
Identification of Biomarker Volatile Organic Compounds Released by Three Stored-Grain Insect Pests in Wheat

 , and
, and
Abstract
:1. Introduction
2. Results
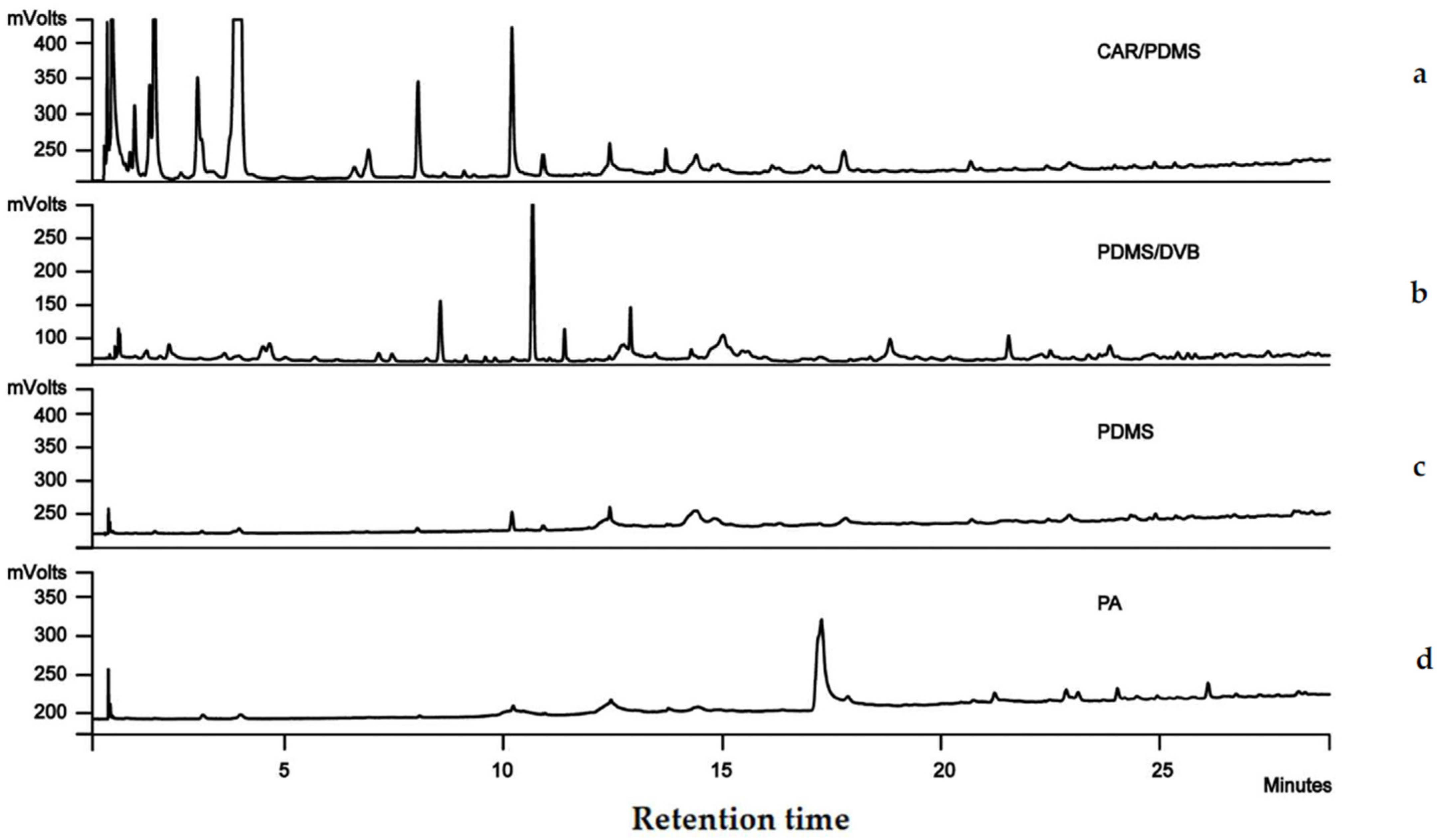
2.1. SPME Fibre Selection
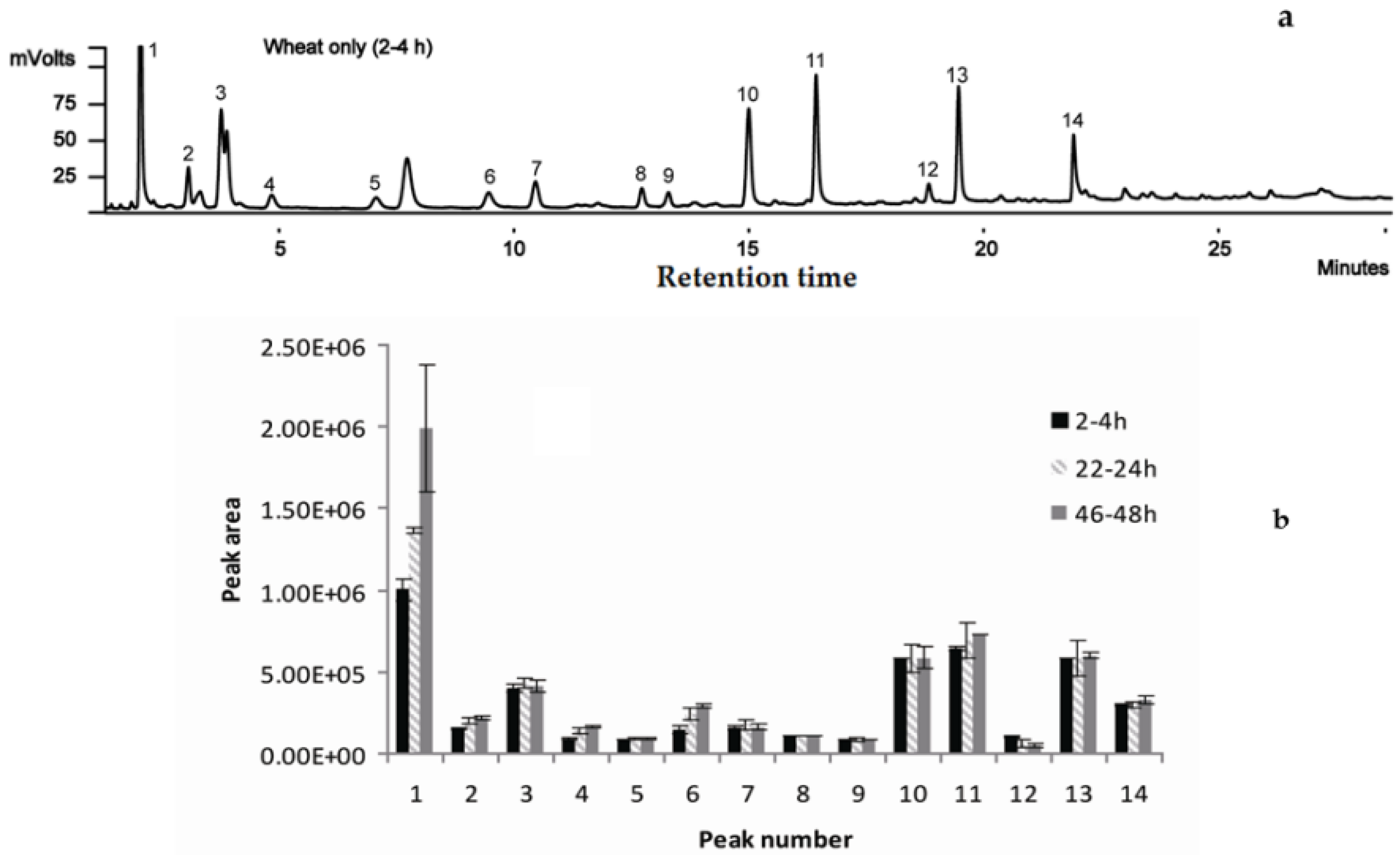
2.2. Selection of Sampling Time
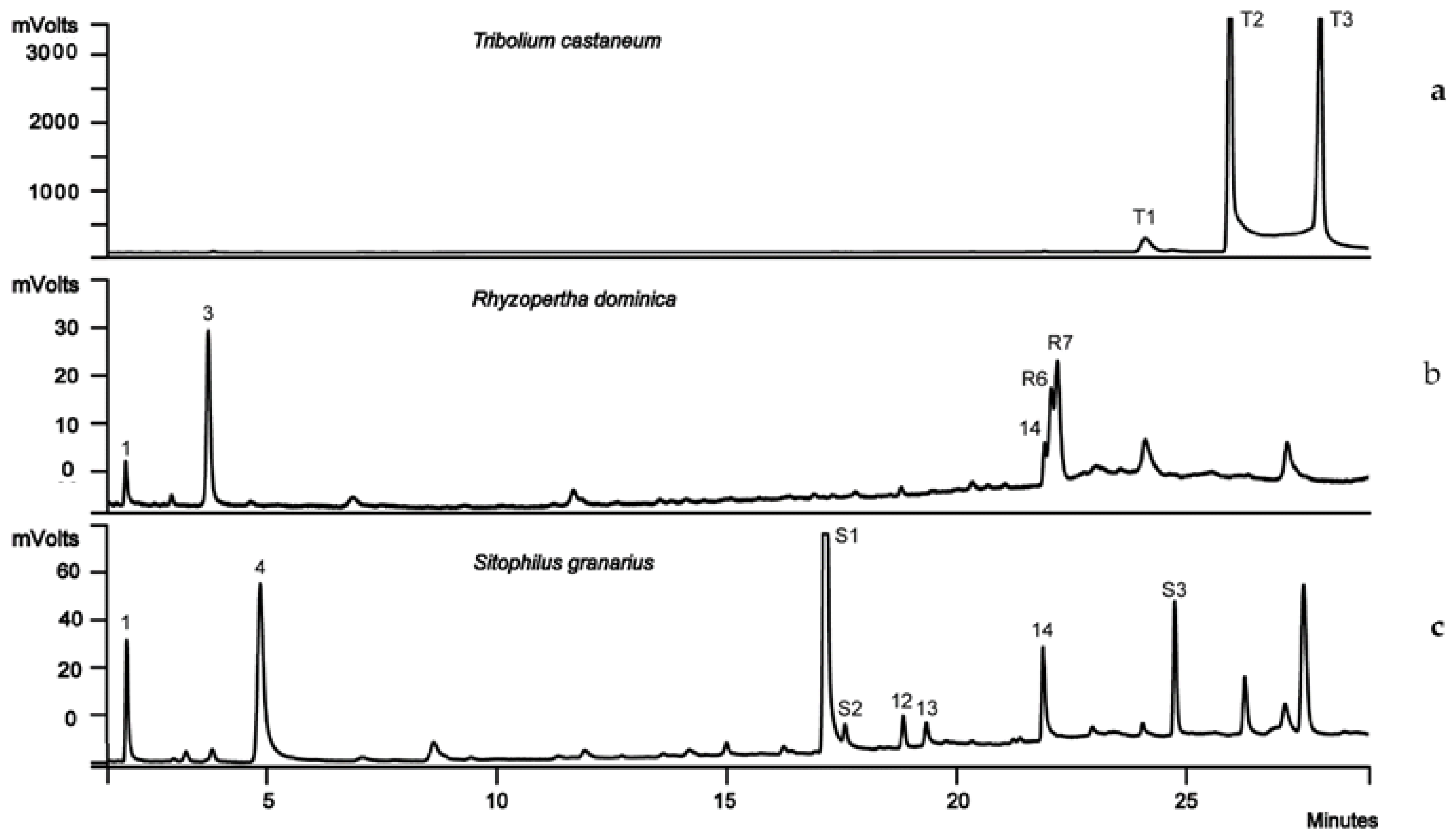
2.3. Single-Species Experiments
3. Discussion
4. Materials and Methods
4.1. Insect Materials
4.2. SPME Fibre Selection
4.3. Selection of Sampling Time
4.4. Single-Species Experiments
4.5. GC and GC-MS Analysis
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Orriss, G.D.; Whitehead, A.J. Hazard analysis and critical control point (HACCP) as a part of an overall quality assurance system in international food trade. Food Control 2000, 11, 345–351. [Google Scholar] [CrossRef]
- Whitehead, A.J. Elements of an effective national food control-system. Food Control 1995, 6, 247–251. [Google Scholar] [CrossRef]
- Kumar, D.; Kalita, P. Reducing postharvest losses during storage of grain crops to strengthen food security in developing countries. Foods 2017, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seitz, L.M.; Ram, M.S. Metabolites of lesser grain borer in grains. J. Agric. Food Chem. 2004, 52, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Cassells, J.A.; Reuss, R.; Osborne, B.G.; Wesley, I.J. Near infrared spectroscopic studies of changes in stored grain. J. Near Infrared Spec. 2007, 15, 161–167. [Google Scholar] [CrossRef]
- Brosnan, T.; Sun, D.W. Inspection and grading of agricultural and food products by computer vision systems: A review. Comput. Electron. Agric. 2002, 36, 193–213. [Google Scholar] [CrossRef]
- Chen, Y.R.; Chao, K.L.; Kim, M.S. Machine vision technology for agricultural applications. Comput. Electron. Agric. 2002, 36, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Jayas, D.S.; Symons, S.J. Identification of damaged kernels in wheat using a colour machine vision system. J. Cereal Sci. 1999, 30, 49–59. [Google Scholar] [CrossRef]
- Olsson, J.; Borjesson, T.; Lundstedt, T.; Schnurer, J. Detection and quantification of ochratoxin A and deoxynivalenol in barley grains by GC-MS and electronic nose. Int. J. Food Microbiol. 2002, 72, 203–214. [Google Scholar] [CrossRef]
- Evans, P.; Persaud, K.C.; McNeish, A.S.; Sneath, R.W.; Hobson, N.; Magan, N. Evaluation of a radial basis function neural network for the determination of wheat quality from electronic nose data. Sens. Actuators B-Chem. 2000, 69, 348–358. [Google Scholar] [CrossRef]
- Keshri, G.; Magan, N. Detection and differentiation between mycotoxigenic and non-mycotoxigenic strains of two Fusarium spp. using volatile production profiles and hydrolytic enzymes. J. Appl. Microbiol. 2000, 89, 825–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshri, G.; Voysey, P.; Magan, N. Early detection of spoilage moulds in bread using volatile production patterns and quantitative enzyme assays. J. Appl. Microbiol. 2002, 92, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridgway, C.; Chambers, J.; Portero-Larragueta, E.; Prosser, O. Detection of mite infestation in wheat by electronic nose with transient flow sampling. J. Sci. Food Agric. 1999, 79, 2067–2074. [Google Scholar] [CrossRef]
- Neethirajan, S.; Karunakaran, C.; Jayas, D.S.; White, N.D.G. Detection techniques for stored-product insects in grain. Food Control 2007, 18, 157–162. [Google Scholar] [CrossRef]
- Biswas, S.; Staff, C. Analysis of headspace compounds of distillers grains using SPME in conjunction with GC/MS and TGA. J. Cereal Sci. 2001, 33, 223–229. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Hanks, L.M. Role of host plant volatiles in mate location for three species of longhorned beetles. J. Chem. Ecol. 2005, 31, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Lacey, E.S.; Ginzel, M.D.; Millar, J.G.; Hanks, L.M. Male-produced aggregation pheromone of the Cerambycid beetle Neoclytus acuminatus acuminatus. J. Chem. Ecol. 2004, 30, 1493–1507. [Google Scholar] [CrossRef]
- Pelusio, F.; Nilsson, T.; Montanarella, L.; Tilio, R.; Larsen, B.; Facchetti, S.; Madsen, J.O. Headspace solid-phase microextraction analysis of volatile organic sulfur-compounds in black-and-white truffle aroma. J. Agric. Food Chem. 1995, 43, 2138–2143. [Google Scholar] [CrossRef]
- Rochat, D.; Ramirez-Lucas, P.; Malosse, C.; Aldana, R.; Kakul, T.; Morin, J.P. Role of solid-phase microextraction in the identification of highly volatile pheromones of two Rhinoceros beetles Scapanes australis and Strategus aloeus (Coleoptera, Scarabaeidae, Dynastinae). J. Chromatogr. A 2000, 885, 433–444. [Google Scholar] [CrossRef]
- Arnaud, L.; Lognay, G.; Verscheure, M.; Leenaers, L.; Gaspar, C.; Haubruge, E. Is dimethyldecanal a common aggregation pheromone of Tribolium flour beetles? J. Chem. Ecol. 2002, 28, 523–532. [Google Scholar] [CrossRef]
- Villaverde, M.L.; Juarez, M.P.; Mijailovsky, S. Detection of Tribolium castaneum (Herbst) volatile defensive secretions by solid phase micro extraction-capillary gas chromatography (SPME-CGC). J. Stored Prod. Res. 2007, 43, 540–545. [Google Scholar] [CrossRef]
- Phillips, J.K.; Chong, J.M.; Andersen, J.F.; Burkholder, W.E. Determination of the enantiomeric composition of (R-Star, S-Star)-1-ethylpropyl 2-methyl-3-hydroxypentanoate, the male-produced aggregation pheromone of Sitophilus granarius. Entomol Exp. Appl. 1989, 51, 149–153. [Google Scholar] [CrossRef]
- Phillips, J.K.; Miller, S.P.F.; Andersen, J.F.; Fales, H.M.; Burkholder, W.E. The chemical-identification of the granary weevil aggregation pheromone. Tetrahedron Lett. 1987, 28, 6145–6146. [Google Scholar] [CrossRef]
- Jelen, H.H.; Majcher, M.; Zawirska-Wojtasiak, R.; Wiewiorowska, M.; Wasowicz, E. Determination of geosmin, 2-methylisoborneol, and a musty-earthy odor in wheat grain by SPME-GC-MS, profiling volatiles, and sensory analysis. J. Agric. Food Chem. 2003, 51, 7079–7085. [Google Scholar] [CrossRef]
- Laopongsit, W.; Srzednicki, G.; Craske, J. Preliminary study of solid phase micro-extraction (SPME) as a method for detecting insect infestation in wheat grain. J. Stored Prod. Res. 2014, 59, 88–95. [Google Scholar] [CrossRef]
- Maeda, T.; Kim, J.H.; Ubukata, Y.; Morita, N. Analysis of volatile compounds in Polished-graded wheat flours using headspace sorptive extraction. Eur. Food Res. Technol. 2008, 227, 1233–1241. [Google Scholar]
- Mcwilliams, M.; Mackey, A.C. Wheat flavor components. J. Food Sci. 1969, 34, 493–496. [Google Scholar] [CrossRef]
- Yezerski, A.; Gilmor, T.P.; Stevens, L. Genetic analysis of benzoquinone production in Tribolium confusum. J. Chem. Ecol. 2004, 30, 1035–1044. [Google Scholar] [CrossRef]
- Alnajim, I.; Agarwal, M.; Liu, T.; Ren, Y.L. A novel method for the analysis of volatile organic compounds (VOCs) from red glour beetle Tribolium castaneum (H.) using headspace-SPME technology. Curr. Anal. Chem. 2020, 16, 404–412. [Google Scholar] [CrossRef]
- Qazi, M.C.B.; Boake, C.R.B.; Lewis, S.M. The femoral setiferous glands of Tribolium castaneum males and production of the pheromone 4,8-dimethyldecanal. Entomol. Exp. Appl. 1998, 89, 313–317. [Google Scholar] [CrossRef]
- Suzuki, T.; Sugawara, R. Isolation of an aggregation pheromone from the flour beetles, Tribolium castaneum and Tribolium confusum (Coleoptera, Tenebrionidae). Appl. Entomol. Zool. 1979, 14, 228–230. [Google Scholar] [CrossRef] [Green Version]
- Bashir, T.; Birkinshaw, L.A.; Farman, D.; Hall, D.R.; Hodges, R.J. Pheromone release by Rhyzopertha dominica (F) (Coleoptera: Bostrichidae) in the laboratory: Daily rhythm, inter-male variation and association with body weight and/or boring activity. J. Stored Prod. Res. 2002, 39, 159–169. [Google Scholar] [CrossRef]
- Niu, Y.; Hua, L.; Hardy, G.; Agarwal, M.; Ren, Y. Analysis of volatiles from stored wheat and Rhyzopertha dominica (F.) with solid phase microextraction-gas chromatography mass spectrometry. J. Sci. Food Agric. 2016, 96, 1697–1703. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.; Chen, S.C.; Chen, Y.R.; Ho, H.Y. 3-Hydroxy-2-butanone and the first encounter fight in the male lobster cockroach, Nauphoeta cinerea. Naturwissenschaften 2006, 93, 286–291. [Google Scholar] [CrossRef]
- Dukic, N.; Andric, G.; Glinwood, R.; Ninkovic, V.; Andjelkovic, B.; Radonjic, A. The effect of 1-pentadecene on Tribolium castaneum behaviour: Repellent or attractant? Pest. Manag. Sci. 2021, 77, 4034–4039. [Google Scholar] [CrossRef]
- Stevenson, B.J.; Cai, L.; Faucher, C.; Michie, M.; Berna, A.; Ren, Y.; Anderson, A.; Chyb, S.; Xu, W. Walking responses of Tribolium castaneum (Coleoptera: Tenebrionidae) to its aggregation pheromone and odors of wheat infestations. J. Econ. Entomol. 2017, 110, 1351–1358. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | Volatile Collection Time (h) |
|---|---|
| 20 g Homogenized wheat only | 2–4, 22–24, 46–48 |
| 20 Tribolium castaneum (Herbst) adults only | 2–4 |
| 20 T. castaneum adults + 20 g homogenized wheat | 2–4 |
| 80 Rhyzopertha dominica (F.) adults only | 22–24 |
| 80 R. dominica adults + 20 g homogenized wheat | 22–24 |
| 100 Sitophilus granarius (L.) only | 46–48 |
| 100 S. granarius adults + 20 g homogenized wheat | 46–48 |
| Compounds | RT | NIST RI | Kovats | Match Quality | GC Response (105) ± SD, n = 4 | |||
|---|---|---|---|---|---|---|---|---|
| (min) | indices | (%) | Wheat | Wheat + T. castaneum | Wheat + R. dominica | Wheat + S. granarius | ||
| Acetone | 1.18 | 1116 | 862 | 79.5 | 112.53 ± 6.77 | 124.36 ± 8.03 | 17.53 ± 3.35 | 127.18 ± 9.06 |
| Methanol | 2.51 | 1157 | 901 | 77.0 | 32.42 ± 4.29 | 43.28 ± 5.51 | 33.09 ± 4.52 | 32.57 ± 4.19 |
| Ethanol | 3.48 | 1215 | 943 | 79.1 | 75.52 ± 6.14 | 5.049 ± 2.75 | 49.51 ± 5.02 | 27.55 ± 3.66 |
| 2,3-Butanedione | 4.43 | 1248 | 977 | 91.8 | 9.88 ± 2.22 | 15.49 ± 3.93 | 65.16 ± 5.15 | 12.17 ± 3.83 |
| Ethyl acetate | 5.38 | 1291 | 988 | 85.0 | nd | nd | 27.16 ± 3.33 | nd |
| 2-Butanol | 7.02 | 1332 | 996 | 71.3 | 8.83 ± 2.04 | 7.59 ± 2.09 | 7.65 ± 2.08 | 6.94 ± 2.11 |
| Hexanal | 9.47 | 1360 | 1087 | 77.2 | 11.27 ± 3.15 | 19.08 ± 3.31 | 26.14 ± 3.81 | 23.51 ± 3.05 |
| 2-Methyl-1-propanol | 10.49 | 1381 | 1125 | 80.1 | 23.53 ± 3.27 | 6.86 ± 2.88 | 7.15 ± 2.11 | 7.22 ± 2.02 |
| 11-Methylpentacosane | 11.47 | 1418 | 1150 | 89.0 | nd | nd | 102.27 ± 8.04 | nd |
| 1-Butanol | 12.53 | 1435 | 1173 | 71.9 | 13.84 ± 4.09 | 9.55 ± 2.73 | 9.17 ± 2.50 | 8.58 ± 2.22 |
| 1-Penten-3-ol | 13.25 | 1463 | 1198 | 97.1 | 11.62 ± 3.15 | 8.11 ± 3.11 | 9.59 ± 2.72 | 8.06 ± 3.06 |
| 3-Methyl-1-butanol | 14.82 | 1488 | 1216 | 94.3 | 76.77 ± 5.26 | 58.92 ± 5.38 | 61.49 ± 4.85 | 51.64 ± 5.17 |
| 1-Pentanol | 16.34 | 1502 | 1238 | 84.4 | 103.52 ± 6.33 | 95.13 ± 7.09 | 117.25 ± 8.88 | 99.64 ± 7.61 |
| Palmitic acid | 17.57 | 1527 | 1251 | 96.0 | nd | nd | 55.72 ± 4.92 | nd |
| 3-Methyl-2-buten-1-ol | 18.72 | 1544 | 1280 | 91.0 | 10.08 ± 2.01 | 4.73 ± 2.10 | 22.57 ± 3.27 | 4.08 ± 2.06 |
| 1-Hexanol | 19.44 | 1581 | 1305 | 75.9 | 85.83 ± 5.09 | 78.59 ± 7.77 | 101.55 ± 8.21 | 91.21 ± 5.06 |
| Unknown | 20.81 | - | - | - | nd | nd | 31.09 ± 4.48 | nd |
| 1-Pentadecene | 21.91 | 1640 | 1349 | 92.2 | nd | nd | >250 | nd |
| Dominicalure 1 | 23.58 | 1685 | 1377 | 88.7 | nd | nd | 26.05 ± 4.17 | nd |
| Apparent homologs of dominicalure 1 | 24.23 | 1751 | 1391 | 94.0 | nd | nd | 143.95 ± 8.49 | nd |
| Benzene, 1-ethoxy-4-isothiocyanato- | 24.68 | 1830 | 1424 | 95.7 | nd | 74.93 ± 6.28 | nd | nd |
| Stearic acid | 26.11 | 1892 | 1451 | 75.9 | nd | 68.37 ± 5.92 | nd | nd |
| Unknown | 27.84 | - | - | - | nd | 11.5.08 ± 2.50 | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, L.; Macfadyen, S.; Hua, B.; Zhang, H.; Xu, W.; Ren, Y. Identification of Biomarker Volatile Organic Compounds Released by Three Stored-Grain Insect Pests in Wheat. Molecules 2022, 27, 1963. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061963
Cai L, Macfadyen S, Hua B, Zhang H, Xu W, Ren Y. Identification of Biomarker Volatile Organic Compounds Released by Three Stored-Grain Insect Pests in Wheat. Molecules. 2022; 27(6):1963. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061963
Chicago/Turabian StyleCai, Lijun, Sarina Macfadyen, Baozhen Hua, Haochuan Zhang, Wei Xu, and Yonglin Ren. 2022. "Identification of Biomarker Volatile Organic Compounds Released by Three Stored-Grain Insect Pests in Wheat" Molecules 27, no. 6: 1963. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27061963