

Selenium Nanoparticles Control H1N1 Virus by Inhibiting Inflammatory Response and Cell Apoptosis

Abstract
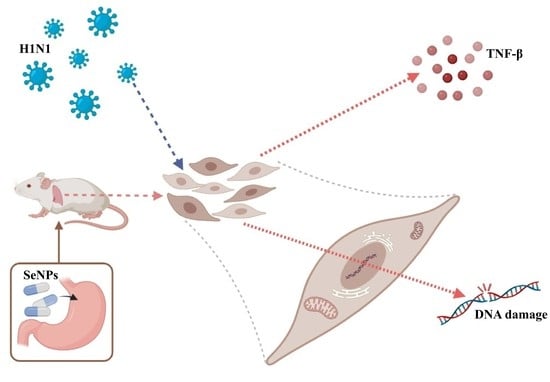
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Drug Toxicity and Antiviral Results of SeNPs
2.2. The Weight Change and Lung Inflammation in H1N1 Mice
2.3. The Inflammatory Cytokine in H1N1 Mice
2.4. Results of Apoptosis and Inflammatory Response in Mouse Lung Tissue
2.5. Immunohistochemical Detection of Mouse Lung Tissue
2.6. Selenium Levels in Tissues and Serum of Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Antiviral Effects Test
4.3. The Body Weight and the Lung Index Calculated
4.4. Detection of Cytokines
4.5. TUNEL-DAPI Co-Staining, HE Staining, and Immunohistochemistry
4.6. Determination of Selenium Content in Lung Tissue and Serum of Mice
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Krammer, F.; Smith, G.; Fouchier, R.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef]
- Honce, R.; Schultz-Cherry, S. Impact of obesity on influenza a virus pathogenesis, immune response, and evolution. Front. Immunol. 2019, 10, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Rubio, G.; Ponce-Gallegos, M.A.; Dominguez-Mazzocco, B.A.; Ponce-Gallegos, J.; Garcia-Ramirez, R.A.; Falfan-Valencia, R. Role of the host genetic susceptibility to 2009 pandemic influenza a H1N1. Viruses 2021, 13, 344. [Google Scholar] [CrossRef]
- Gaitonde, D.Y.; Moore, F.C.; Morgan, M.K. Influenza: Diagnosis and treatment. Am. Fam. Physician 2019, 100, 751–758. [Google Scholar] [PubMed]
- Nowlan, M.; Willing, E.; Turner, N. Influences and policies that affect immunisation coverage—A summary review of literature. N. Z. Med. J. 2019, 132, 79–88. [Google Scholar] [PubMed]
- Vousden, N.; Knight, M. Lessons learned from the a (H1N1) influenza pandemic. Best Pract. Res. Clin. Obstet. Gynaecol. 2021, 76, 41–52. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Gorini, F.; Sabatino, L.; Pingitore, A.; Vassalle, C. Selenium: An element of life essential for thyroid function. Molecules 2021, 26, 7084. [Google Scholar] [CrossRef]
- Gac, P.; Czerwinska, K.; Macek, P.; Jaremkow, A.; Mazur, G.; Pawlas, K.; Poreba, R. The importance of selenium and zinc deficiency in cardiovascular disorders. Environ. Toxicol. Pharmacol. 2021, 82, 103553. [Google Scholar] [CrossRef]
- Akahoshi, N.; Anan, Y.; Hashimoto, Y.; Tokoro, N.; Mizuno, R.; Hayashi, S.; Yamamoto, S.; Shimada, K.I.; Kamata, S.; Ishii, I. Dietary selenium deficiency or selenomethionine excess drastically alters organ selenium contents without altering the expression of most selenoproteins in mice. J. Nutr. Biochem. 2019, 69, 120–129. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium deficiency due to diet, pregnancy, severe illness, or COVID-19-a preventable trigger for autoimmune disease. Int. J. Mol. Sci. 2021, 22, 8532. [Google Scholar] [CrossRef] [PubMed]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prasad, K.S. Role of nano-selenium in health and environment. J. Biotechnol. 2021, 325, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Johnson, H.; Singh, A.; Sharma, A.K. Impact of selenium nanoparticles in the regulation of inflammation. Arch. Biochem. Biophys. 2022, 732, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Lin, Y.; Lin, R.; Liu, J.; Wang, H.; Hu, W.; Chen, B.; Chen, T. Traditional Chinese medicine active ingredients-based selenium nanoparticles regulate antioxidant selenoproteins for spinal cord injury treatment. J. Nanobiotechnol. 2022, 20, 278. [Google Scholar] [CrossRef]
- Ye, R.; Huang, J.; Wang, Z.; Chen, Y.; Dong, Y. Trace element selenium effectively alleviates intestinal diseases. Int. J. Mol. Sci. 2021, 22, 11708. [Google Scholar] [CrossRef]
- Zheng, Z.; Liu, L.; Zhou, K.; Ding, L.; Zeng, J.; Zhang, W. Anti-oxidant and anti-endothelial dysfunctional properties of nano-selenium in vitro and in vivo of hyperhomocysteinemic rats. Int. J. Nanomed. 2020, 15, 4501–4521. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, J.; Xu, J.F.; Pi, J. The advancing of selenium nanoparticles against infectious diseases. Front. Pharmacol. 2021, 12, 682284. [Google Scholar] [CrossRef]
- Chen, N.; Yao, P.; Zhang, W.; Zhang, Y.; Xin, N.; Wei, H.; Zhang, T.; Zhao, C. Selenium nanoparticles: Enhanced nutrition and beyond. Crit. Rev. Food Sci. Nutr. 2022, 1–12. [Google Scholar] [CrossRef]
- Lai, W.Y.; Wang, J.W.; Huang, B.T.; Lin, E.P.; Yang, P.C. A Novel TNF-α-Targeting Aptamer for TNF-α-Mediated Acute Lung Injury and Acute Liver Failure. Theranostics 2019, 9, 1741–1751. [Google Scholar] [CrossRef]
- Morris, G.; Bortolasci, C.C.; Puri, B.K.; Marx, W.; O’Neil, A.; Athan, E.; Walder, K.; Berk, M.; Olive, L.; Carvalho, A.F.; et al. The cytokine storms of COVID-19, H1N1 influenza, CRS and MAS compared. Can one sized treatment fit all? Cytokine 2021, 144, 155593. [Google Scholar] [CrossRef]
- Gu, H.; Zhu, Y.; Zhou, Y.; Huang, T.; Zhang, S.; Zhao, D.; Liu, F. LncRNA MALAT1 affects Mycoplasma pneumoniae pneumonia via NF-κB regulation. Front. Cell. Dev. Biol. 2020, 8, 563693. [Google Scholar] [CrossRef]
- Abood, R.N.; Mchugh, K.J.; Rich, H.E.; Ortiz, M.A.; Tobin, J.M.; Ramanan, K.; Robinson, K.M.; Bomberger, J.M.; Kolls, J.K.; Manni, M.L.; et al. Il-22-binding protein exacerbates influenza, bacterial super-infection. Mucosal Immunol. 2019, 12, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Jiang, S.C.; Zhang, Z.W.; Fu, Y.F.; Hu, J.; Li, Z.L. Quantification of cytokine storms during virus infections. Front. Immunol. 2021, 12, 659419. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.P.; Wang, Q.W.; Su, Y.; Gu, L.M.; Zhao, Y.; Chen, X.X.; Chen, C.; Li, W.Z.; Wang, G.F.; Li, K.S. Emodin Inhibition of Influenza A Virus Replication and Influenza Viral Pneumonia via the Nrf2, TLR4, p38/JNK and NF-kappaB Pathways. Molecules 2017, 22, 1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Han, Q.; Wang, L.; Wang, B.; Chen, J.; Cai, B.; Wu, C.; Zhu, X.; Liu, F.; Han, D.; et al. Jinhua Qinggan granules attenuates acute lung injury by promotion of neutrophil apoptosis and inhibition of TLR4/MyD88/NF-κB pathway. J. Ethnopharmacol. 2023, 301, 2–10. [Google Scholar] [CrossRef]
- Liu, X.; Chen, D.; Su, J.; Zheng, R.; Ning, Z.; Zhao, M.; Zhu, B.; Li, Y. Selenium nanoparticles inhibited H1N1 influenza virus-induced apoptosis by ROS-mediated signaling pathways. RSC Adv. 2022, 12, 3862–3870. [Google Scholar] [CrossRef]
- Ling, L.J.; Lu, Y.; Zhang, Y.Y.; Zhu, H.Y.; Tu, P.; Li, H.; Chen, D.F. Flavonoids from houttuynia cordata attenuate H1N1-induced acute lung injury in mice via inhibition of influenza virus and toll-like receptor signalling. Phytomedicine 2020, 67, 2–39. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Hsu, A.C.; Pang, Z.; Pan, H.; Zuo, X.; Wang, G.; Zheng, J.; Wang, F. Role of the innate cytokine storm induced by the influenza A virus. Viral Immunol. 2019, 32, 244–251. [Google Scholar] [CrossRef]
- Chen, D.; Ning, Z.; Su, J.; Zheng, R.; Liu, X.; Wu, H.L.; Zhu, B.; Li, Y. Inhibition of H1N1 by Picochlorum sp. 122 via AKT and p53 signaling pathways. Food Sci. Nutr. 2023, 11, 743–751. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Gong, G.; Xia, Y.; Wang, C.; Chen, Y.; Hua, L.; Zhong, J.; Tang, Y.; Liu, X.; et al. Restriction of H1N1 influenza virus infection by selenium nanoparticles loaded with ribavirin via resisting caspase-3 apoptotic pathway. Int. J. Nanomed. 2018, 13, 5787–5797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, D.; Su, J.; Chen, M.; Chen, T.; Jia, W.; Zhu, B. Selenium-ruthenium complex blocks H1N1 influenza virus-induced cell damage by activating GPx1/TrxR1. Theranostics 2023, 13, 1843–1859. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. Optimal nutritional status for a well-functioning immune system is an important factor to protect against viral infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullahi, H.; Elnahas, A.; Konje, J.C. Seasonal influenza during pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 258, 235–239. [Google Scholar] [CrossRef]
- Li, X.; Xu, Y.; Li, W.; Che, J.; Zhao, X.; Cao, R.; Li, X.; Li, S. Design, synthesis, biological evaluation, and molecular dynamics simulation of influenza polymerase PB2 inhibitors. Molecules 2023, 28, 1849. [Google Scholar] [CrossRef]
- Wu, Y.; Min, J.; Ge, C.; Shu, J.; Tian, D.; Yuan, Y.; Zhou, D. Interleukin 22 in liver injury, inflammation and cancer. Int. J. Biol. Sci. 2020, 16, 2405–2413. [Google Scholar] [CrossRef]
- Bakshi, H.A.; Quinn, G.A.; Nasef, M.M.; Mishra, V.; Aljabali, A.; El-Tanani, M.; Serrano-Aroca, A.; Webba, D.S.M.; Mccarron, P.A.; Tambuwala, M.M. Crocin inhibits angiogenesis and metastasis in colon cancer via TNF-α/NF-kB/VEGF pathways. Cells 2022, 11, 1502. [Google Scholar] [CrossRef]
- Gu, Y.; Zuo, X.; Zhang, S.; Ouyang, Z.; Jiang, S.; Wang, F.; Wang, G. The mechanism behind influenza virus cytokine storm. Viruses 2021, 13, 1362. [Google Scholar] [CrossRef]
- Basavaraju, S.; Mishra, S.; Jindal, R.; Kesavardhana, S. Emerging Role of ZBP1 in Z-RNA Sensing, Influenza Virus-Induced Cell Death, and Pulmonary Inflammation. mBio 2022, 13, e00401-22. [Google Scholar] [CrossRef]
- Zinatizadeh, M.R.; Schock, B.; Chalbatani, G.M.; Zarandi, P.K.; Jalali, S.A.; Miri, S.R. The Nuclear Factor Kappa B (NF-kB) signaling in cancer development and immune diseases. Genes Dis. 2021, 8, 287–297. [Google Scholar] [CrossRef]
- Catz, S.D.; Johnson, J.L. Transcriptional regulation of BCL-2 by nuclear factor Kappa B and its significance in prostate cancer. Oncogene 2001, 20, 7342–7351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; Lopez, J.M. Understanding MAPK signaling pathways in apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef]
- Haselager, M.V.; Thijssen, R.; Bax, D.; Both, D.; De Boer, F.; Mackay, S.; Dubois, J.; Mellink, C.; Kater, A.P.; Eldering, E. JAK–STAT signalling shapes the NF-κB response in CLL towards venetoclax sensitivity or resistance via Bcl-XL. Mol. Oncol. 2023, 17, 1112–1128. [Google Scholar] [CrossRef]
- Butturini, E.; Carcereri, D.P.A.; Mariotto, S. Redox regulation of STAT1 and STAT3 signaling. Int. J. Mol. Sci. 2020, 21, 7034. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: Its role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Chen, D.; Zheng, R.; Su, J.; Lai, J.; Chen, H.; Ning, Z.; Liu, X.; Zhu, B.; Li, Y. Inhibition of H1N1 Influenza Virus-induced Apoptosis by Ebselen Through ROS-mediated ATM/ATR Signaling Pathways. Biol. Trace Elem. Res. 2023, 201, 2811–2822. [Google Scholar] [CrossRef]
- Xu, T.; Lai, J.; Su, J.; Chen, D.; Zhao, M.; Li, Y.; Zhu, B. Inhibition of H3N2 Influenza Virus Induced Apoptosis by Selenium Nanoparticles with Chitosan through ROS-Mediated Signaling Pathways. ACS Omega 2023, 8, 8473–8480. [Google Scholar] [CrossRef]
- Su, J.; Chen, D.; Zheng, R.; Liu, X.; Zhao, M.; Zhu, B.; Li, Y. Duvira Antarctic polysaccharide inhibited H1N1 influenza virus-induced apoptosis through ROS mediated ERK and STAT-3 signaling pathway. Mol. Biol. Rep. 2022, 49, 6225–6233. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Lai, J.; Li, J.; Li, C.; Liu, X.; Wang, C.; Zhu, B.; Li, Y. Selenium Nanoparticles Control H1N1 Virus by Inhibiting Inflammatory Response and Cell Apoptosis. Molecules 2023, 28, 5920. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28155920
Su J, Lai J, Li J, Li C, Liu X, Wang C, Zhu B, Li Y. Selenium Nanoparticles Control H1N1 Virus by Inhibiting Inflammatory Response and Cell Apoptosis. Molecules. 2023; 28(15):5920. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28155920
Chicago/Turabian StyleSu, Jingyao, Jia Lai, Jiali Li, Chuqing Li, Xia Liu, Chenyang Wang, Bing Zhu, and Yinghua Li. 2023. "Selenium Nanoparticles Control H1N1 Virus by Inhibiting Inflammatory Response and Cell Apoptosis" Molecules 28, no. 15: 5920. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28155920