
Quantification of Protein “Biomarkers” in Wheat-Based Food Systems: Dealing with Process-Related Issues

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
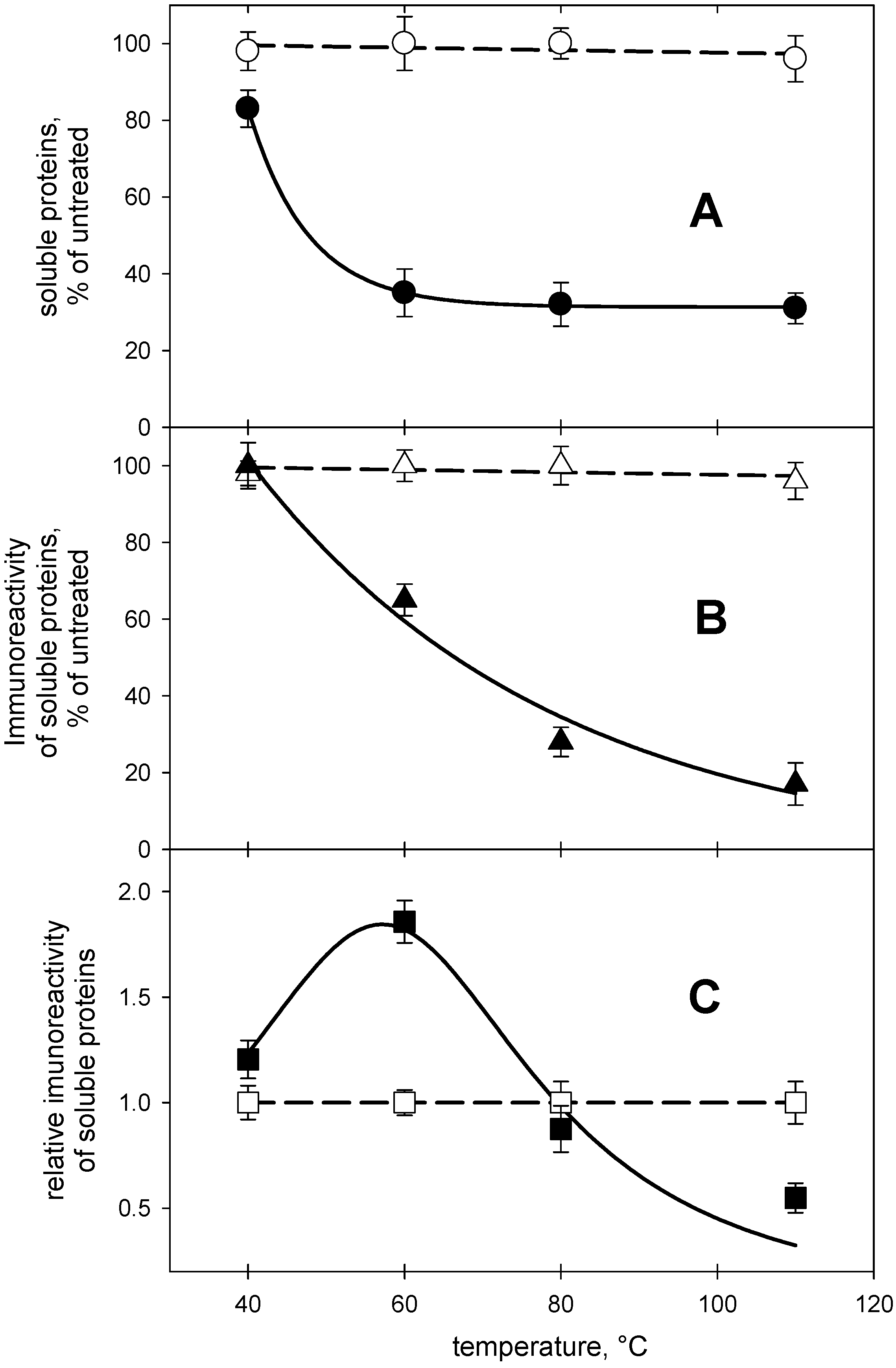
2.1. Detection of Gliadin in Processed Dough
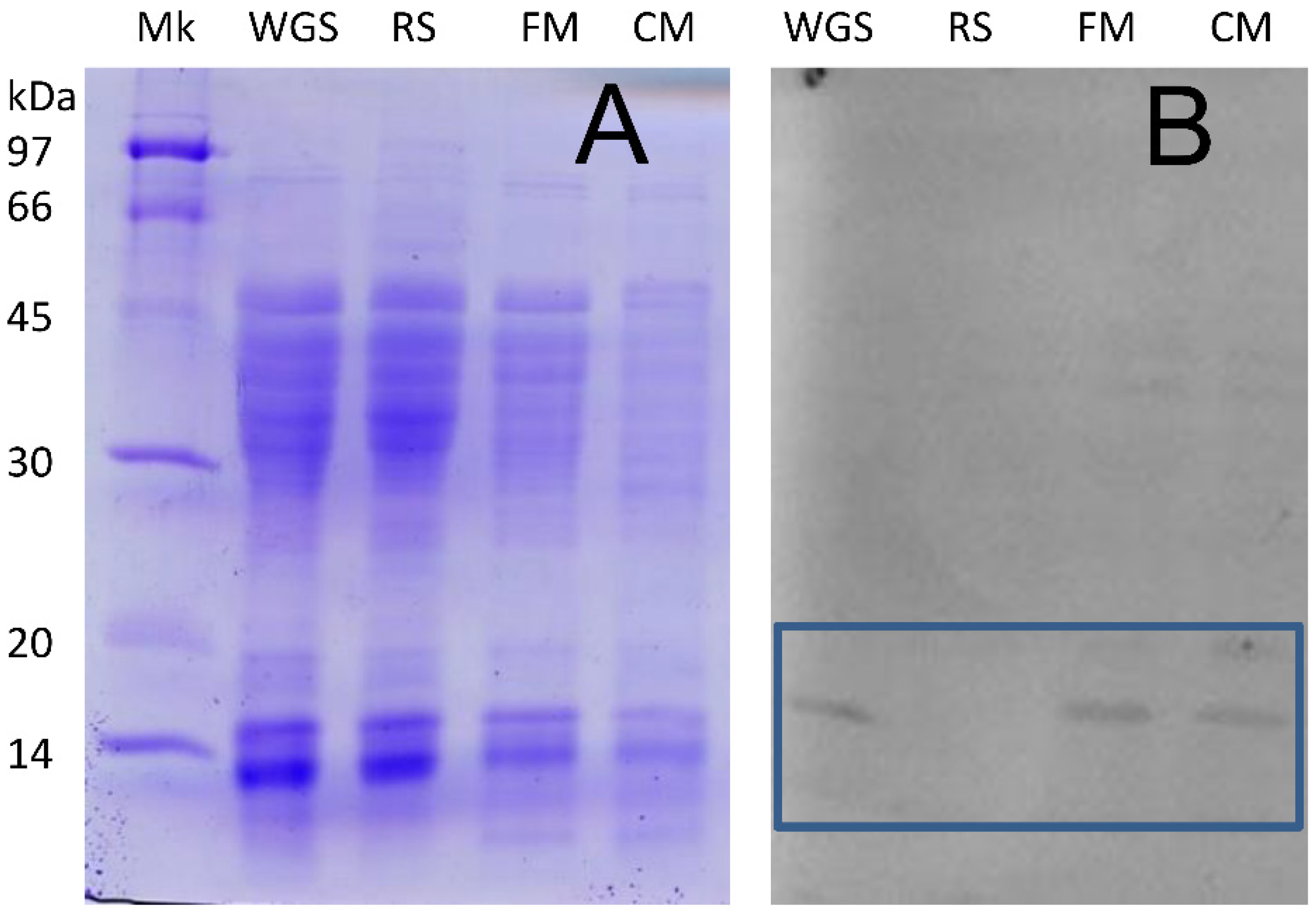
2.2. Detection of Non-Gluten Wheat Proteins
2.3. Detection of Non-Wheat Proteins in Pasta
3. Materials and Methods
3.1. Materials
3.2. Protein Solubilization, Enrichment, and Identification
3.3. Immunochemistry
3.4. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Senyuva, H.Z.; Jones, I.B.; Sykes, M.; Baumgartner, S. A critical review of the specifications and performance of antibody and DNA-based methods for detection and quantification of allergens in foods. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2019, 36, 507–547. [Google Scholar] [CrossRef] [PubMed]
- Korte, R.; Oberleitner, D.; Brockmeyer, J. Determination of food allergens by LC-MS: Impacts of sample preparation, food matrix, and thermal processing on peptide detectability and quantification. J. Proteom. 2019, 196, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Calvo, E.; García-García, A.; Madrid, R.; Martin, R.; García, T. From polyclonal sera to recombinant antibodies: A review of immunological detection of gluten in foodstuff. Foods 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Mattarozzi, M.; Careri, M. The role of incurred materials in method development and validation to account for food processing effects in food allergen analysis. Anal. Bioanal. Chem. 2019, 411, 4465–4480. [Google Scholar] [CrossRef]
- Bonomi, F.; Iametti, S. Thiolomics of the Gluten Protein Network of Wheat Dough. In Encyclopedia of Food Grains 2: Nutrition and food grains; Wrigley, C., Corke, H., Seetharaman, K., Faubion, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 2, pp. 154–160. [Google Scholar]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.; Khosla, C. Structural Basis for Gluten Intolerance in Celiac Sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [Green Version]
- Marengo, M.; Amoah, I.; Carpen, A.; Benedetti, S.; Zanoletti, M.; Buratti, S.; Lutterodt, H.E.; Johnson, P.N.T.; Manful, J.; Marti, A.; et al. Enriching gluten-free rice pasta with soybean and sweet potato flours. J. Food Sci. Technol. 2018, 55, 2641–2648. [Google Scholar] [CrossRef]
- Marti, A.; Barbiroli, A.; Marengo, M.; Fongaro, L.; Iametti, S.; Pagani, M.A. Structuring and texturing gluten-free pasta: Egg albumen or whey proteins? Eur. Food Res. Technol. 2014, 238, 217–224. [Google Scholar] [CrossRef]
- Bradauskiene, V.; Vaiciulyte-Funk, L.; Shah, B.R.; Cernauskas, D.; Tita, M.A. Recent Advances in Biotechnological Methods for Wheat Gluten Immunotoxicity Abolishment—A Review. Pol. J. Food Nutr. Sci. 2021, 71, 5–20. [Google Scholar] [CrossRef]
- Slot, I.; Bruins, D.; Van der Fels-Klerx, H.J.; Bremer, M.G.E.G.; Hamer, R.J. Immunochemical Detection Methods for Gluten in Food Products: Where Do We Go from Here? Crit. Rev. Food Sci. Nutr. 2016, 56, 2455–2466. [Google Scholar] [CrossRef]
- Denery-Papini, S.; Nicolas, Y.; Popineau, Y. Efficiency and limitations of immunochemical assays for the testing of gluten-free foods. J. Cereal Sci. 1999, 30, 124–131. [Google Scholar] [CrossRef]
- Gianfrani, C.; Mamone, G.; La Gatta, B.; Camarca, A.; Di Stasio, L.; Maurano, F.; Picascia, S.; Capozzi, V.; Perna, G.; Picariello, G.; et al. Microwave-based treatments of wheat kernels do not abolish gluten epitopes implicated in celiac disease. Food Chem. Toxicol. 2017, 101, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Rondanini, A.; Bonomi, F.; Iametti, S.; Lucisano, M.; Pagani, M.A.; Rovere, P.P. On the nature and the practical relevance of the molecular modifications induced in wheat proteins by high pressure treatment. High Press. Res. 2000, 19, 557–563. [Google Scholar] [CrossRef]
- Bonomi, F.; Iametti, S.; Mamone, G.; Ferranti, P. The Performing Protein: Beyond Wheat Proteomics? Cereal Chem. 2013, 90, 358–366. [Google Scholar] [CrossRef]
- van der Kamp, J.W.; Lupton, J. Definitions, regulations and health claims associated with dietary fibre and wholegrain foods. In Fibre-Rich and Wholegrain Foods: Improving Quality; Delcour, J.A., Poutanen, K., Eds.; Woodhead Publishing India: Old Dehli, India, 2013; Volume 237, pp. 3–24. [Google Scholar]
- Seal, C.J.; Nugent, A.P.; Tee, E.S.; Thielecke, F. Whole-grain dietary recommendations: The need for a unified global approach. Br. J. Nutr. 2016, 115, 2031–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Kamp, J.W.; Poutanen, K.; Seal, C.J.; Richardson, D.P. The HEALTHGRAIN definition of ‘whole grain’. Food Nutr. Res. 2014, 58, 22100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemery, Y.; Lullien-Pellerin, V.; Rouau, X.; Abecassis, J.; Samson, M.F.; Aman, P.; Reding, W.; Spoerndlic, C.; Barron, C. Biochemical biomarkers: Efficient tools for the assessment of wheat grain tissue proportions in milling fractions. J. Cereal Sci. 2009, 49, 55–64. [Google Scholar] [CrossRef]
- Lempereur, I.; Rouau, X.; Abecassis, J. Genetic and agronomic variation in arabinoxylan and ferulic acid contents of durum wheat (Triticum durum L.) grain and its milling fractions. J. Cereal Sci. 1997, 25, 103–110. [Google Scholar] [CrossRef]
- Chen, Y.; Ross, A.B.; Aman, P.; Kamal-Eldin, A. Alkylresorcinols as markers of whole grain wheat and rye in cereal products. J. Agric. Food Chem. 2004, 52, 8242–8246. [Google Scholar] [CrossRef]
- Nagata, Y.; Burger, M.M. Wheat-germ agglutinin—Isolation and crystallization. J. Biol. Chem. 1972, 247, 2248–2256. [Google Scholar] [CrossRef]
- Barron, C.; Samson, M.F.; Lullien-Pellerin, X.; Abecassis, V.; Rouau, X. Wheat grain tissue proportions in milling fractions using biochemical biomarker measurements: Application to different wheat cultivars. J. Cereal Sci. 2011, 53, 306–311. [Google Scholar] [CrossRef]
- Cammue, B.P.A.; Raikhel, N.V.; Peumans, W.J. Occurrence and synthesis of isolectins in different tissues of wheat. Biochem. Physiol. Pflanz. 1988, 183, 379–387. [Google Scholar] [CrossRef]
- Allen, A.K.; Desai, N.N.; Neuberger, A. Purification of glycoprotein lectin from broad bean (Vicia faba) and a comparison of its properties with lectins of similar specificity. Biochem. J. 1976, 155, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Calderón de la Barca, A.M.; Vázquez-Moreno, L.; Robles-Burgueo, M.R. Active soybean lectin in foods: Isolation and quantitation. Food Chem. 1991, 39, 321–327. [Google Scholar] [CrossRef]
- Nachbar, M.S.; Oppenheim, J.D. Lectins in the US diet: A survey of lectins in commonly consumed foods and a review of the literature. Am. J. Clin. Nutr. 1980, 33, 2338–2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoccatelli, G.; Dalla Pellegrina, C.; Vincenzi, S.; Rizzi, C.; Chignola, R.; Peruffo, A.D. Egg-matrix for large-scale single-step affinity purification of plant lectins with different carbohydrate specificities. Protein Expr. Purif. 2003, 27, 182–185. [Google Scholar] [CrossRef]
- Vincenzi, S.; Zoccatelli, G.; Perbellini, F.; Rizzi, C.; Chignola, R.; Curioni, A.; Peruffo, A.D. Quantitative determination of dietary lectin activities by enzyme-linked immunosorbent assay using specific glycoproteins immobilized on microtiter plates. J. Agric. Food Chem. 2002, 50, 6266–6270. [Google Scholar] [CrossRef]
- Matucci, A.; Veneri, G.; Pellegrin, C.D.; Zoccatelli, G.; Vincenzi, S.; Chignola, R.; Peruffo, A.D.B.; Rizzi, C. Temperature-dependent decay of wheat germ agglutinin activity and its implications for food processing and analysis. Food Control 2004, 15, 391–395. [Google Scholar] [CrossRef]
- Killilea, D.W.; McQueen, R.; Abegania, J.R. Wheat germ agglutinin is a biomarker of whole grain content in wheat flour and pasta. J. Food Sci. 2020, 85, 808–814. [Google Scholar] [CrossRef]
- Smith, J.J.; Raikhel, N.V. Production of an antibody specific for the propeptide of wheat germ agglutinin. Plant Physiol. 1989, 91, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Marti, A.; Cattaneo, S.; Buratti, S.; Parizad Abasi, P.; Masotti, F.; Iametti, S.; Pagani, M.A. Characterization of whole grain pasta: Integrating physical, chemical, molecular, and instrumental sensory approaches. J. Food Sci. 2017, 82, 2583–2590. [Google Scholar] [CrossRef]
- Jazaeri, S.; Bock, J.E.; Pavan Bagagli, M.; Iametti, S.; Bonomi, F.; Seetharaman, K. Structural evolution of gluten proteins in strong and weak wheat dough during mixing. Cereal Chem. 2015, 92, 105–113. [Google Scholar] [CrossRef]
- Bock, J.E.; West, R.; Iametti, S.; Bonomi, F.; Marengo, M.; Seetharaman, K. Gluten structural evolution during pasta processing of refined and whole grain pasta: The influence of mixing, drying, and cooking. Cereal Chem. 2015, 92, 460–465. [Google Scholar] [CrossRef]
- Bonomi, F.; D’Egidio, M.G.; Iametti, S.; Marengo, M.; Marti, A.; Pagani, M.A.; Ragg, E.M. Structure–quality relationship in commercial pasta: A molecular glimpse. Food Chem. 2012, 135, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Marengo, M.; Mamone, G.; Ferranti, P.; Polito, L.; Iametti, S.; Bonomi, F. Topological features of the intermolecular contacts in gluten-forming proteins: Exploring a novel methodological approach based on gold nanoparticles. Food Res. Int. 2019, 119, 492–498. [Google Scholar] [CrossRef]
- Azarnia, S.; Boye Joyce, I.; Mongeon, V.; Sabik, H. Detection of ovalbumin in egg white, whole egg and incurred pasta using LC-ESI-MS/MS and ELISA. Food Res. Int. 2013, 52, 526–534. [Google Scholar] [CrossRef]
- Faeste, F.K.F.; Lovberg, K.E.; Lindvik, H.; Egaas, E. Extractability, stability, and allergenicity of egg white proteins in differently heat-processed foods. J. AOAC Int. 2007, 90, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Marengo, M.; Bonomi, F.; Iametti, S.; Ferranti, P.; Barbiroli, A. Monitoring the carryover of egg proteins in pasta making to support allergen risk management. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2021, 38, 1087–1095. [Google Scholar] [CrossRef]
- Ito, K.; Yamamoto, T.; Oyama, Y.; Tsuruma, R.; Saito, E.; Saito, Y.; Ozu, T.; Honjoh, T.; Adachi, R.; Sakai, S.; et al. Food allergen analysis for processed food using a novel extraction method to eliminate harmful reagents for both ELISA and lateral-flow tests. Anal. Bioanal. Chem. 2016, 408, 5973–5984. [Google Scholar] [CrossRef]
- Iametti, S.; Donnizzelli, E.; Pittia, P.; Rovere, P.P.; Squarcina, N.; Bonomi, F. Characterization of high-pressure-treated egg albumen. J. Agric. Food Chem. 1999, 47, 3611–3616. [Google Scholar] [CrossRef]
- Iametti, S.; Donnizzelli, E.; Vecchio, G.; Rovere, P.P.; Gola, S.; Bonomi, F. Macroscopic and structural consequences of high-pressure treatment of ovalbumin solutions. J. Agric. Food Chem. 1998, 46, 3521–3527. [Google Scholar] [CrossRef]
- Ciarrocchi, M.; Boniglia, C.; Giammarioli, S.; Sanzini, E. Tutela dei Consumatori Affetti da Allergie Alimentari: Presupposti e Modalità di Intervento; Istituto Superiore di Sanità: Roma, Italy, 2011; Rapporti ISTISAN 11/27.
- Iametti, S.; Rasmussen, P.; Frøkiær, H.; Ferranti, P.; Addeo, F.; Bonomi, F. Proteolysis of bovine beta-lactoglobulin during thermal treatment in sub-denaturing conditions highlights some structural features of the temperature-modified protein and yields fragments with low immunoreactivity. Eur. J. Biochem. 2002, 269, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Iametti, S.; De Gregori, B.; Vecchio, G.; Bonomi, F. Modifications occur at different structural levels during the heat denaturation of beta-lactoglobulin. Eur. J. Biochem. 1996, 237, 106–112. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | WGA (μg/g) |
|---|---|
| whole grain semolina | 5.70 ± 0.38 |
| refined semolina | <0.025 |
| fine millings | 0.50 ± 0.06 |
| coarse millings | 0.69 ± 0.09 |
| Sample | WGA (μg/g) | Furosine (mg/g Proteins) |
|---|---|---|
| Pasta 1 * | 2.04 ± 0.14 | 3.50 ± 0.006 |
| Pasta 2 § | 0.21 ± 0.03 | 8.36 ± 0.022 |
| Time after Production Switch, min | Egg Protein Content (mg/kg) | |||
|---|---|---|---|---|
| Fresh Pasta | Dried Pasta | |||
| “R” Buffer + Detergent | “M” Buffer + Detergent + 2-ME | “R” Buffer + Detergent | “M” Buffer + Detergent + 2-ME | |
| 0 | 6.0 ± 0.7 | 13.7 ± 0.8 | 2.6 ± 0.2 | 11.1 ± 0.4 |
| 20 | 0.5 ± 0.04 | 1.4 ± 0.1 | <0.25 | 1.1 ± 0.1 |
| 45 | 0.4 ± 0.04 | 0.6 ± 0.06 | <0.25 | <0.3 |
| 60 | <0.25 | <0.3 | <0.25 | <0.3 |
| 90 | <0.25 | <0.3 | <0.25 | <0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marengo, M.; Carpen, A.; Mamone, G.; Ferranti, P.; Iametti, S. Quantification of Protein “Biomarkers” in Wheat-Based Food Systems: Dealing with Process-Related Issues. Molecules 2022, 27, 2637. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27092637
Marengo M, Carpen A, Mamone G, Ferranti P, Iametti S. Quantification of Protein “Biomarkers” in Wheat-Based Food Systems: Dealing with Process-Related Issues. Molecules. 2022; 27(9):2637. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27092637
Chicago/Turabian StyleMarengo, Mauro, Aristodemo Carpen, Gianfranco Mamone, Pasquale Ferranti, and Stefania Iametti. 2022. "Quantification of Protein “Biomarkers” in Wheat-Based Food Systems: Dealing with Process-Related Issues" Molecules 27, no. 9: 2637. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27092637