. Food Drug Anal. 2020, 28, 283–291. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Racic, G.; Rakk, D.; Szekeres, A.; Vágvölgyi, C.; Kredics, L. New 19-Residue Peptaibols from Trichoderma Clade Viride. Microorganisms 2018, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Marik, T.; Urbán, P.; Tyagi, C.; Szekeres, A.; Leitgeb, B.; Vágvölgyi, M.; Manczinger, L.; Druzhinina, I.S.; Kredics, L. Diversity Profile and Dynamics of Peptaibols Produced by Green Mould Trichoderma Species in Interactions with Their Hosts Agaricus bisporus and Pleurotus ostreatus. Chem. Biodivers. 2017, 14, e1700033. [Google Scholar] [CrossRef]
- Katoch, M.; Singh, D.; Kapoor, K.K.; Vishwakarma, R.A. Trichoderma lixii (IIIM-B4), an endophyte of Bacopa monnieri L. producing peptaibols. BMC Microbiol. 2019, 19, 98. [Google Scholar] [CrossRef]
- Inostroza, A.; Lara, L.; Paz, C.; Perez, A.; Galleguillos, F.; Hernandez, V.; Becerra, J.; Gonzalez-Rocha, G.; Silva, M. Antibiotic activity of Emerimicin IV isolated from Emericellopsis minima from Talcahuano Bay, Chile. Nat. Prod. Res. 2018, 32, 1361–1364. [Google Scholar] [CrossRef]
- Norine. Available online: https://bioinfo.cristal.univ-lille.fr/norine/dform.jsp (accessed on 1 February 2022).
- Oppong-Danquah, E.; Passaretti, C.; Chianese, O.; Blümel, M.; Tasdemir, D. Mining the metabolome and the agricultural and pharmaceutical potential of sea foam-derived fungi. Mar. Drugs 2020, 18, 128. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawa, M.S.R.; Nogawa, T.; Okano, A.; Futamura, Y.; Wahab, H.A.; Osada, H. Zealpeptaibolin, an 11-mer cytotoxic peptaibol group with 3 Aib-Pro motifs isolated from Trichoderma sp. RK10-F026. J. Antibiot. 2022, 74, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Anne-Isaline van Bohemen, A.; Ruiz, N.; Zalouk-Vergnoux, A.; Michaud, A.; du Pont, T.R.; Druzhinina, I.; Atanasova, L.; Prado, S.; Bodo, B.; Meslet-Cladiere, L.; et al. Pentadecaibins I–V: 15-Residue Peptaibols Produced by a Marine-Derived Trichoderma sp. of the Harzianum Clade. J. Nat. Prod. 2021, 84, 1271–1282. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Liu, D.; Yang, Y. Anti-Cancer Peptides: Classification, Mechanism of Action, Reconstruction and Modification. Open Biol. 2020, 10, 200004. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Antimicrobial Peptides (AMPs) with Dual Mechanisms: Membrane Disruption and Apoptosis. J. Microbiol. Biotechnol. 2015, 25, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Harati, K.; Behr, B.; Daigeler, A.; Hirsch, T.; Jacobsen, F.; Renner, M.; Harati, A.; Wallner, C.; Lehnhardt, M.; Becerikli, M. Curcumin and Viscum album Extract Decrease Proliferation and Cell Viability of Soft-Tissue Sarcoma Cells: An In Vitro Analysis of Eight Cell Lines Using Real-Time Monitoring and Colorimetric Assays. Nutr. Cancer. 2017, 69, 340–351. [Google Scholar] [CrossRef]
- Kho, D.; MacDonald, C.; Johnson, R.; Unsworth, C.P.; O’Carroll, S.J.; Mez, E.D.; Graham, E.S. Application of xCELLigence RTCA biosensor technology for revealing the profile and window of drug responsiveness in real time. Biosensors 2015, 5, 199–222. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 10 February 2022).
- MAFFT Version 7. Multiple Alignment Program for Amino Acid or Nucleotide Sequences. Available online: https://mafft.cbrc.jp/alignment/server/ (accessed on 10 February 2022).
- IQ-TREE Web Server: Fast and Accurate Phylogenetic Trees under Maximum Likelihood. Available online: http://iqtree.cibiv.univie.ac.at (accessed on 10 February 2022).
- Baranova, A.A.; Rogozhin, E.A.; Georgieva, M.L.; Bilanenko, E.N.; Kulko, A.B.; Yakushev, A.V.; Alferova, V.A.; Sadykova, V.S. Antimicrobial Peptides Produced by Alkaliphilic Fungi Emericellopsis alkalina: Biosynthesis and Biological Activity Against Pathogenic Multidrug-Resistant Fungi. Appl. Biochem. Microbiol. 2019, 55, 145–151. [Google Scholar] [CrossRef]
- CLSI M38-A2. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 2nd ed.; Approved Standart; Clinical and Laboratory Standards Institute: Pittsburgh, PA, USA, 2008. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | No. of Strains, No. of VKM, VKPM, CBS Collections | pH of the Soil | Total Salts (g/kg) | EmiA, CB, mg/L | EmiA, M, mg/L | Presence of Homologues B–E |
|---|---|---|---|---|---|---|
| Soda soils | ||||||
| 1 | E101, F-4108, CBS 127350 T | 10.1 | 73 | 429.5 | 173 | d. |
| 2 | A112 | 10.1 | 33 | 26 | 18 | n.d. |
| 3 | A113, FW-1476 | 11 | 57 | 358.75 | 356.5 | d. |
| 4 | A120 | 9.9 | 310 | 28.5 | 162.5 | n.d. |
| 5 | A121 | 10.2 | 73 | 140.25 | 84.75 | n.d. |
| 6 | A123 | 219.5 | 157.25 | d. | ||
| 7 | A124 | 10.1 | 60 | 166.75 | 266.25 | n.d. |
| 8 | A125 | 10.1 | 7.1 | 71.25 | 303.5 | d. |
| 9 | A126 | 10.1 | 1.9 | 158 | 85.5 | n.d. |
| 10 | A127 | 10.1 | 1.9 | 154.75 | 160.5 | d. |
| 11 | M14, F-3905, CBS 120043 | 9.9 | 310 | 28.5 | 26 | n.d. |
| 12 | 5KS17-3 | 11.0 | 30.5 | 74.25 | n.d. | |
| 13 | 6KS17-1 | 11.0 | 140 | 268.5 | d. | |
| 14 | 7KS17-1 | 11.0 | 15.5 | 42.25 | n.d. | |
| 15 | 8KS17-1 | 11.0 | 104.5 | 283.5 | d. | |
| 16 | 9KS17-3 | 11.0 | 0 | 66 | n.d. | |
| 17 | 10KS17-1 | 11.0 | 136 | 56.25 | n.d. | |
| 18 | 14KS17-1 | 11.0 | 33 | 128 | d. | |
| 19 | 6KS10-2 | 9.8 | 48 | 27.75 | 103.5 | d. |
| Soda-chloride-sulfate soils | ||||||
| 20 | A103 | 9.6 | 100 | 28.5 | 321.75 | d. |
| 21 | A116 | 9.6 | 100 | 25.75 | 338.75 | d. |
| Chloride soils | ||||||
| 22 | A114, FW-1473 | 10 | 187 | 85 | 116 | n.d. |
| 23 | A122 | 9.5 | 65 | 101.25 | 129.5 | n.d. |
| 24 | 3KS17-13 | 8.0 | 350 | 68.5 | 136 | n.d. |
| 25 | 1KS17-1 * | 8.5 | 200 | 20.25 | 295.5 | n.d. |
| 26 | 1KS17-4 * | 8.5 | 200 | n.d. | n.d. | n.d. |
| 27 | 2KS17-1 * | 7.5 | 350 | 32.75 | 85.25 | n.d. |
| 28 | 3KS17-1 * | 8.0 | 350 | n.d. | n.d. | n.d. |
| 29 | 3KS17-2 * | 8.0 | 350 | n.d. | n.d. | n.d. |
| 30 | 3KS17-12 * | 8.0 | 350 | n.d. | n.d. | n.d. |
| Chloride-sulfate soils | ||||||
| 31 | A115, FW-1474 | 9.6 | 225 | 61 | 67.25 | n.d. |
| 32 | A117, FW-1471 | 9.9 | 53 | 61 | 231.25 | n.d. |
| 33 | A119 | 10.1 | 38 | 75.5 | 152.5 | n.d. |
| Soda-chloride soils | ||||||
| 34 | A118, VKPM F1428 | 9.6 | 137 | 262 | 184 | d. |
| 35 | M20, FW-3040,CBS 120044 | 9.6 | 137 | 342 | 202 | d. |
| Sulfate-soda soils | ||||||
| 36 | A128 | 10.3 | 139.4 | 74.5 | 249.75 | d. |
| 37 | M71, F-3907, CBS 120049 | 10.3 | 139 | 112.25 | 55 | d. |
| Undefined | ||||||
| 38 | 1KS18-5 | 9 | 3 | 131.5 | 100.5 | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuvarina, A.E.; Gavryushina, I.A.; Sykonnikov, M.A.; Efimenko, T.A.; Markelova, N.N.; Bilanenko, E.N.; Bondarenko, S.A.; Kokaeva, L.Y.; Timofeeva, A.V.; Serebryakova, M.V.; et al. Exploring Peptaibol’s Profile, Antifungal, and Antitumor Activity of Emericellipsin A of Emericellopsis Species from Soda and Saline Soils. Molecules 2022, 27, 1736. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051736
Kuvarina AE, Gavryushina IA, Sykonnikov MA, Efimenko TA, Markelova NN, Bilanenko EN, Bondarenko SA, Kokaeva LY, Timofeeva AV, Serebryakova MV, et al. Exploring Peptaibol’s Profile, Antifungal, and Antitumor Activity of Emericellipsin A of Emericellopsis Species from Soda and Saline Soils. Molecules. 2022; 27(5):1736. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051736
Chicago/Turabian StyleKuvarina, Anastasia E., Irina A. Gavryushina, Maxim A. Sykonnikov, Tatiana A. Efimenko, Natalia N. Markelova, Elena N. Bilanenko, Sofiya A. Bondarenko, Lyudmila Y. Kokaeva, Alla V. Timofeeva, Marina V. Serebryakova, and et al. 2022. "Exploring Peptaibol’s Profile, Antifungal, and Antitumor Activity of Emericellipsin A of Emericellopsis Species from Soda and Saline Soils" Molecules 27, no. 5: 1736. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27051736