
The Introduction of Detergents in Thermal Proteome Profiling Requires Lowering the Applied Temperatures for Efficient Target Protein Identification
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
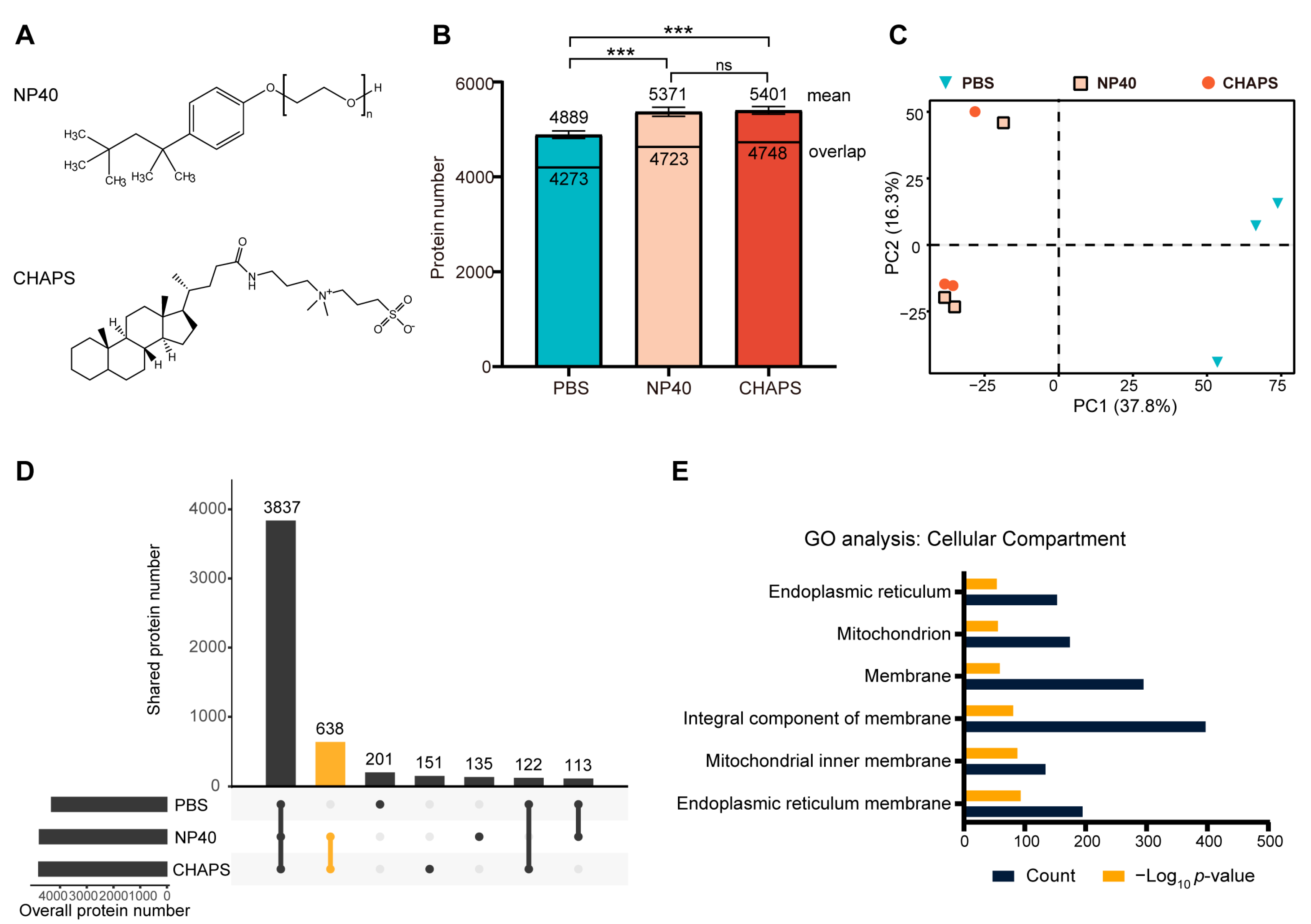
2.1. Comparing the Proteomes Extracted Using Different Detergents
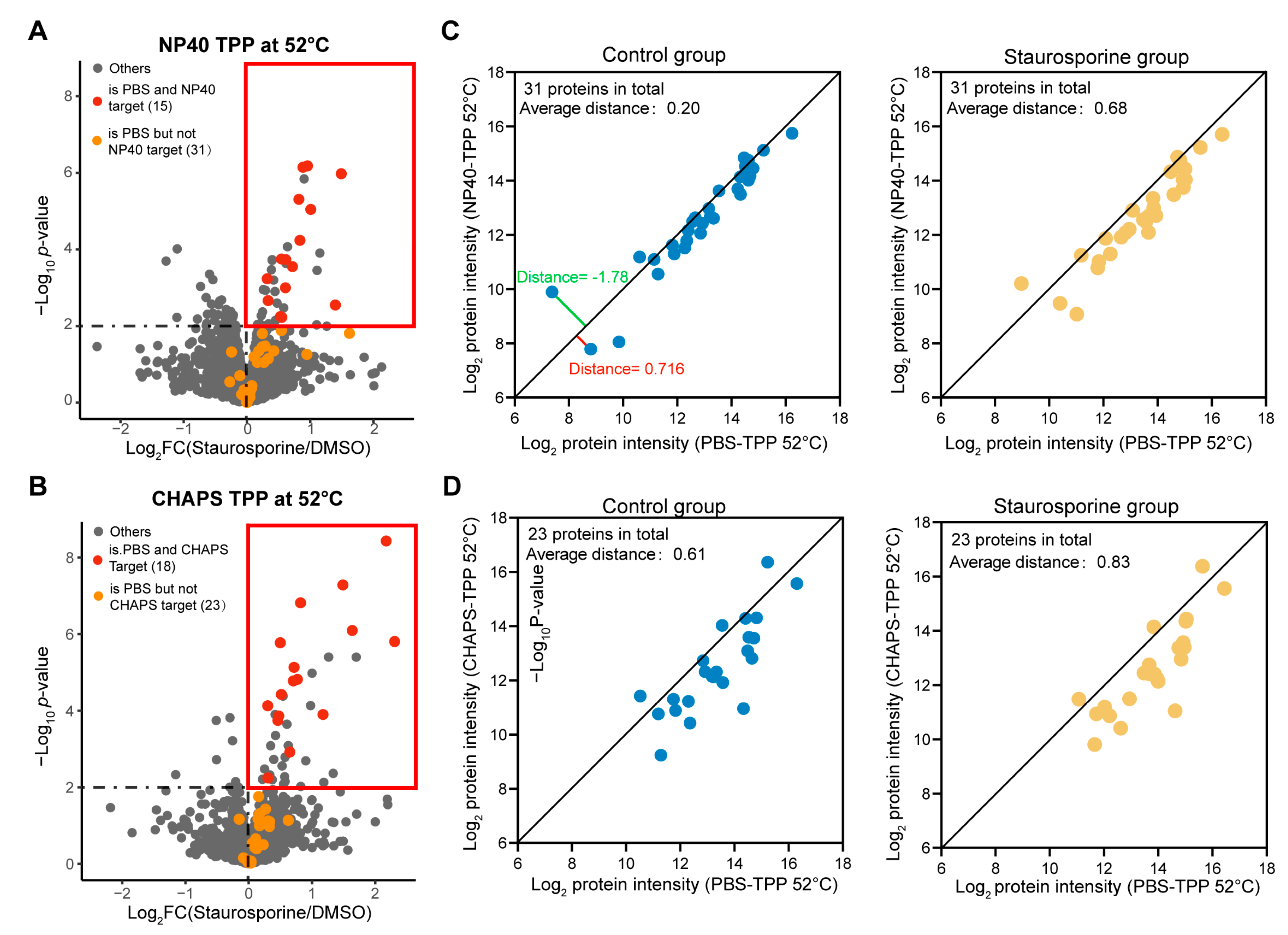
2.2. The Addition of Mild Detergents Remarkably Impaired the Target Identification Performance of TPP-52 °C
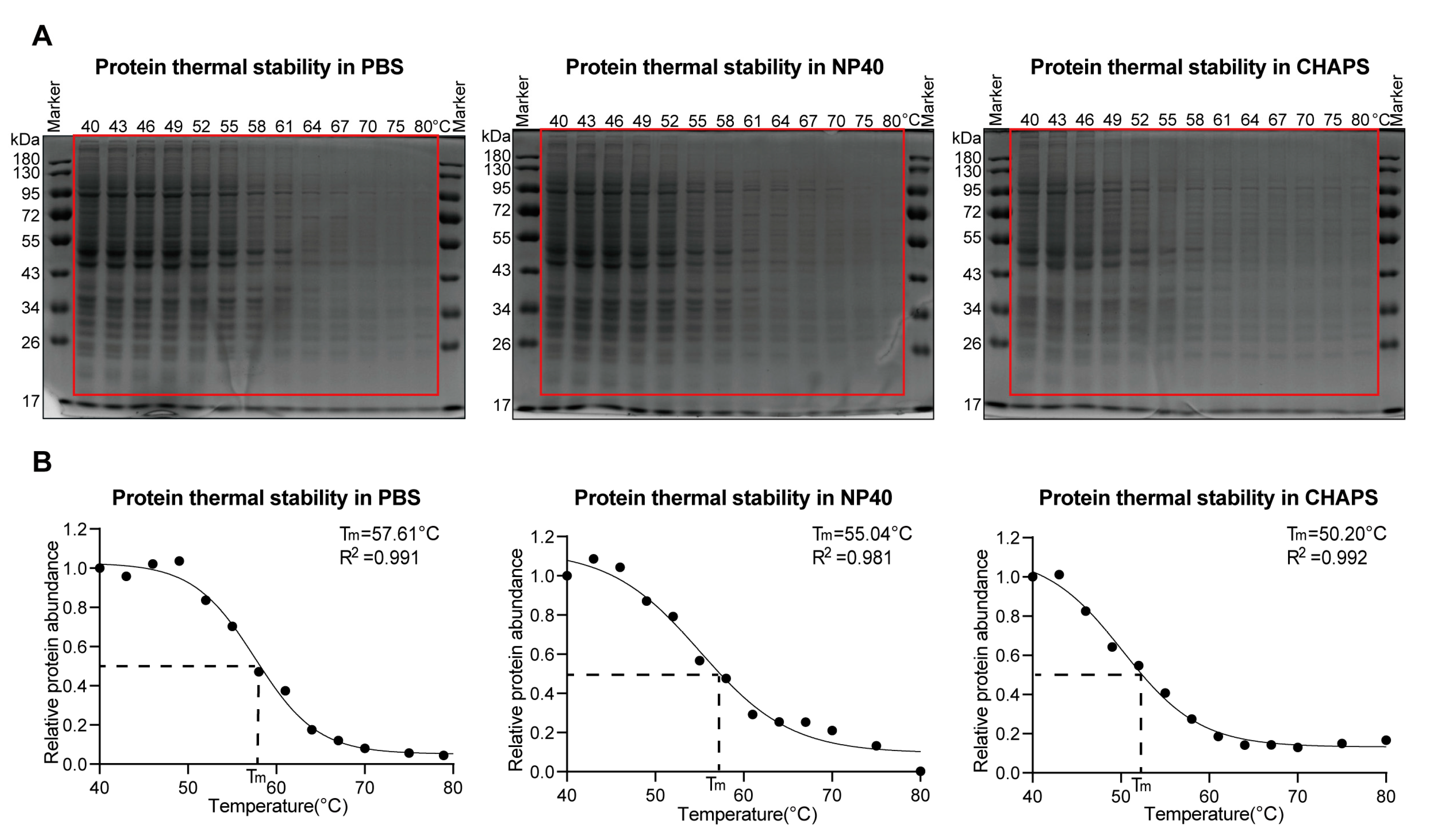
2.3. Both Non-Ionic and Zwitterionic Detergents Decrease Protein Thermal Stabilities
2.4. Using a Lower Temperature Can Restore the Target Identification Performance
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Cell Culture
4.3. Cell Lysis and Proteome Extraction
4.4. Thermal Proteome Profiling (TPP) Analysis
4.5. SDS-PAGE Analysis
4.6. LC-MS/MS Analysis
4.7. Data Processing
4.8. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Schenone, M.; Dančík, V.; Wagner, B.K.; Clemons, P.A. Target identification and mechanism of action in chemical biology and drug discovery. Nat. Chem. Biol. 2013, 9, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef]
- Klaeger, S.; Heinzlmeir, S.; Wilhelm, M.; Polzer, H.; Vick, B.; Koenig, P.A.; Reinecke, M.; Ruprecht, B.; Petzoldt, S.; Meng, C.; et al. The target landscape of clinical kinase drugs. Science 2017, 358, eaan4368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Ando, H.; Suzuki, T.; Ogura, T.; Hotta, K.; Imamura, Y.; Yamaguchi, Y.; Handa, H. Identification of a Primary Target of Thalidomide Teratogenicity. Science 2010, 327, 1345–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Kim, Y.; Kwon, H.J. Advances in identification and validation of protein targets of natural products without chemical modification. Nat. Prod. Rep. 2016, 33, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.G.; Galmozzi, A.; Wang, Y.; Correia, B.E.; Sasaki, K.; Joslyn, C.M.; Kim, A.S.; Cavallaro, C.L.; Lawrence, R.M.; Johnson, S.R.; et al. Ligand and Target Discovery by Fragment-Based Screening in Human Cells. Cell 2017, 168, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Mateus, A.; Kurzawa, N.; Becher, I.; Sridharan, S.; Helm, D.; Stein, F.; Typas, A.; Savitski, M.M. Thermal proteome profiling for interrogating protein interactions. Mol. Syst. Biol. 2020, 16, e9232. [Google Scholar] [CrossRef]
- Savitski, M.M.; Reinhard, F.B.M.; Franken, H.; Werner, T.; Savitski, M.F.; Eberhard, D.; Molina, D.M.; Jafari, R.; Dovega, R.B.; Klaeger, S.; et al. Tracking cancer drugs in living cells by thermal profiling of the proteome. Science 2014, 346, 55. [Google Scholar] [CrossRef] [Green Version]
- Lomenick, B.; Hao, R.; Jonai, N.; Chin, R.M.; Aghajan, M.; Warburton, S.; Wang, J.; Wu, R.P.; Gomez, F.; Loo, J.A.; et al. Target identification using drug affinity responsive target stability (DARTS). Proc. Natl. Acad. Sci. USA 2009, 106, 21984–21989. [Google Scholar] [CrossRef] [Green Version]
- West, G.M.; Tucker, C.L.; Xu, T.; Park, S.K.; Han, X.; Yates, J.R., 3rd; Fitzgerald, M.C. Quantitative proteomics approach for identifying protein-drug interactions in complex mixtures using protein stability measurements. Proc. Natl. Acad. Sci. USA 2010, 107, 9078–9082. [Google Scholar] [CrossRef] [Green Version]
- Piazza, I.; Kochanowski, K.; Cappelletti, V.; Fuhrer, T.; Noor, E.; Sauer, U.; Picotti, P. A Map of Protein-Metabolite Interactions Reveals Principles of Chemical Communication. Cell 2018, 172, 358–372.e.323. [Google Scholar] [CrossRef] [Green Version]
- Zinn, N.; Werner, T.; Doce, C.; Mathieson, T.; Boecker, C.; Sweetman, G.; Fufezan, C.; Bantscheff, M. Improved Proteomics-Based Drug Mechanism-of-Action Studies Using 16-Plex Isobaric Mass Tags. J. Proteome Res. 2021, 20, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Ball, K.A.; Webb, K.J.; Coleman, S.J.; Cozzolino, K.A.; Jacobsen, J.; Jones, K.R.; Stowell, M.H.B.; Old, W.M. An isothermal shift assay for proteome scale drug-target identification. Commun. Biol. 2020, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Ruan, C.; Wang, Y.; Zhang, X.; Lyu, J.; Zhang, N.; Ma, Y.; Shi, C.; Qu, G.; Ye, M. Matrix Thermal Shift Assay for Fast Construction of Multidimensional Ligand–Target Space. Anal. Chem. 2022, 94, 6482–6490. [Google Scholar] [CrossRef]
- Lyu, J.; Ruan, C.; Zhang, X.; Wang, Y.; Li, K.; Ye, M. Microparticle-Assisted Precipitation Screening Method for Robust Drug Target Identification. Anal. Chem. 2020, 92, 13912–13921. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.; Ning, W.; Liu, Z.; Zhang, X.; Fang, Z.; Li, Y.; Dang, Y.; Xue, Y.; Ye, M. Precipitate-supported thermal proteome profiling coupled with deep learning for comprehensive screening of drug target proteins. ACS Chem. Biol. 2022, 17, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Qoronfleh, M.W.; Benton, B.; Ignacio, R.; Kaboord, B. Selective enrichment of membrane proteins by partition phase separation for proteomic studies. J. Biomed. Biotechnol. 2003, 2003, 249. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, F.B.M.; Eberhard, D.; Werner, T.; Franken, H.; Childs, D.; Doce, C.; Savitski, M.F.; Huber, W.; Bantscheff, M.; Savitski, M.M.; et al. Thermal proteome profiling monitors ligand interactions with cellular membrane proteins. Nat. Methods 2015, 12, 1129–1131. [Google Scholar] [CrossRef]
- Li, J.; Van Vranken, J.G.; Pontano Vaites, L.; Schweppe, D.K.; Huttlin, E.L.; Etienne, C.; Nandhikonda, P.; Viner, R.; Robitaille, A.M.; Thompson, A.H.; et al. TMTpro reagents: A set of isobaric labeling mass tags enables simultaneous proteome-wide measurements across 16 samples. Nat. Methods 2020, 17, 399–404. [Google Scholar] [CrossRef]
- Van Vranken, J.G.; Li, J.; Mitchell, D.C.; Navarrete-Perea, J.; Gygi, S.P. Assessing target engagement using proteome-wide solvent shift assays. eLife 2021, 10, e70784. [Google Scholar] [CrossRef]
- Berlin, E.; Lizano-Fallas, V.; Carrasco del Amor, A.; Fresnedo, O.; Cristobal, S. Nonionic surfactants can modify the thermal stability of globular and membrane proteins interfering with the thermal proteome profiling principles to identify protein targets. Anal. Chem. 2023, 95, 4033–4042. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Yates, J.R. The application of mass spectrometry to membrane proteomics. Nat. Biotechnol. 2003, 21, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Huttlin, E.L.; Bruckner, R.J.; Navarrete-Perea, J.; Cannon, J.R.; Baltier, K.; Gebreab, F.; Gygi, M.P.; Thornock, A.; Zarraga, G.; Tam, S.; et al. Dual proteome-scale networks reveal cell-specific remodeling of the human interactome. Cell 2021, 184, 3022–3040.e28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Q.; Li, Y.; Ruan, C.; Wang, S.; Hu, L.; Ye, M. Solvent-induced protein precipitation for drug target discovery on the proteomic scale. Anal. Chem. 2019, 92, 1363–1371. [Google Scholar] [CrossRef]
- Lyu, J.; Wang, Y.; Ruan, C.; Zhang, X.; Li, K.; Ye, M. Mechanical stress induced protein precipitation method for drug target screening. Anal. Chim. Acta 2021, 1168, 338612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, K.; Wu, S.; Ruan, C.; Li, K.; Wang, Y.; Zhu, H.; Liu, X.; Liu, Z.; Li, G. Highly effective identification of drug targets at the proteome level by pH-dependent protein precipitation. Chem. Sci. 2022, 13, 12403–12418. [Google Scholar] [CrossRef] [PubMed]
- Callister, S.J.; Barry, R.C.; Adkins, J.N.; Johnson, E.T.; Qian, W.J.; Webb-Robertson, B.J.; Smith, R.D.; Lipton, M.S. Normalization approaches for removing systematic biases associated with mass spectrometry and label-free proteomics. J. Proteome Res. 2006, 5, 277–286. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Li, K.; Ma, Y.; Zhang, X.; Li, Y.; Yu, T.; Wang, Y.; Ye, M. The Introduction of Detergents in Thermal Proteome Profiling Requires Lowering the Applied Temperatures for Efficient Target Protein Identification. Molecules 2023, 28, 4859. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28124859
Ye Y, Li K, Ma Y, Zhang X, Li Y, Yu T, Wang Y, Ye M. The Introduction of Detergents in Thermal Proteome Profiling Requires Lowering the Applied Temperatures for Efficient Target Protein Identification. Molecules. 2023; 28(12):4859. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28124859
Chicago/Turabian StyleYe, Yuying, Kejia Li, Yanni Ma, Xiaolei Zhang, Yanan Li, Ting Yu, Yan Wang, and Mingliang Ye. 2023. "The Introduction of Detergents in Thermal Proteome Profiling Requires Lowering the Applied Temperatures for Efficient Target Protein Identification" Molecules 28, no. 12: 4859. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28124859