
NMR, LC-MS Characterization of Rydingia michauxii Extracts, Identification of Natural Products Acting as Modulators of LDLR and PCSK9

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
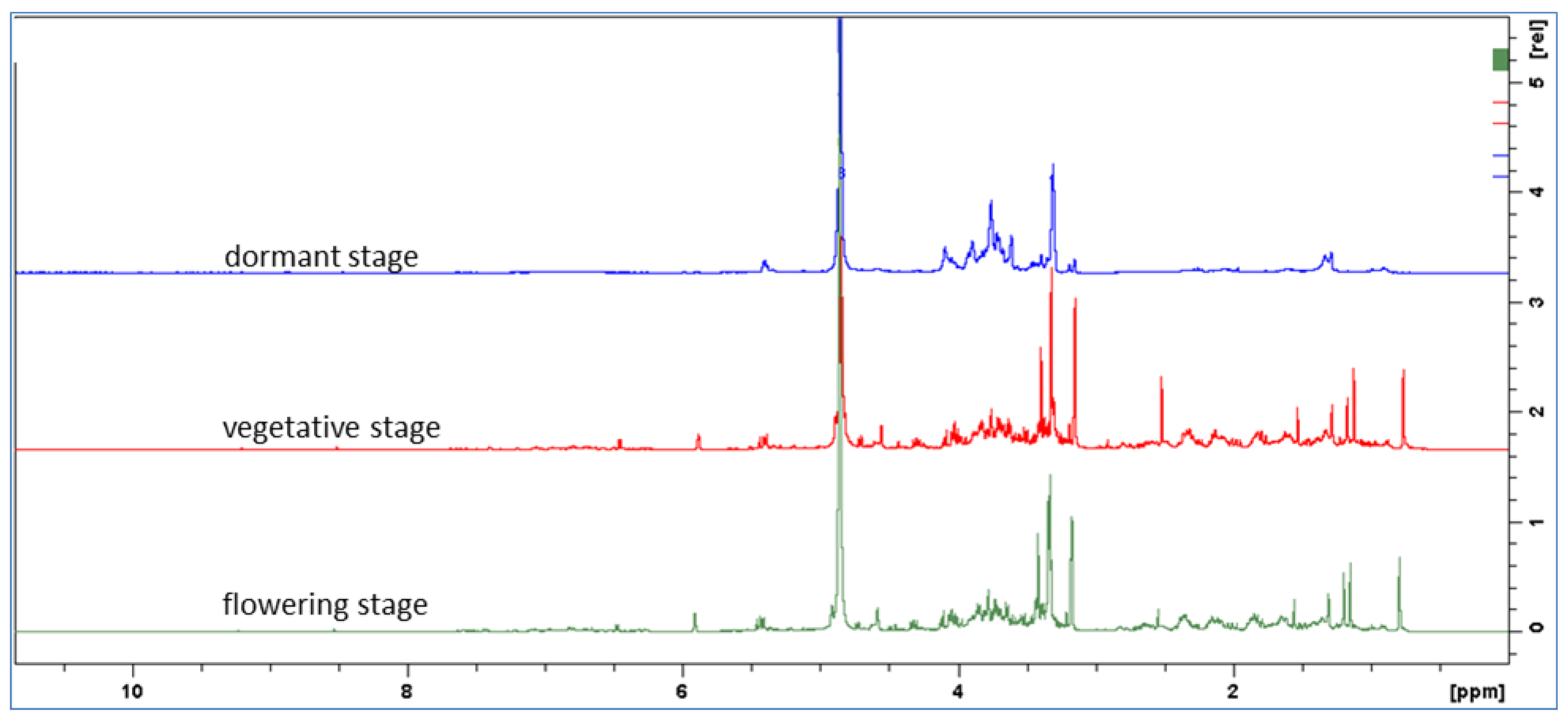
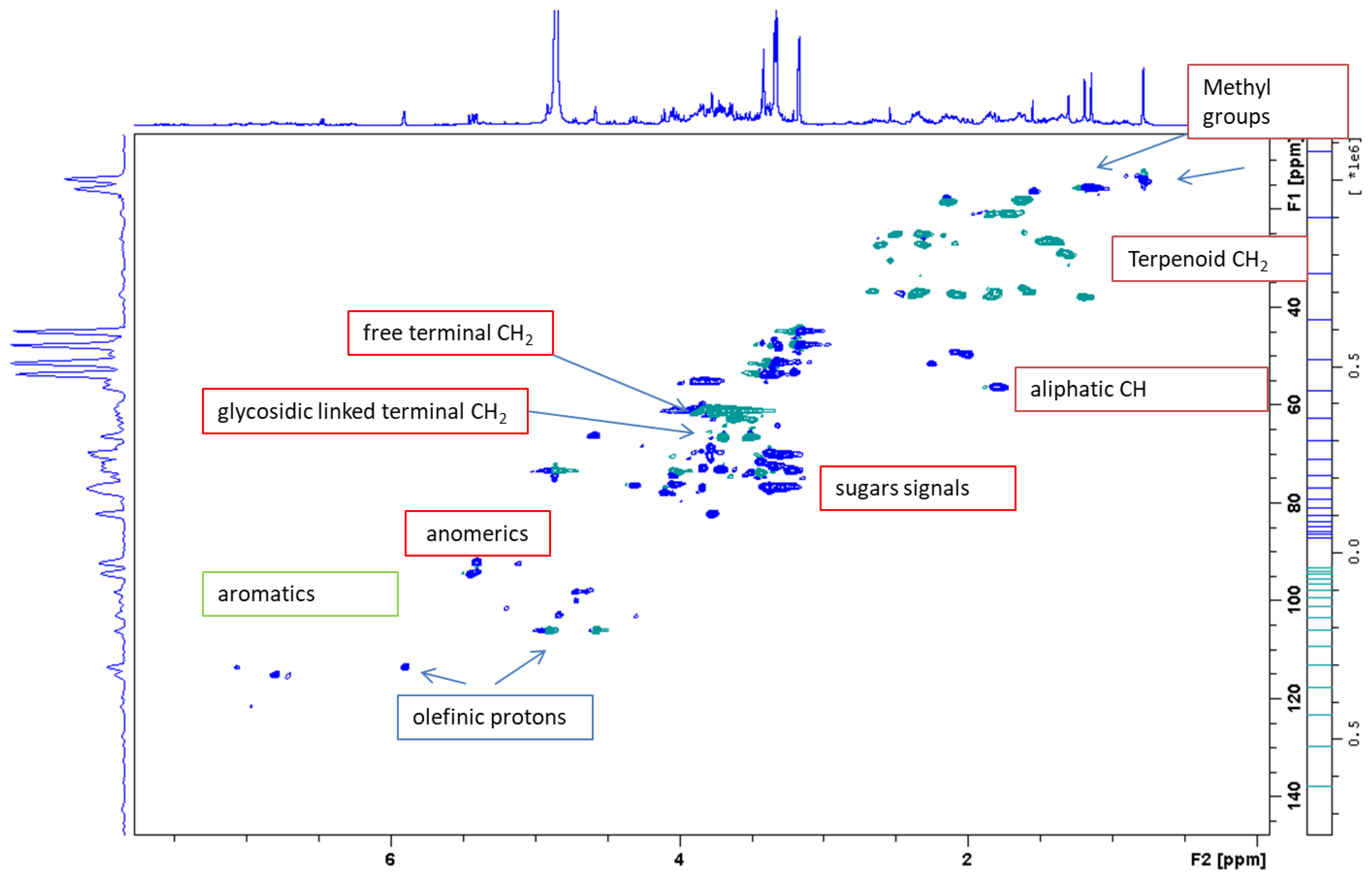
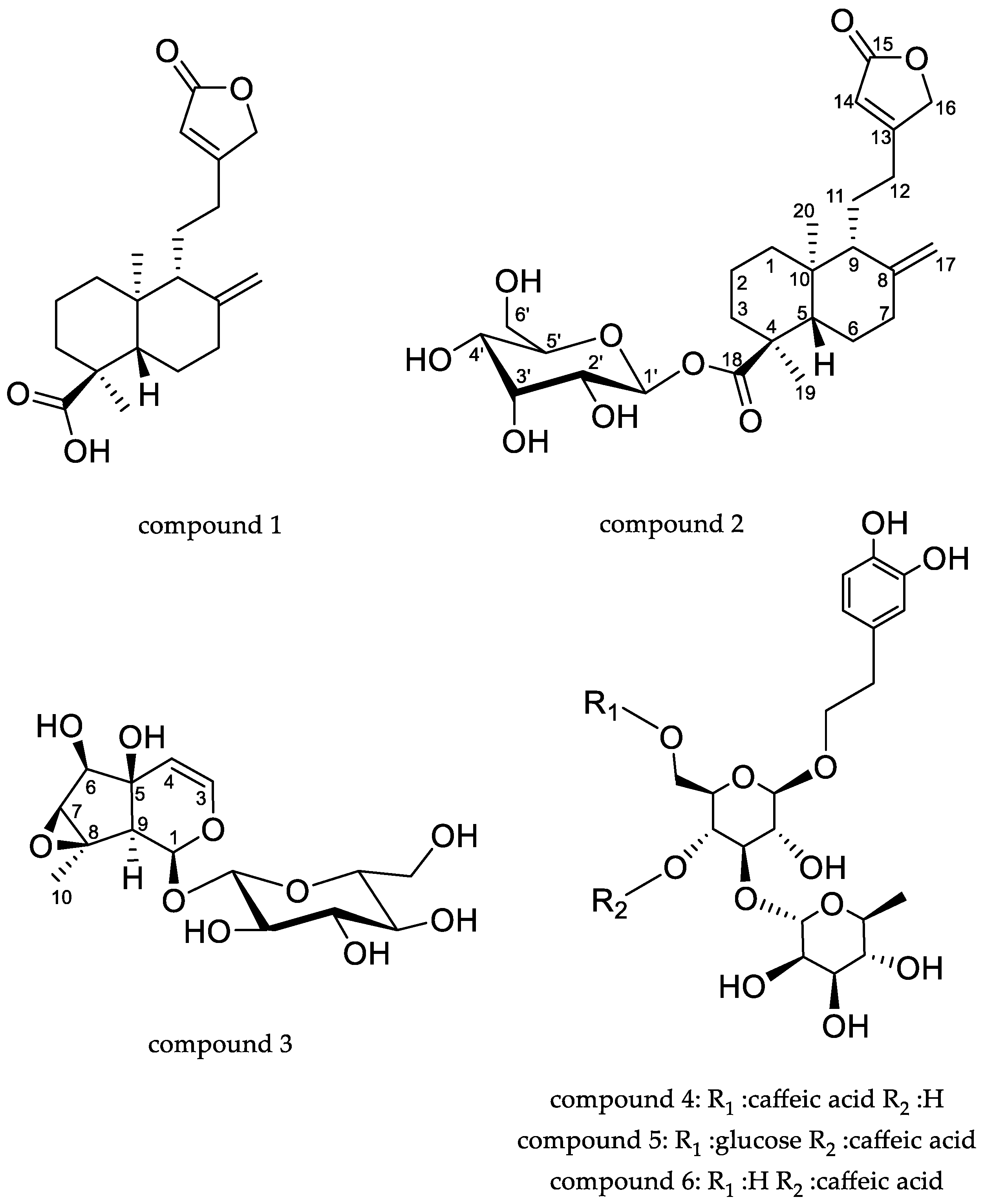
2.1. Phytochemical Characterization of the Extracts
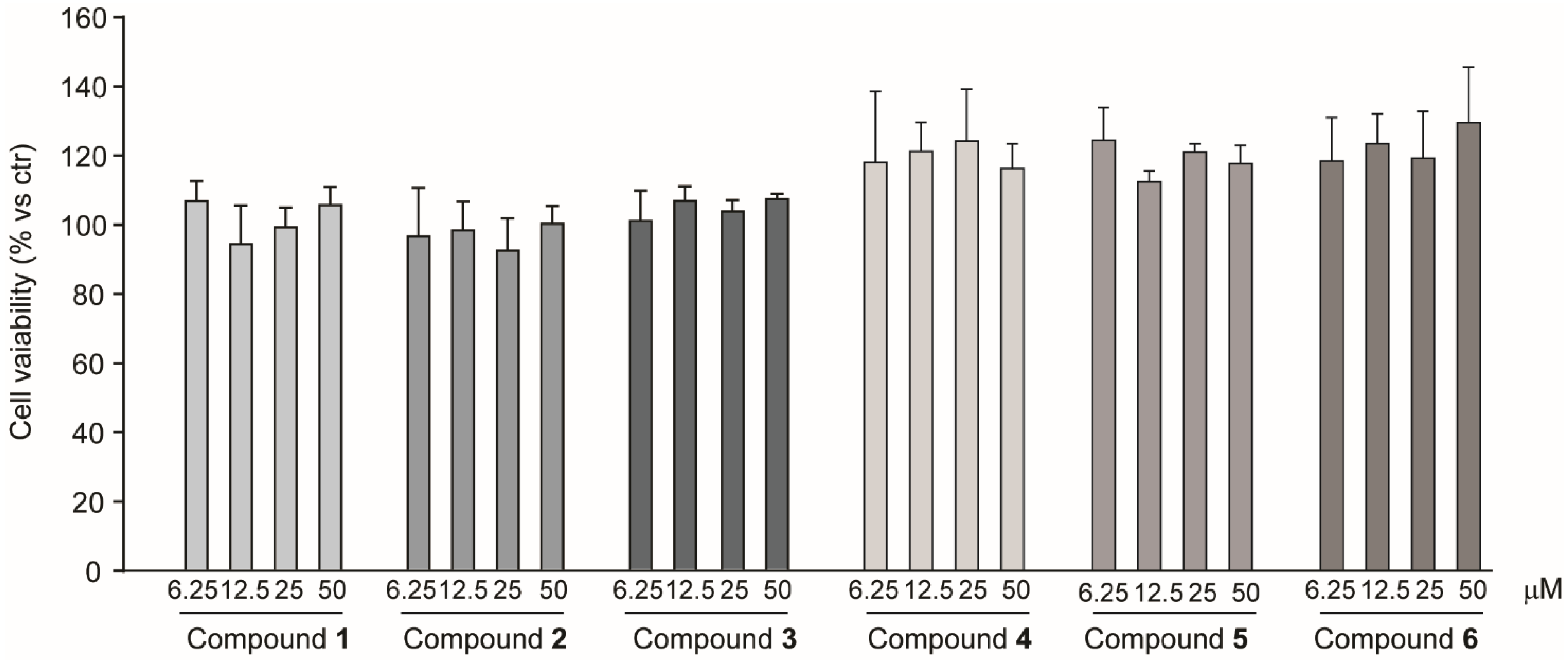
2.2. Cytotoxic Effect on Hepatocarcinoma Cell Line Huh7 of Compounds Isolated from R. michauxii Extracts
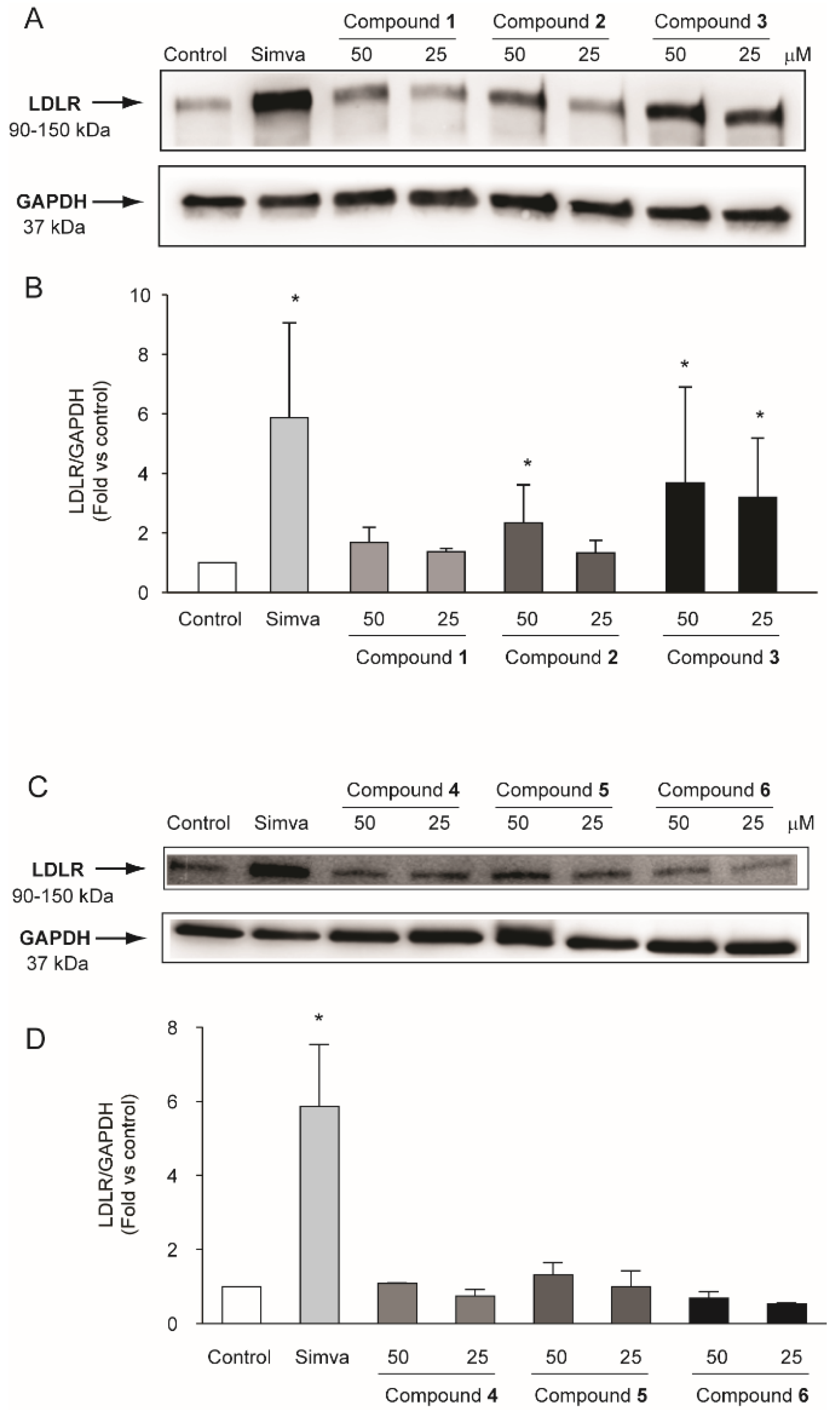
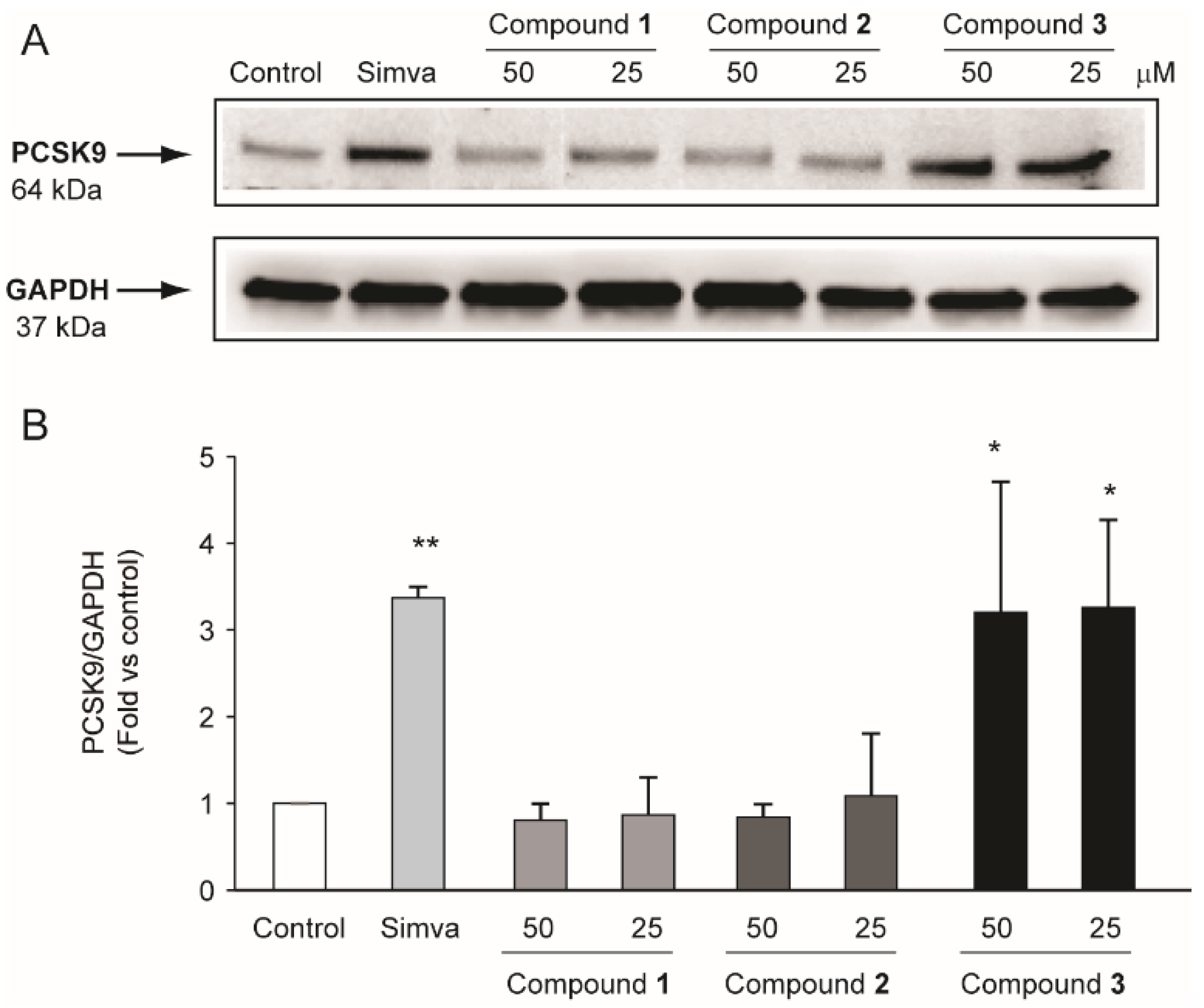
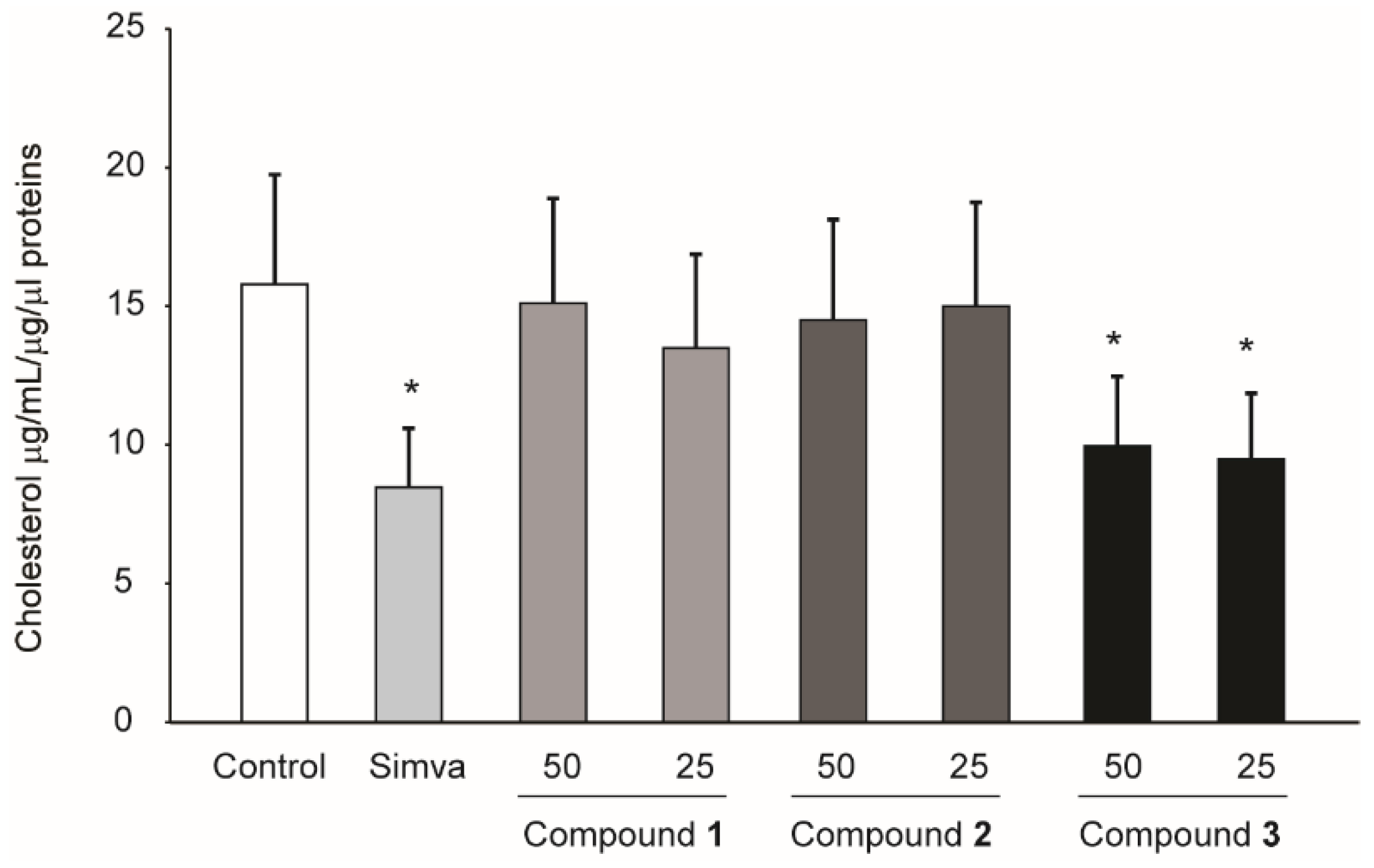
2.3. Compounds Isolated from R. michauxii Extract Increases LDLR and PCSK9 Expression in the Huh7 Cell Line
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Extract Preparation
4.3. Phytochemical Characterization, NMR, HPLC-DAD-MSn, Optical Rotation Power Measurament
4.4. Isolation and Structural Elucidation of Phytoconstituents: Semipreparative HPLC, Determination of the Absolute Configuration of the Sugar Residues after Hydrolysis
4.5. In Vitro Experiments
4.5.1. Reagents
4.5.2. Cell Cultures
4.5.3. Cell Viability Assay on Huh7
4.5.4. Western Blot Analysis
4.5.5. Cholesterol Determination
4.5.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheen, A.; Albert, V.A. Nomenclatural and Taxonomic Changes within the Leucas Clade (Lamioideae; Lamiaceae). Syst. Geogr. Plants 2007, 77, 229–238. [Google Scholar]
- Karami, A.; Esmaeili, H.; Shahbazian, D. Essential oil composition of Rydingia michauxii (Briq.) Scheen & V.A. Albert endemic of Iran. Nat. Prod. Res. 2017, 32, 229–233. [Google Scholar] [PubMed]
- Sadeghi, Z.; Kuhestani, K.; Abdollahi, V.; Mahmood, A. Ethnopharmacological studies of indigenous medicinal plants of Saravan region, Baluchistan, Iran. J. Ethnopharmacol. 2014, 153, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, A.; Hosseini, S.M.; Karami, A.; Reza, A.; Olounabadi, S. Variation in essential oil composition of Rydingia michauxii at the three developmental stages Variation in essential oil composition of Rydingia. Nat. Prod. Res. 2021, 35, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Candela, R.G.; Rosselli, S.; Bruno, M.; Fontana, G. A Review of the Phytochemistry, Traditional Uses, and Biological Activities of the Genus Ballota and Otostegia. Planta Med. 2021, 87, 432–479. [Google Scholar] [PubMed]
- Alilou, M.; Marzocco, S.; Hofer, D.; Rapa, S.F.; Asadpour, R.; Schwaiger, S.; Troppmair, J.; Stuppner, H. Labdane-Type Diterpenes from the Aerial Parts of Rydingia persica: Their Absolute Configurations and Protective Effects on LPS-Induced Inflammation in Keratinocytes. J. Nat. Prod. 2020, 83, 2456–2468. [Google Scholar] [CrossRef] [PubMed]
- Mach, F.; Baigent, C.; Catapano, A.L.; Koskinas, K.C.; Casula, M.; Badimon, L.; Chapman, M.J.; De Backer, G.G.; Delgado, V.; Ference, B.A.; et al. 2019 ESC/EAS Guidelines for the management of dyslipidaemias: Lipid modification to reduce cardiovascular risk. Eur. Heart J. 2020, 41, 111–188. [Google Scholar] [CrossRef]
- Wang, N.; Fulcher, J.; Abeysuriya, N.; Park, L.; Kumar, S.; Di Tanna, G.L.; Wilcox, I.; Keech, A.; Rodgers, A.; Lal, S. Intensive LDL cholesterol-lowering treatment beyond current recommendations for the prevention of major vascular events: A systematic review and meta-analysis of randomised trials including 327,037 participants. Lancet Diabetes Endocrinol. 2020, 8, 36–49. [Google Scholar] [CrossRef]
- Mihaylova, B.; Emberson, J.; Blackwell, L.; Keech, A.; Simes, J.; Barnes, E.H.; Voysey, M.; Gray, A.; Collins, R.; Baigent, C. The effects of lowering LDL cholesterol with statin therapy in people at low risk of vascular disease: Meta-analysis of individual data from 27 randomised trials. Lancet 2012, 380, 581–590. [Google Scholar]
- Santos, R.D. EUROASPIRE V and uncontrolled risk factors in primary prevention: Atherosclerotic cardiovascular disease in the making. Eur. J. Prev. Cardiol. 2020, 28, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Stroes, E.S.; Thompson, P.D.; Corsini, A.; Vladutiu, G.D.; Raal, F.J.; Ray, K.K.; Roden, M.; Stein, E.; Tokgözoğlu, L.; Nordestgaard, B.G.; et al. Statin-associated muscle symptoms: Impact on statin therapy-European Atherosclerosis Society Consensus Panel Statement on Assessment, Aetiology and Management. Eur. Heart J. 2015, 36, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Fogacci, F.; Banach, M.; Mikhailidis, D.P.; Bruckert, E.; Toth, P.P.; Watts, G.F.; Reiner, Ž.; Mancini, J.; Rizzo, M.; Mitchenko, O.; et al. Safety of red yeast rice supplementation: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2019, 143, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Gallelli, L.; Cannataro, R.; Perri, M.; Calignano, A.; Citraro, R.; Russo, E.; Gareri, P.; Corsonello, A.; Sarro, G. De When Nutraceuticals Reinforce Drugs Side Effects: A Case Report. Curr. Drug Saf. 2016, 11, 264–266. [Google Scholar] [CrossRef]
- Polsani, V.R.; Jones, P.H.; Ballantyne, C.M.; Nambi, V. A case report of myopathy from consumption of red yeast rice. J. Clin. Lipidol. 2008, 2, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Raschi, E.; Girardi, A.; Poluzzi, E.; Forcesi, E.; Menniti-Ippolito, F.; Mazzanti, G.; De Ponti, F. Adverse Events to Food Supplements Containing Red Yeast Rice: Comparative Analysis of FAERS and CAERS Reporting Systems. Drug Saf. 2018, 41, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Bergonzi, M.C.; Mazzi, G.; Vincieri, F.F. NMR spectroscopy: A useful tool for characterisation of plant extracts, the case of supercritical CO2 arnica extract. J. Pharm. Biomed. Anal. 2002, 30, 321–330. [Google Scholar] [CrossRef]
- Chauthe, S.K.; Sharma, R.J.; Aqil, F.; Gupta, R.C.; Singh, I.P. Quantitative NMR: An applicable method for quantitative analysis of medicinal plant extracts and herbal products. Phytochem. Anal. 2012, 23, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Deborde, C.; Fontaine, J.X.; Jacob, D.; Botana, A.; Nicaise, V.; Richard-Forget, F.; Lecomte, S.; Decourtil, C.; Hamade, K.; Mesnard, F.; et al. Optimizing 1D 1H-NMR profiling of plant samples for high throughput analysis: Extract preparation, standardization, automation and spectra processing. Metabolomics 2019, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Etienne, O.K.; Dall’Acqua, S.; Sinan, K.I.; Ferrarese, I.; Sut, S.; Sadeer, N.B.; Mahomoodally, M.F.; Ak, G.; Zengin, G. Chemical characterization, antioxidant and enzyme inhibitory effects of Mitracarpus hirtus extracts. J. Pharm. Biomed. Anal. 2021, 194, 113799. [Google Scholar] [CrossRef]
- Sut, S.; Dall’Acqua, S.; Uysal, S.; Zengin, G.; Aktumsek, A.; Picot-Allain, C.; Mahomoodally, F. LC-MS, NMR fingerprint of Potentilla argentea and Potentilla recta extracts and their in vitro biopharmaceutical assessment. Ind. Crops Prod. 2019, 131, 125–133. [Google Scholar] [CrossRef]
- Dall’acqua, S.; Sinan, K.I.; Ferrarese, I.; Sut, S.; Bene, K.; Mahomoodally, M.F.; Bibi Sadeer, N.; Ak, G.; Zengin, G. Chromatographic separation of Breynia retusa (Dennst.) alston bark, fruit and leaf constituents from bioactive extracts. Molecules 2020, 25, 5537. [Google Scholar] [CrossRef] [PubMed]
- Markham, K.R.; Chari, V.M. Carbon-13 NMR Spectroscopy of Flavonoids. In The Flavonoids: Advances in Research; Harborne, J.B., Mabry, T.J., Eds.; Springer: Boston, MA, USA, 1982; pp. 19–134. ISBN 978-1-4899-2915-0. [Google Scholar]
- Andary, C.; Wylde, R.; Laffite, C.; Privat, G.; Winternitz, F. Structures of verbascoside and orobanchoside, caffeic acid sugar esters from Orobanche rapum-genistae. Phytochemistry 1982, 21, 1123–1127. [Google Scholar] [CrossRef]
- Zou, P.; Song, Y.; Lei, W.; Li, J.; Tu, P.; Jiang, Y. Application of 1H NMR-based metabolomics for discrimination of different parts and development of a new processing workflow for Orobanche rapum-genistae. Acta Pharm. Sin. B 2017, 7, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Blunder, M.; Orthaber, A.; Bauer, R.; Bucar, F.; Kunert, O. Efficient identification of flavones, flavanones and their glycosides in routine analysis via off-line combination of sensitive NMR and HPLC experiments. Food Chem. 2017, 218, 600–609. [Google Scholar] [CrossRef]
- Clifford, M.N.; Kirkpatrick, J.; Kuhnert, N.; Roozendaal, H.; Salgado, P.R. LC–MSn analysis of the cis isomers of chlorogenic acids. Food Chem. 2008, 106, 379–385. [Google Scholar] [CrossRef]
- Li, L.; Tsao, R.; Liu, Z.; Liu, S.; Yang, R.; Young, J.C.; Zhu, H.; Deng, Z.; Xie, M.; Fu, Z. Isolation and purification of acteoside and isoacteoside from Plantago psyllium L. by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1063, 161–169. [Google Scholar] [CrossRef]
- Kachlicki, P.; Piasecka, A.; Stobiecki, M.; Marczak, L. molecules Structural Characterization of Flavonoid Glycoconjugates and Their Derivatives with Mass Spectrometric Techniques. Molecules 2016, 21, 1494. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Yin, C.; Qin, Y.; Cheng, Z.; Chen, D. Approach to the study of flavone di-C-glycosides by high performance liquid chromatography-tandem ion trap mass spectrometry and its application to characterization of flavonoid composition in Viola yedoensis. J. Mass Spectrom. 2014, 49, 1010–1024. [Google Scholar] [CrossRef]
- Zdero, C.; Bohlmann, F.; King, R.M. Guaianolides and labdanes from Brickellia species. Phytochemistry 1991, 30, 1591–1595. [Google Scholar] [CrossRef]
- Henrick, C.A.; Jefferies, P.R. The chemistry of the euphorbiaceae—XI: The diterpenes of ricinocarpus muricatus and the stereochemistry of eperuic acid. Tetrahedron 1965, 21, 1175–1190. [Google Scholar] [CrossRef]
- Hu, C.L.; Xiong, J.; Wang, P.P.; Ma, G.L.; Tang, Y.; Yang, G.X.; Li, J.; Hu, J.F. Diterpenoids from the needles and twigs of the cultivated endangered pine Pinus kwangtungensis and their PTP1B inhibitory effects. Phytochem. Lett. 2017, 20, 239–245. [Google Scholar] [CrossRef]
- Franzyk, H.; Frederiksen, S.M.; Jensen, S.R. Synthesis of monoterpene piperidines from the iridoid glucoside antirrhinoside. J. Nat. Prod. 1997, 60, 1012–1016. [Google Scholar] [CrossRef]
- Kobayashi, H.; Oguchi, H.; Takizawa, N.; Miyase, T.; Ueno, A.; Usmanghani, K.; Ahmad, M. New Phenylethanoid Glycosides from Cistanche tubulosa (SCHRENK) HOOK. f. I. Chem. Pharm. Bull. 1987, 35, 3309–3314. [Google Scholar] [CrossRef] [Green Version]
- Mamadalieva, N.Z.; Turginov, O.T.; Zengin, G.; Akramov, D.K.; Ak, G.; Honkeldieva, M.T.; Tojibaev, K.S. Chemical Composition and Biological Activity of Constituents of Otostegia bucharica. Chem. Nat. Compd. 2021, 57, 180–182. [Google Scholar] [CrossRef]
- Parvin, A.; Yaghmaei, P.; Noureddini, M.; Haeri Roohani, S.A.; Aminzadeh, S. Comparative effects of quercetin and hydroalcoholic extract of Otostegia persica boiss with atorvastatin on atherosclerosis complication in male wistar rats. Food Sci. Nutr. 2019, 7, 2875–2887. [Google Scholar] [CrossRef] [Green Version]
- Danielewski, M.; Matuszewska, A.; Szeląg, A.; Sozański, T. The impact of anthocyanins and iridoids on transcription factors crucial for lipid and cholesterol homeostasis. Int. J. Mol. Sci. 2021, 22, 6074. [Google Scholar] [CrossRef]
- Liu, L.; Liao, P.; Wang, B.; Fang, X.; Li, W.; Guan, S. Oral administration of baicalin and geniposide induces regression of atherosclerosis via inhibiting dendritic cells in ApoE-knockout mice. Int. Immunopharmacol. 2014, 20, 197–204. [Google Scholar] [CrossRef]
- Shen, D.; Zhao, D.; Yang, X.; Zhang, J.; He, H.; Yu, C. Geniposide against atherosclerosis by inhibiting the formation of foam cell and lowering reverse lipid transport via p38/MAPK signaling pathways. Eur. J. Pharmacol. 2019, 864, 172728. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Sun, C.; Liu, S.; Yan, Y.; Pan, H.; Fan, M.; Xue, L.; Nie, C.; Zhang, H.; et al. Geniposide reduces cholesterol accumulation and increases its excretion by regulating the FXR-mediated liver-gut crosstalk of bile acids. Pharmacol. Res. 2020, 152, 104631. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Jin, Q.; Xia, K.L.; Jiang, M.; Cui, B.W.; Wu, Y.L.; Song, S.Z.; Lian, L.H.; Nan, J.X. Liver kinase B1/AMP-activated protein kinase-mediated regulation by gentiopicroside ameliorates P2X7 receptor-dependent alcoholic hepatosteatosis. Br. J. Pharmacol. 2018, 175, 1451–1470. [Google Scholar] [CrossRef] [Green Version]
- Sut, S.; Pavela, R.; Kolarcik, V.; Cappellacci, L.; Petrelli, R.; Maggi, F.; Dall’Acqua, S.; Benelli, G. Identification of onosma visianii roots extract and purified shikonin derivatives as potential acaricidal agents against tetranychus urticae. Molecules 2017, 22, 1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comai, S.; Dall’Acqua, S.; Grillo, A.; Castagliuolo, I.; Gurung, K.; Innocenti, G. Essential oil of Lindera neesiana fruit: Chemical analysis and its potential use in topical applications. Fitoterapia 2010, 81, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, E.; Nossova, L.; Korenevsky, A.; Beveridge, T.J. The structure of the capsular polysaccharide of Shewanella oneidensis strain MR-4. Carbohydr. Res. 2005, 340, 1750–1753. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Constituents and Assignments | δ H | δ C | Significant Correlations |
|---|---|---|---|
| Flavonoids: apigenin and other derivatives | |||
| H 2′-6′ (apigenin) | 7.60 m | 130.3 | HMBC: 162.0 COSY: 6.42 |
| H 3′-5′ (apigenin) | 6.42 d (J = 8.0 Hz) | 115.0 | |
| H-3 (apigenin or other flavonoids) | 6.68 s | 102.8 | |
| H 6-8 of O-glycosidic flavonoids | 6.81 * m | 90.7 | HMBC: 163.5 |
| 6.78 m | 94.7 | ||
| 6.59 * m | 97.3 | ||
| 6.57 m | 89.9 | ||
| Anomeric signals of sugar residues (O-glycosides) | 5.11 d (J = 7.5 Hz) | 92.7 | COSY 3.30–3.41–3.85–4.10 |
| 5.18 * m | 101.7 | ||
| 5.38 * m | 92.3 | ||
| 5.42 * m | 94.6 | ||
| 4.73 * m | 98.5 | ||
| 4.71 * m | 100.0 | ||
| 4.67 * m | 98.0 | ||
| Anomeric signals of sugar residues (C-glycosides) | 4.91 * m | 73.2 | COSY 3.32–3.30–3.46 |
| 4.85 * m | 74.1 | ||
| Sugar positions of interglycosydic bonds | 3.77 m | 82.0 | COSY 3.30–3.41–3.85–4.10; |
| 3.51 m | 81.8 | HMBC: 101.7–98.5 | |
| Free CH2OH of sugar | 3.80–3.53 m | 60.5–61.1 | |
| CH2OH of sugar with glycosydic linkage | 3.71–3.52 m | 67.2 | |
| Caffeic acid derivatives, Verbascoside/Echinacoside and similar | |||
| H-7 (double bond) | 7.55 d (J = 16 Hz) | 144.6 | HMBC: 165.6- 166.8 |
| 7.48 d (J = 16 Hz) | 146.9 | COSY: 6.30 | |
| H-8 (double bond) | 6.30 d (J = 16 Hz) | 113.5 | COSY: 7.55–7.48 |
| H-2 (aromatic ring) | 7.07 (J = 2 Hz) | 114.7 | |
| H-6 | 6.93 m | 121.8 | |
| H-5 | 6.90 m | 115.3 | HMBC: 147.9 |
| Terpenoids | |||
| Quaternary methyl group | 0.75 s | 14.3 | HMBC: 177.3 49.0 36.3 |
| Quaternary methyl group | 1.15 s | 16.3 | HMBC: 182.4 49.0 36.3 |
| Quaternary methyl group | 1.18 s | 16.3 | HMBC: 56.2 49.1 40.0 36.3 |
| aliphatic CH2 | 1.17 m | 38.5 | |
| 1.66 m | 18.5 | ||
| 2.10–2.45 m | 37.6 | ||
| 1.68–1.80 m | 21.8 | ||
| Olefinic proton signals | 4.48–4.92 | 106.5 | HMBC: 37.6 146.0 56 |
| 5.90 brs | 114.0 | HMBC 176.0 21.8 |
| RT | [M − H]− | Fragmentation | Compound | Dormant Stage % | Vegetative Stage % | Flowering Stage % |
|---|---|---|---|---|---|---|
| hydroxycinnamic derivative | ||||||
| 9.0 | 353 | 191 171 | chlorogenic acid * | nd | 0.045 ± 0.005 | 0.166 ± 0.008 |
| 10.3 | 917 | 755 593 461 315 | lavandulofolioside hexoside/samioside hexoside | 0.081 ± 0.010 | 0.079 ± 0.010 | 0.057 ± 0.002 |
| 10.4 | 785 | 623 461 315 | Echinacoside * (5) | 0.497 ± 0.018 | 1.766 ± 0.199 | 1.633 ± 0.174 |
| 11.2 | 799 | 623 461 315 | feruloyl verbascoside | 0.462 ± 0.015 | 0.546 ± 0.060 | 0.220 ± 0.046 |
| 11.4 | 931 | 755 | feruloyl samioside | 0.019 ± 0.0015 | 0.031 ± 0.002 | nd |
| 11.6 | 623 | 461 315 | Verbascoside * (6) | 0.066 ± 0.005 | 2.364 ± 0.067 | 4.004 ± 0.122 |
| 11.8 | 755 | 593 461 315 | lavandulofolioside/samioside | 0.104 ± 0.010 | 0.556 ± 0.010 | 0.758 ± 0.010 |
| 12.0 | 623 | 461 315 | Isoverbascoside (4) | 0.110 ± 0.010 | 2.363 ± 0.205 | 1.145 ± 0.241 |
| 12.1 | 945 | 769 637 491 | unknown phenylethanoid glycosides | 0.048 ± 0.009 | 0.301 ± 0.008 | 0.154 ± 0.080 |
| 12.2 | 813 | 637 491 | unknown phenylethanoid glycosides | 0.057 ± 0.006 | 0.585 ± 0.080 | 0.298 ± 0.030 |
| 14.5 | 651 | 505 475 328 | martynoside | 0.088 ± 0.0013 | 0.283 ± 0.020 | 0.219 ± 0.005 |
| total amount of hydroxycinnamic derivative | 1.531 | 8.918 | 8.655 | |||
| C glycosylated flavonoid | ||||||
| 9.4 | 623 | 533 503 413 383 | 4′-methoxyluteolin -6,8-di-C-glucopyranoside | nd | 0.005 ± 0.001 | 0.007 ± 0.001 |
| 9.6 | 609 | 519 489 399 369 | luteolin-6,8-di-C-glucoside | nd | 0.011 ± 0.002 | 0.014 ± 0.002 |
| 10.2 | 593 | 503 473 383 353 | apigenin 6,8-di-C-glucoside * | 0.004 ± 0.001 | 0.131 ± 0.010 | 0.220 ± 0.004 |
| 10.8 | 563 | 503 473 443 353 | apigenin-6-C-glucoside-8-C-xyloside | nd | 0.119 ± 0.034 | 0.149 ± 0.008 |
| 11.8 | 577 | 487 457 367 337 | deoxy apigenin 6,8-di-C-glucoside | nd | 0.028 ± 0.006 | 0.030 ± 0.005 |
| total amount of C glycosylated flavonoid | 0.004 | 0.295 | ||||
| O glycosylated flavonoid | ||||||
| 8.4 | 417 | 285 179 163 152 | kaempferol pentoside | nd | 0.026 ± 0.001 | 0.049 ± 0.001 |
| 9.1 | 401 | 269 | apigenin pentoside | 0.001 ± 0.001 | 0.004 ± 0.001 | 0.007 ± 0.001 |
| 13.0 | 447 | 285 243 199 175 | luteolin hexoside | 0.001 ± 0.001 | 0.153 ± 0.001 | 0.169 ± 0.001 |
| 13.4 | 489 | 447 285 | kaempferol/luteolin acetyl hexoside | nd | 0.069 ± 0.001 | 0.036 ± 0.001 |
| 13.4 | 577 | 269 | apigenin rutinoside (isomer 2) | nd | 0.019 ± 0.001 | 0.030 ± 0.001 |
| 13.5 | 431 | 269 | apigenin-7-O-glucoside * | nd | 0.019 ± 0.001 | 0.034 ± 0.001 |
| 14.5 | 285 | 241 199 175 154 133 | Luteolin * | nd | 0.020 ± 0.001 | 0.026 ± 0.001 |
| 15.6 | 577 | 269 | apigenin rutinoside (isomer 1) | nd | 0.099 ± 0.001 | 0.284 ± 0.001 |
| total amount of O glycosylated flavonoid | 0.002 | 0.408 | 0.634 | |||
| gallic acid derivative | ||||||
| 10.1 | 387 | 207 163 | medioresinol | 0.026 ± 0.003 | 2.033 ± 0.087 | 0.105 ± 0.017 |
| labdane diterpenoids | ||||||
| 16.1 | 539 # 493 | 331 303 233 221 | ent-labda-8(17),13-dien-18-oic acid-15,16- glucopyranoside (2) | 0.165 ± 0.006 | 9.387 ± 0.001 | 10.081 ± 0.090 |
| 16.1 | 825 | 667 331 303 233 221 | ent-labda-8(17),13-dien-18-oic acid-15,16-olide derivative | 0.002 ± 0.001 | 3.662 ± 0.001 | 2.954 ± 0.090 |
| 16.5 | 529 | 331 303 233 221 | ent-labda-8(17),13-dien-18-oic acid-15,16-olide derivative | 0.040 ± 0.005 | 1.305 ± 0.001 | 1.089 ± 0.058 |
| 17.5 | 331 | 303 233 221 | ent-labda-8(17),13-dien-18-oic acid-15,16-olide (1) | 0.057 ± 0.002 | 6.161 ± 0.302 | 6.448 ± 0.203 |
| total labdane derivatives | 0.26 | 20.5 | 20.6 | |||
| iridoid | ||||||
| 1.2 | 407 # 361 | 199 179 | antirrhinoside (3) | nd | 7.58 ± 0.05 § | 5.53 ± 0.09 § |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sut, S.; Tahmasebi, A.; Ferri, N.; Ferrarese, I.; Rossi, I.; Panighel, G.; Lupo, M.G.; Maggi, F.; Karami, A.; Dall’Acqua, S. NMR, LC-MS Characterization of Rydingia michauxii Extracts, Identification of Natural Products Acting as Modulators of LDLR and PCSK9. Molecules 2022, 27, 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072256
Sut S, Tahmasebi A, Ferri N, Ferrarese I, Rossi I, Panighel G, Lupo MG, Maggi F, Karami A, Dall’Acqua S. NMR, LC-MS Characterization of Rydingia michauxii Extracts, Identification of Natural Products Acting as Modulators of LDLR and PCSK9. Molecules. 2022; 27(7):2256. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072256
Chicago/Turabian StyleSut, Stefania, Aminallah Tahmasebi, Nicola Ferri, Irene Ferrarese, Ilaria Rossi, Giovanni Panighel, Maria Giovanna Lupo, Filippo Maggi, Akbar Karami, and Stefano Dall’Acqua. 2022. "NMR, LC-MS Characterization of Rydingia michauxii Extracts, Identification of Natural Products Acting as Modulators of LDLR and PCSK9" Molecules 27, no. 7: 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27072256