


Potential of Tamarind Shell Extract against Oxidative Stress In Vivo and In Vitro
 ,
,
Abstract
:1. Introduction
2. Results
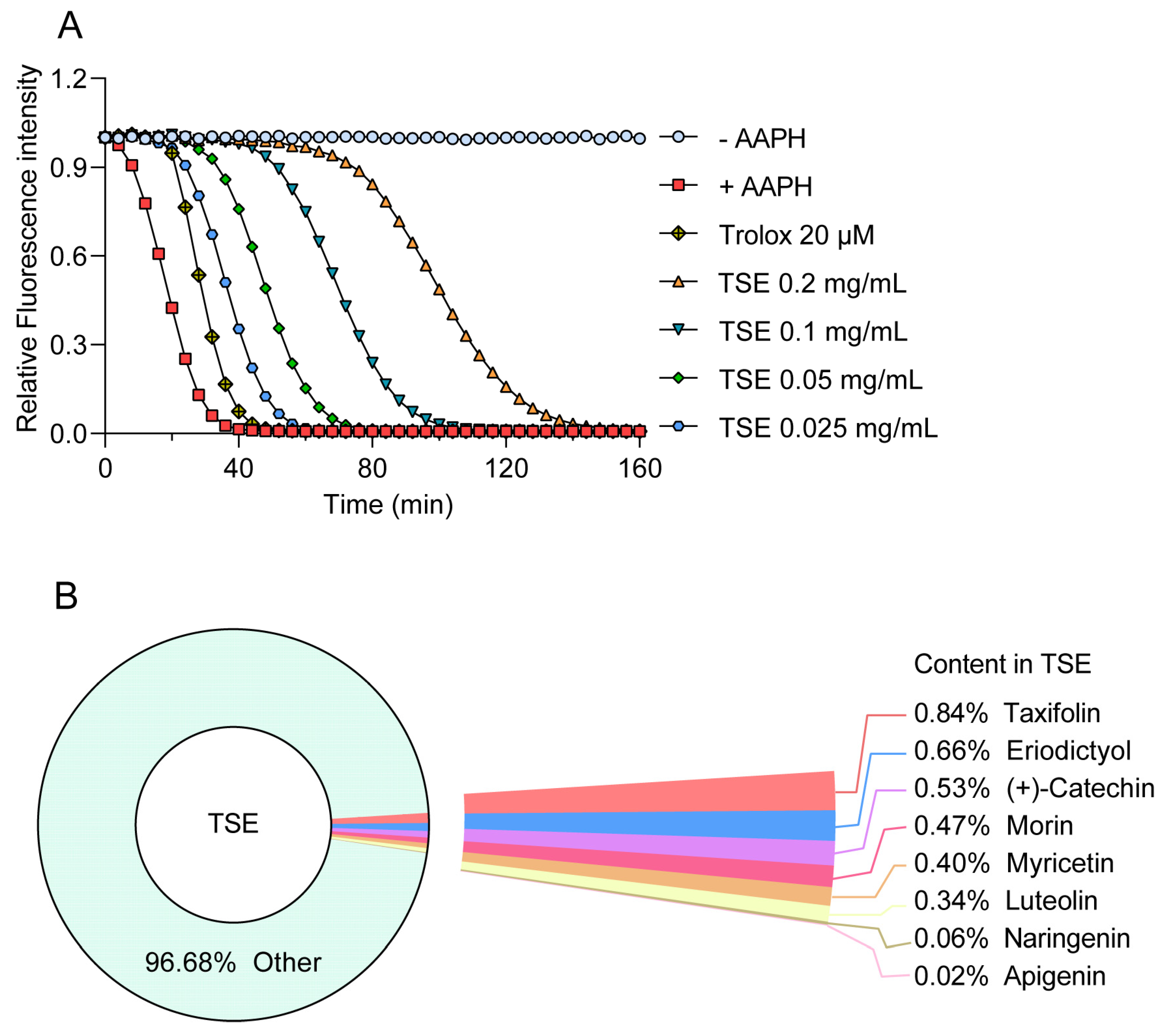
2.1. Quantitative Analysis of the Primary Components in TSE
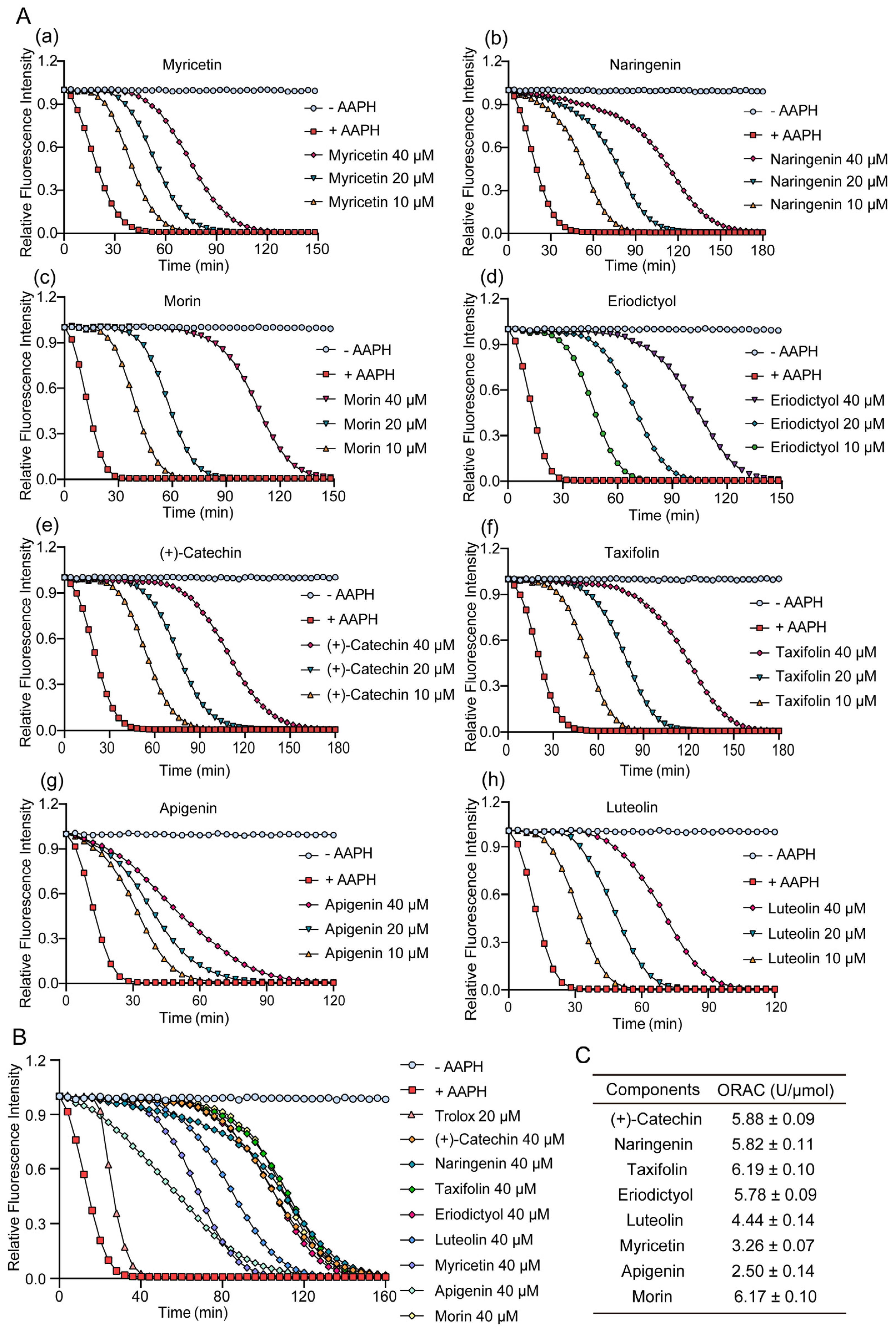
2.2. Antioxidative Capacity of TSE and Its Primary Components Evaluated by ORAC
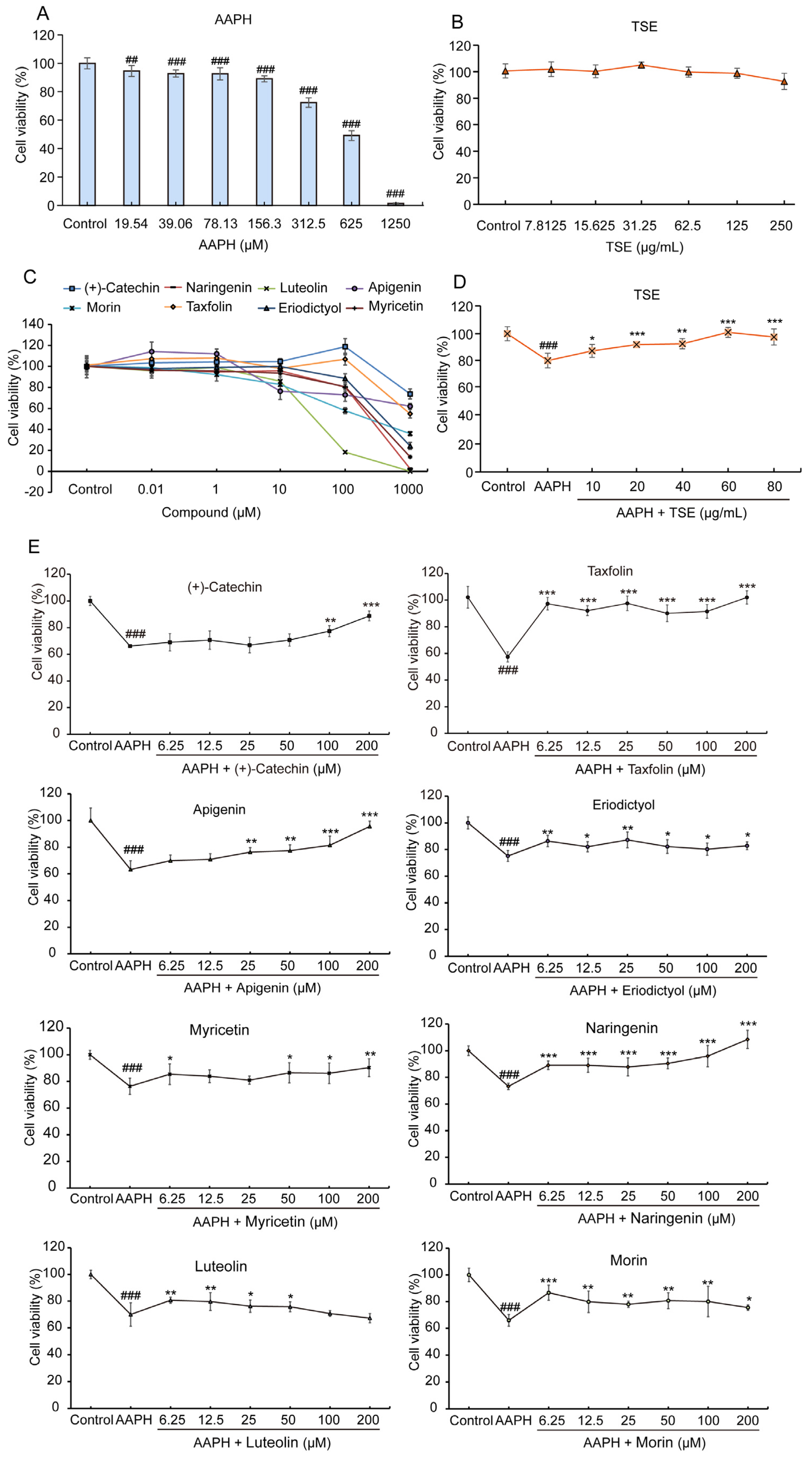
2.3. Cytoprotective Effects of TSE and the Individual Flavonoid inAAPH-Treated ATDC5 Cells
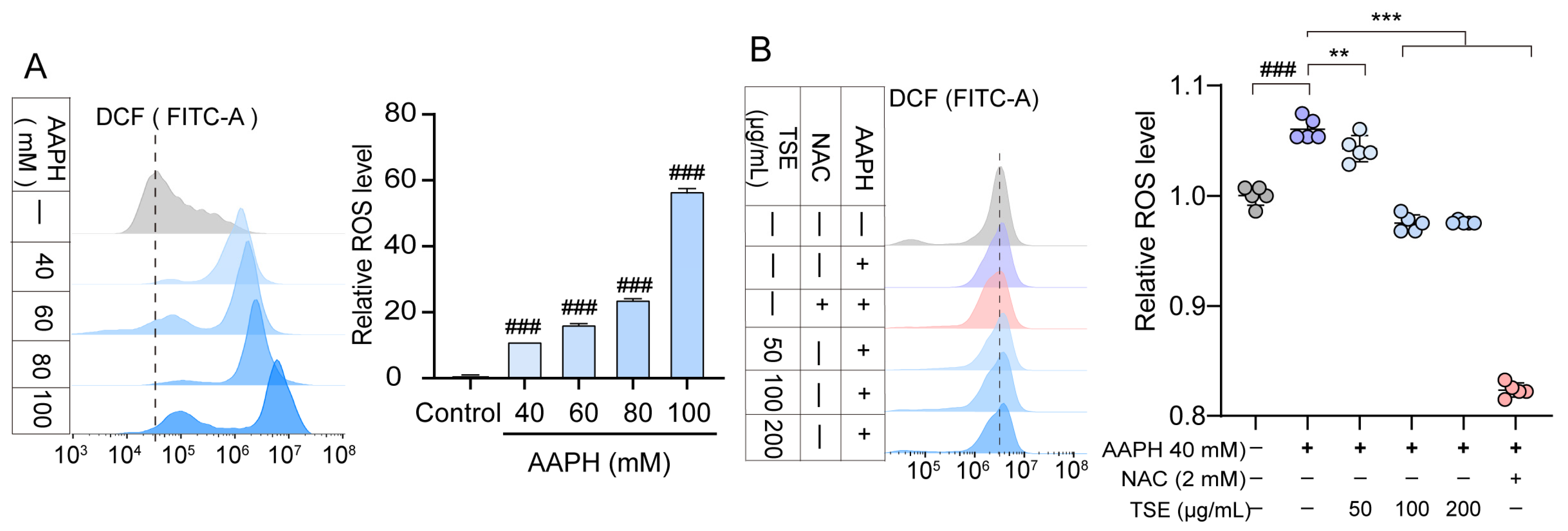
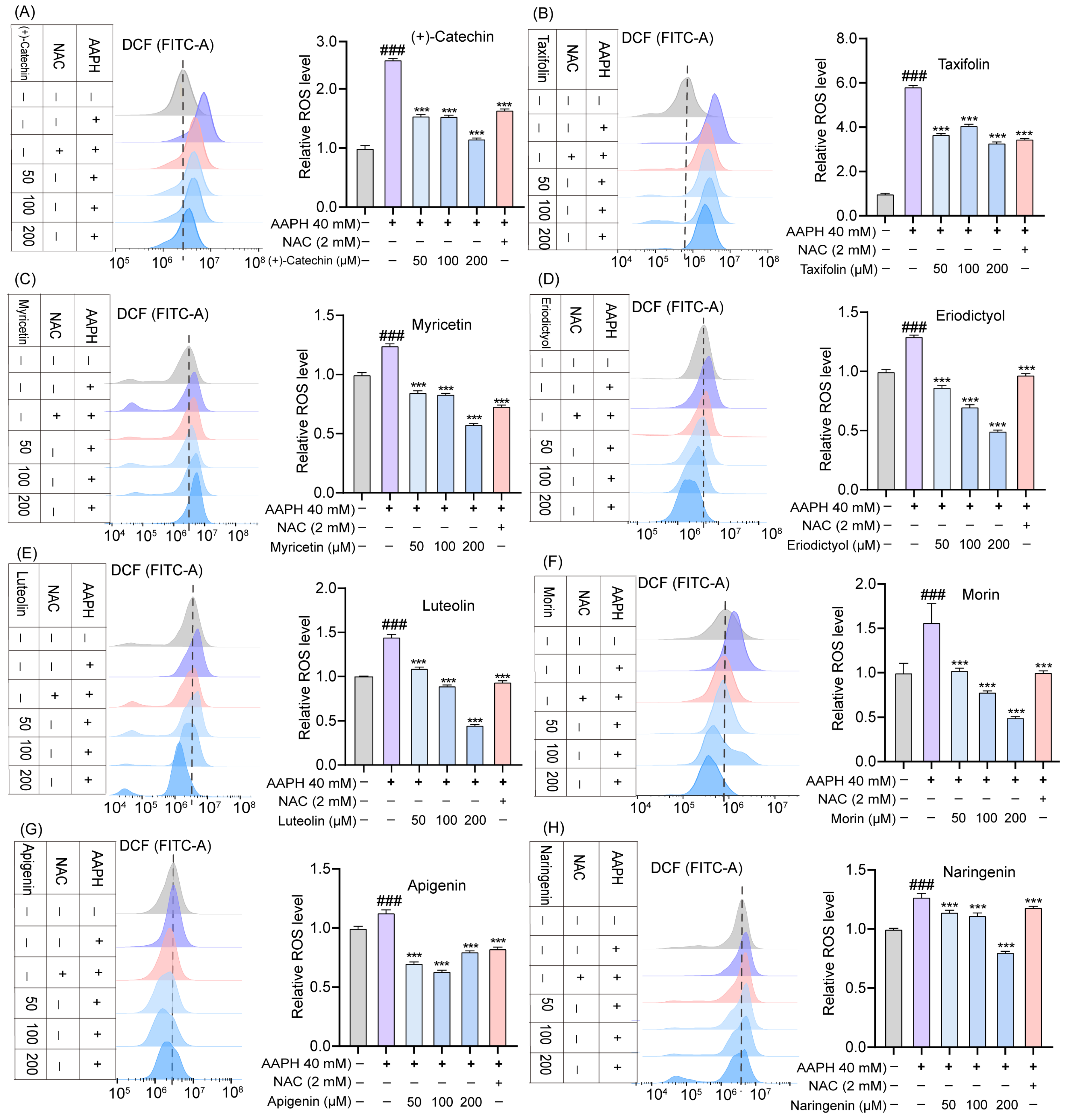
2.4. Effects of TSE and Flavonoids on ROS Level in AAPH-Treated ATDC5 Cells
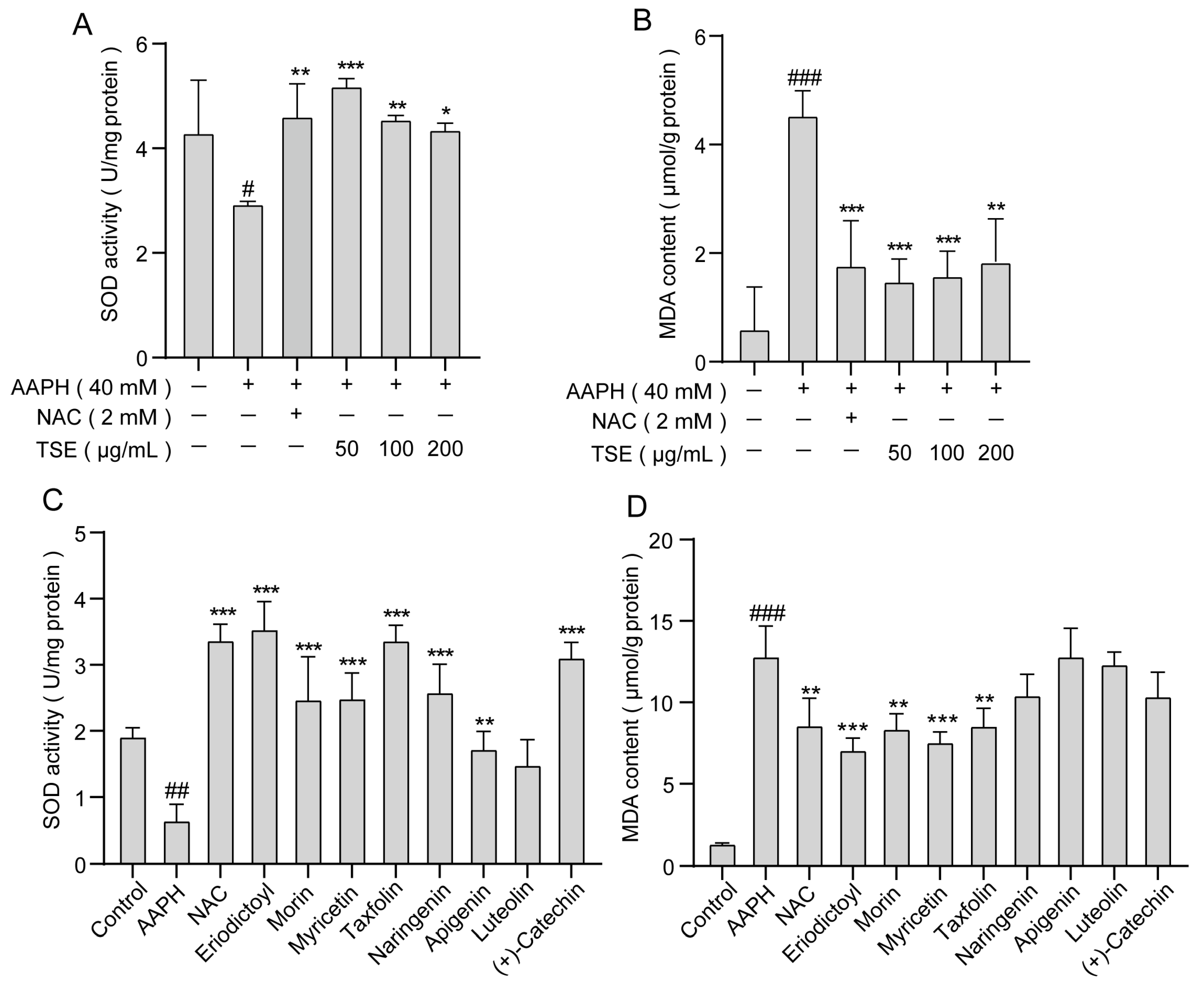
2.5. Effects of TSE and the 8 Flavonoids on Intracellular MDA Content and SOD Activity in AAPH-Treated ATDC5 Cells
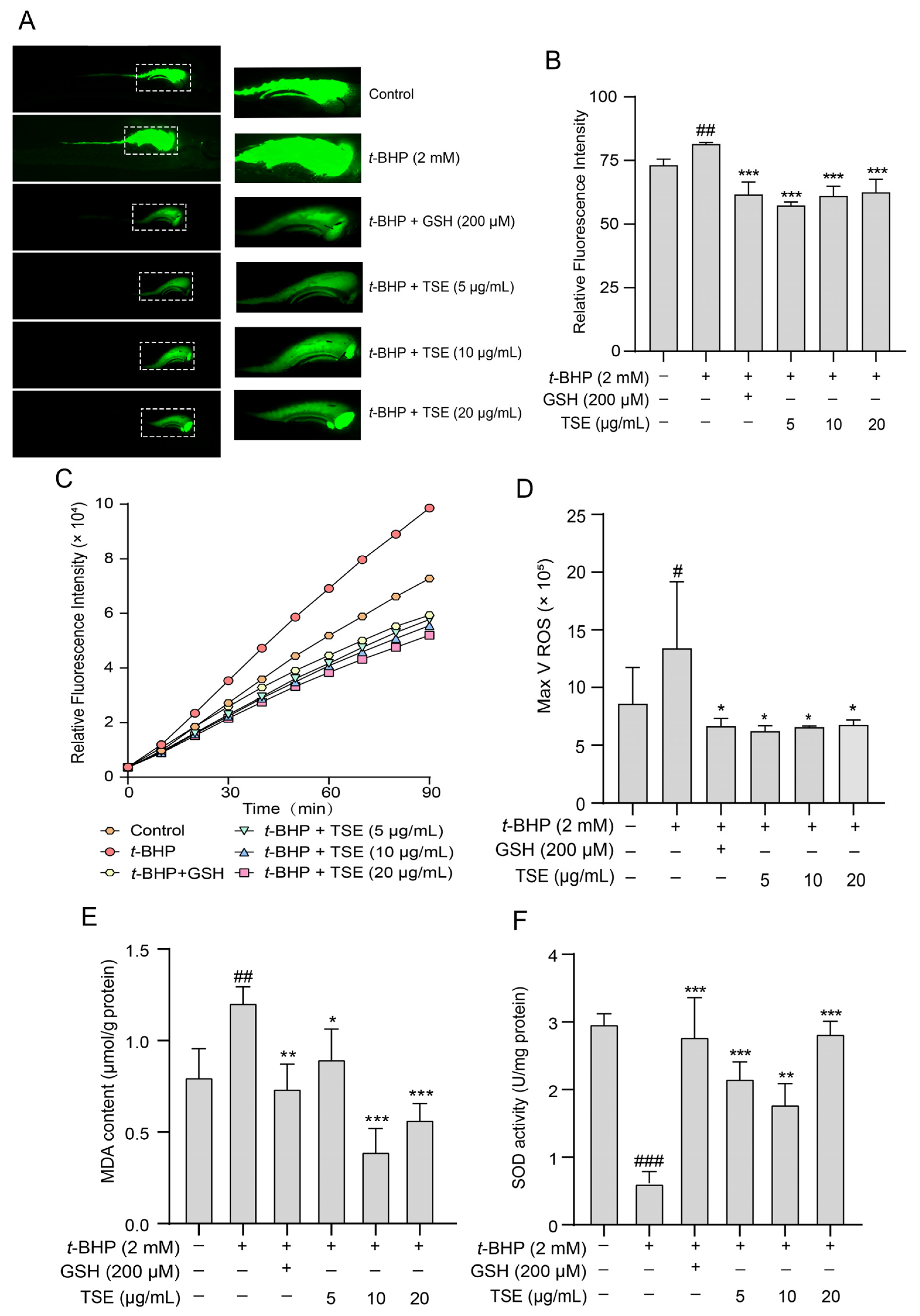
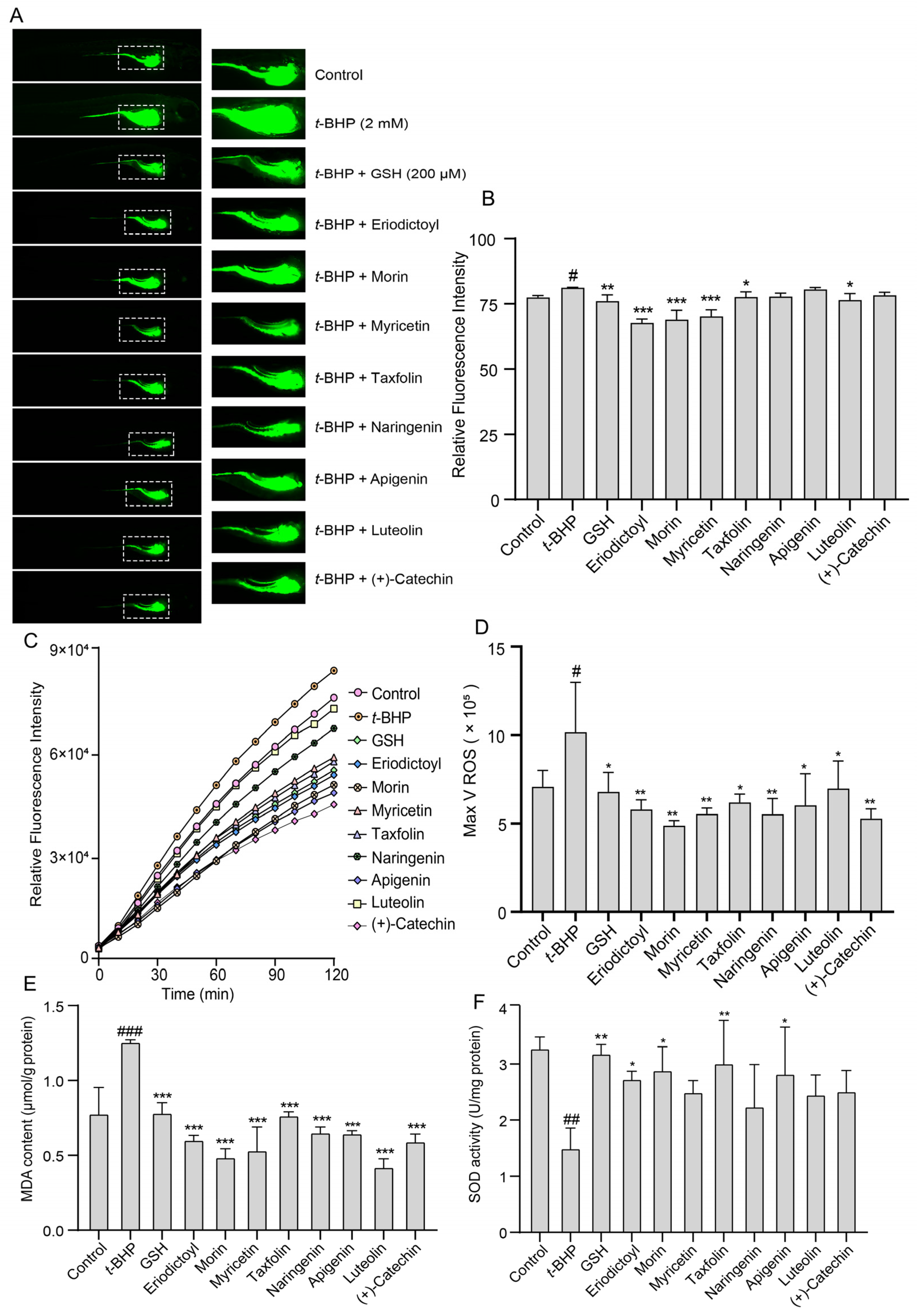
2.6. Effects of TSE and Its Components on t-BHP-Induced Oxidative Stress in Zebrafish
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Plant Materials
4.1.2. Chemicals
4.1.3. Zebrafish and Husbandry
4.2. Methods
4.2.1. Preparing the Tamarind Shell Extract (TSE)
4.2.2. Quantitative Analysis of Flavonoids in TSE
4.2.3. ORAC Assay
4.2.4. Cell Culture
4.2.5. MTT Assay
4.2.6. Measurement of Intracellular Antioxidant Capacity
4.2.7. Measurement of SOD Activity and MDA Content in ATDC5 Cells and Zebrafish
4.2.8. Waterborne Exposure of Zebrafish
4.2.9. Measurement of the Intracellular ROS Generation and Image Analysis in Zebrafish
4.2.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, E.G.; Cotter, T.G. New Insight into the Role of Reactive Oxygen Species (ROS) in Cellular Signal-Transduction Processes. Mol. Biol. 2015, 319, 221–254. [Google Scholar]
- Ye, R.; Shi, M.; Liu, Q.; Chen, J. Redox Imbalance and Stroke. Oxidative Med. Cell. Longev. 2016, 2016, 3065263. [Google Scholar] [CrossRef]
- Dossena, S.; Marino, A. Cellular Oxidative Stress. Antioxidants 2021, 10, 399. [Google Scholar] [CrossRef]
- Singh, K.K. Oxidative Stress, Disease and Cancer; Imperial College Press: London, UK, 2006. [Google Scholar]
- Shim, S.Y.; Kim, H.S. Oxidative stress and the antioxidant enzyme system in the developing brain. Korean J. Pediatr. 2013, 56, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Li, W.; Wang, W.; Yang, B.; Zhang, L.; Su, W.; Wang, W. Current research status on chemical components and biological activities of Tamarindus indica Linn. J. Int. Pharm. Res. 2016, 43, 697–704. [Google Scholar]
- Arshad, M.S.; Imran, M.; Ahmed, A.; Sohaib, M.; Ullah, A.; Nisa, M.U.; Hina, G.; Khalid, W.; Rehana, H. Tamarind: A diet-based strategy against lifestyle maladies. Food Sci. Nutr. 2019, 7, 3378–3390. [Google Scholar] [CrossRef] [PubMed]
- Dal, J.-J.; Yuan, L.-C.; Ll, M.-F.; Liu, H.-G.; Luo, H.-Y.; Zhao, Q.-L.; Sha, Y.-C. Reserch Overview of Tamarind Application on Food. Food Res. Dev. 2015, 36, 17–20. [Google Scholar]
- Adeola, A.-A.; Aworh, O.-C. Sugar and dietary fibre components of tamarind (Tamarindus indica L.) fruits from Nigeria. Niger. Food J. 2010, 28, 32–40. [Google Scholar] [CrossRef]
- Shao, J.-L.; Li, Q.-W.; Liu, H.-C.; Du, L.-J.; Yang, F.; Shu, J.-H. Studies on the processing technique of tamarndus indica L. health—Jelly drops. China Food Addit. 2010, 2, 201–205. [Google Scholar]
- Sadik, H.A. The Nutritional Value of “Poha Beer” (Tamarind Fruit Drink) and its Social Usage in Tamale Metropolis. Pak. J. Nutr. 2010, 9, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Razali, N.; Mat Junit, S.; Ariffin, A.; Ramli, N.S.; Abdul Aziz, A. Polyphenols from the extract and fraction of T. indica seeds protected HepG2 cells against oxidative stress. BMC Complement. Altern. Med. 2015, 15, 438. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Malviya, R.; Sharma, P.K. Extraction and Characterization of Tamarind Seed Polysaccharide as a Pharmaceutical Excipient. Pharmacogn. J. 2011, 3, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Remigante, A.; Spinelli, S.; Straface, E.; Gambardella, L.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Antioxidant Activity of Quercetin in a H2O2-Induced Oxidative Stress Model in Red Blood Cells: Functional Role of Band 3 Protein. Int. J. Mol. Sci. 2022, 23, 10991. [Google Scholar] [CrossRef]
- Zhou, D.D.; Luo, M.; Huang, S.Y.; Saimaiti, A.; Shang, A.; Gan, R.Y.; Li, H.B. Effects and Mechanisms of Resveratrol on Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary Anti-Aging Polyphenols and Potential Mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Chawananorasest, K.; Saengtongdee, P.; Kaemchantuek, P. Extraction and Characterization of Tamarind (Tamarind indica L.) Seed Polysaccharides (TSP) from Three Difference Sources. Molecules 2016, 21, 775. [Google Scholar] [CrossRef]
- Nakchat, O.; Nalinratana, N.; Meksuriyen, D.; Pongsamart, S. Tamarind seed coat extract restores reactive oxygen species through attenuation of glutathione level and antioxidant enzyme expression in human skin fibroblasts in response to oxidative stress. Asian Pac. J. Trop. Biomed. 2014, 4, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Lima, Z.M.; da Trindade, L.S.; Santana, G.C.; Padilha, F.F.; da Costa Mendonça, M.; da Costa, L.P.; López, J.A.; Macedo, M.L.H. Effect of Tamarindus indica L. and Manihot esculenta Extracts on Antibiotic-resistant Bacteria. Pharmacogn. Res. 2017, 9, 195–199. [Google Scholar] [CrossRef]
- Li, H.; Cui, X. Extraction of flavonoids and scavenging effect on hydroxyl radicals from tamarind shells. J. South. Agric. 2014, 45, 844–849. [Google Scholar]
- Zhuang, J.Y.; Li, B.S.; Wang, S.Y. Study on the Anti-diabetic Activity of Tamarindus pericarp. Mod. Food Sci. Technol. 2011, 27, 762, 773–776. [Google Scholar] [CrossRef]
- Huang, R.-P.; Ding, Z.-J.; Han, S.-C.; Li, X.-Y.; Zhang, Q.-Y.; Li, W.-X. Study on the Hypoglycemic Activity of Tamarindus Shell Extract. Pharm. Today 2022, 32, 1–15. [Google Scholar]
- Dziubla, T.; Butterfield, D.A. Oxidative Stress and Biomaterials; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Garrett, A.R.; Murray, B.K.; Robison, R.A.; O’Neill, K.L. Measuring antioxidant capacity using the ORAC and TOSC assays. Adv. Protoc. Oxidative Stress II 2010, 594, 251–262. [Google Scholar]
- Alkadi, H. A Review on Free Radicals and Antioxidants. Infect. Disord. Drug Targets 2020, 20, 16–26. [Google Scholar] [CrossRef]
- Tekin, K.; Tekin, M.I. Oxidative stress and diabetic retinopathy. In Pathology; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Pedro, A.C.; Paniz, O.G.; Fernandes, I.A.A.; Bortolini, D.G.; Rubio, F.T.V.; Haminiuk, C.W.I.; Maciel, G.M.; Magalhães, W.L.E. The Importance of Antioxidant Biomaterials in Human Health and Technological Innovation: A Review. Antioxidants 2022, 11, 1644. [Google Scholar] [CrossRef]
- Abbate, F.; Maugeri, A.; Laurà, R.; Levanti, M.; Navarra, M.; Cirmi, S.; Germanà, A. Zebrafish as a Useful Model to Study Oxidative Stress-Linked Disorders: Focus on Flavonoids. Antioxidants 2021, 10, 668. [Google Scholar] [CrossRef]
- Cecerska-Heryć, E.; Surowska, O.; Heryć, R.; Serwin, N.; Napiontek-Balińska, S.; Dołęgowska, B. Are antioxidant enzymes essential markers in the diagnosis and monitoring of cancer patients—A review. Clin. Biochem. 2021, 93, 1–8. [Google Scholar] [CrossRef]
- de Souza Farias, S.A.; da Costa, K.S.; Martins, J.B.L. Analysis of Conformational, Structural, Magnetic, and Electronic Properties Related to Antioxidant Activity: Revisiting Flavan, Anthocyanidin, Flavanone, Flavonol, Isoflavone, Flavone, and Flavan-3-ol. ACS Omega 2021, 6, 8908–8918. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Gong, H.; Mei, H.; Shi, L.; Yu, J.; Hu, Y. Taxifolin Targets PI3K and mTOR and Inhibits Glioblastoma Multiforme. J. Oncol. 2021, 2021, 5560915. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, R.C.; Yang, Z.H.; Sun, G.B.; Wang, M.; Ma, X.J.; Yang, L.J.; Sun, X.B. Taxifolin prevents diabetic cardiomyopathy in vivo and in vitro by inhibition of oxidative stress and cell apoptosis. Food Chem. Toxicol. 2014, 63, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Cho, Y.Y.; Yao, K.; Nadas, J.; Kim, D.J.; Cho, E.J.; Lee, M.H.; Pugliese, A.; Zhang, J.; Bode, A.M.; et al. Eriodictyol inhibits RSK2-ATF1 signaling and suppresses EGF-induced neoplastic cell transformation. J. Biol. Chem. 2011, 286, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Fu, H.; Li, C.; Deng, Z.; Chang, H. Eriodictyol inhibits breast carcinogenesis by targeting circ_0007503 and repressing PI3K/Akt pathway. Phytomed. Int. J. Phytother. Phytopharm. 2022, 102, 154159. [Google Scholar] [CrossRef]
- Jing, X.; Shi, H.; Zhu, X.; Wei, X.; Ren, M.; Han, M.; Ren, D.; Lou, H. Eriodictyol Attenuates β-Amyloid 25–35 Peptide-Induced Oxidative Cell Death in Primary Cultured Neurons by Activation of Nrf2. Neurochem. Res. 2015, 40, 1463–1471. [Google Scholar] [CrossRef]
- Li, L.; Li, W.J.; Zheng, X.R.; Liu, Q.L.; Du, Q.; Lai, Y.J.; Liu, S.Q. Eriodictyol ameliorates cognitive dysfunction in APP/PS1 mice by inhibiting ferroptosis via vitamin D receptor-mediated Nrf2 activation. Mol. Med. 2022, 28, 11. [Google Scholar] [CrossRef]
- Maquera-Huacho, P.M.; Spolidorio, D.P.; Manthey, J.A.; Grenier, D. Eriodictyol Suppresses Porphyromonas gingivalis-Induced Reactive Oxygen Species Production by Gingival Keratinocytes and the Inflammatory Response of Macrophages. Front. Oral Health 2022, 3, 847914. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Chen, Y.; Zhou, B.; Shan, X.; Yang, G. Eriodictyol inhibits IL-1β-induced inflammatory response in human osteoarthritis chondrocytes. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 107, 1128–1134. [Google Scholar] [CrossRef]
- Sroka, Z.; Żbikowska, B.; Hładyszowski, J. The antiradical activity of some selected flavones and flavonols. Experimental and quantum mechanical study. J. Mol. Model. 2015, 21, 307. [Google Scholar] [CrossRef]
- Xu, J.; Yao, X.; Hiroshi, K. Oxygen radical absorbance capacity assay and its application. Chin. Pharmacol. Bull. 2006, 8, 7. [Google Scholar]
- Kuang, X.-P.; Huang, R.-P.; Han, S.-C.; Zhang, Q.-Y.; Yan, C.-Y.; Li, W.-X. Comparison of Antioxidant Activity of Several Animal Protein-derived Peptides by ORAC. Guangdong Chem. Ind. 2022, 49, 14–15, 52. [Google Scholar]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Xu, N.W.; Gao, G.M.; Ni, S.; Miao, K.S.; Li, C.K.; Wang, L.M.; Xie, H.G. Polysaccharide from Angelica sinensis protects chondrocytes from H2O2-induced apoptosis through its antioxidant effects in vitro. Int. J. Biol. Macromol. 2016, 87, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gong, K.; Mao, X.; Li, W. Tetrandrine induces apoptosis by activating reactive oxygen species and repressing Akt activity in human hepatocellular carcinoma. Int. J. Cancer 2011, 129, 1519–1531. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Hu, N.; Ding, D.; Zhao, W.; Feng, Y.; Zhang, H.; Li, G.; Wang, Y. Developmental toxicity and oxidative stress induced by gamma irradiation in zebrafish embryos. Radiat. Environ. Biophys. 2016, 55, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lackmann, C.; Santos, M.M.; Rainieri, S.; Barranco, A.; Hollert, H.; Spirhanzlova, P.; Velki, M.; Seiler, T.B. Novel procedures for whole organism detection and quantification of fluorescence as a measurement for oxidative stress in zebrafish (Danio rerio) larvae. Chemosphere 2018, 197, 200–209. [Google Scholar] [CrossRef]
- Mugoni, V.; Camporeale, A.; Santoro, M.M. Analysis of oxidative stress in zebrafish embryos. J. Vis. Exp. JoVE 2014, 89, e51328. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.Y.; Kim, E.A.; Kang, S.I.; Yang, H.W.; Ryu, B.; Wang, L.; Lee, J.S.; Jeon, Y.J. Protective Effects of Fucoidan Isolated from Celluclast-Assisted Extract of Undaria pinnatifida Sporophylls against AAPH-Induced Oxidative Stress In Vitro and In Vivo Zebrafish Model. Molecules 2020, 25, 2361. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Molecular Formula | Molecular Weight (g/mol) | Structures | Contents (mg/g) |
|---|---|---|---|---|
| (+)-Catechin | C15H14O6 | 290.08 |  | 5.287 |
| Taxifolin | C15H12O7 | 304.06 |  | 8.419 |
| Eriodictyol | C15H12O6 | 288.06 |  | 6.583 |
| Morin | C15H10O7 | 302.04 |  | 4.651 |
| Myricetin | C15H10O8 | 318.04 |  | 4.042 |
| Luteolin | C15H10O6 | 286.05 |  | 3.421 |
| Naringenin | C15H12O5 | 272.07 |  | 0.623 |
| Apigenin | C15H10O5 | 270.05 |  | 0.203 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Huang, R.; Han, S.; Li, X.; Gong, H.; Zhang, Q.; Yan, C.; Li, Y.; He, R. Potential of Tamarind Shell Extract against Oxidative Stress In Vivo and In Vitro. Molecules 2023, 28, 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041885
Li W, Huang R, Han S, Li X, Gong H, Zhang Q, Yan C, Li Y, He R. Potential of Tamarind Shell Extract against Oxidative Stress In Vivo and In Vitro. Molecules. 2023; 28(4):1885. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041885
Chicago/Turabian StyleLi, Weixi, Rongping Huang, Shaocong Han, Xiyou Li, Haibiao Gong, Qiongyi Zhang, Changyu Yan, Yifang Li, and Rongrong He. 2023. "Potential of Tamarind Shell Extract against Oxidative Stress In Vivo and In Vitro" Molecules 28, no. 4: 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041885