
Critical Role of Monooxygenase in Biodegradation of 2,4,6-Trinitrotoluene by Buttiauxella sp. S19-1
 , and
, and 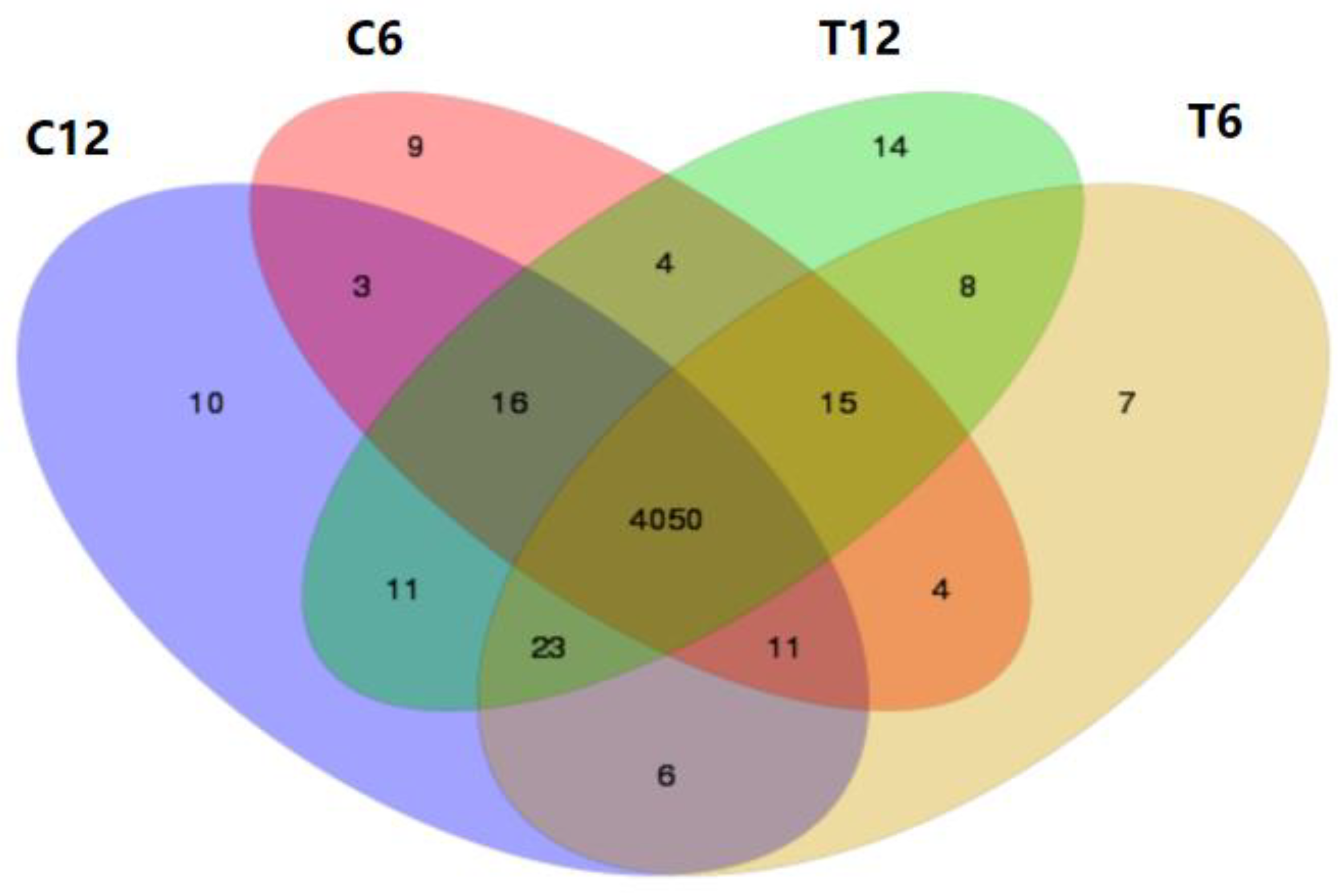{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Transcriptomic Analysis of Strain S19-1
2.1.1. Sequencing Data Filtering
2.1.2. Screening of Differentially Expressed Genes (DEGs) during TNT Degradation
2.1.3. The Gene Ontology (GO) Term Enrichment Analysis of DEGs
2.1.4. Kyoto Encyclopaedia of Genes and Genomes (KEGG) Enrichment Analysis of Differential Genes
2.1.5. Spatiotemporal Sequence Analysis
2.1.6. Transcriptomic Analysis of BuMO
2.2. Bioinformatics Analysis of BuMO
2.3. Biodegradation of TNT by Mutants
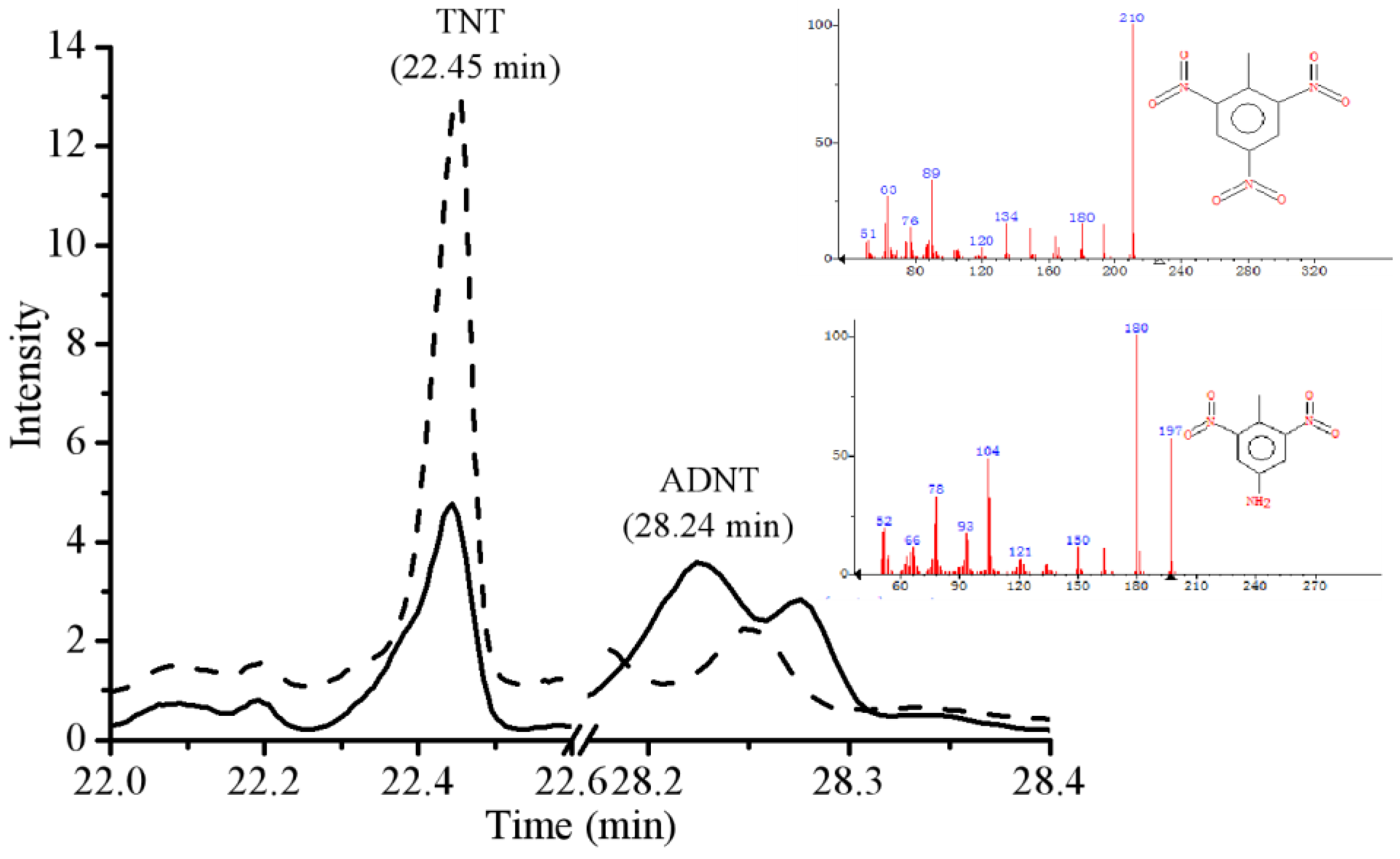
2.4. Gas Chromatography Mass Spectrometry (GC-MS) Analysis
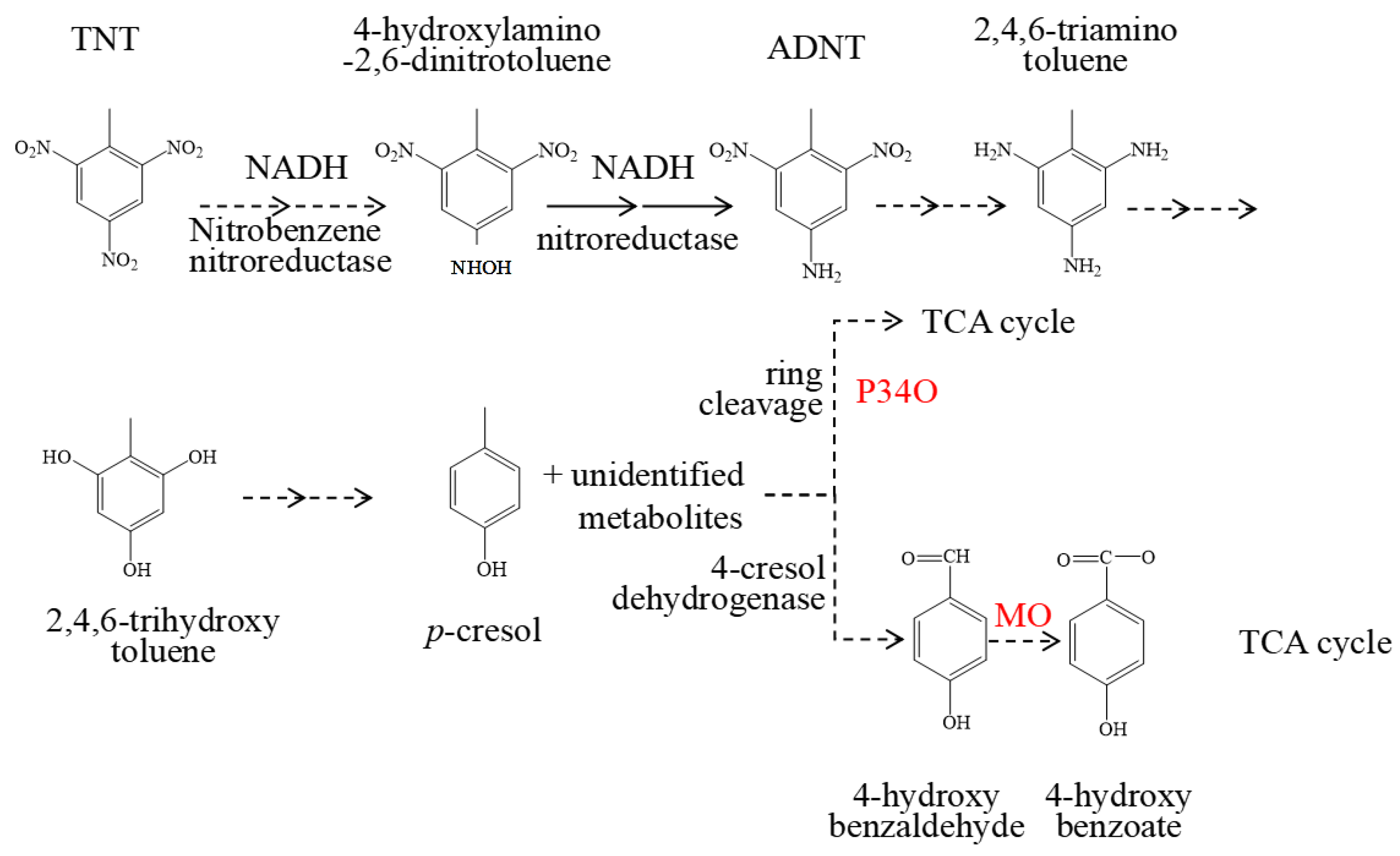
2.5. The Key Enzymes Involved in Biodegradation of TNT in Strain S19-1
3. Materials and Methods
3.1. Culture of Bacteria and Cometabolic Degradation of TNT
3.2. Determination of TNT Concentrations in Bacterial Cultures
3.3. Transcriptomic Analysis of Strain S19-1 under TNT Exposure
3.3.1. Sample Processing
3.3.2. Library Construction, Sequencing, and Bioinformatics Analysis
3.4. Cloning Vectors and Associated Reagents
3.5. Isolation and Amplification of Monooxygenase (BuMO) Wild-Type Gene
3.6. Construction of BuMO Expression Vectors
3.7. Subcloning of BuMO Full Sequence and ΔBuMO Sequence
3.8. Generating the BuMO Knockout Strain (S-ΔMO)
3.9. Construction of E-MO (a BuMO-Expressing E. coli Mutant)
3.10. Construction of BuMO Expression Vector
3.11. Bioinformatics Analysis of BuMO
3.12. Purification and Analysis of Recombinant BuMO (rBuMO)
3.13. Incubation of Strains in TNT-Containing Medium
3.14. GC-MS Analysis of TNT Biodegradation Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Wilbrand, J. Notiz über Trinitrotoluol. Justus. Liebigs. Ann. Chem. 1863, 128, 178–179. [Google Scholar] [CrossRef] [Green Version]
- Travis, A.S. The Synthetic Nitrogen Industry in World War I: Its Emergence and Expansion; Springer: Berlin/Heidelberg, Germany, 2015; Volume 163. [Google Scholar]
- Beck, A.J.; Gledhill, M.; Schlosser, C.; Stamer, B.; Bottcher, C.; Sternheim, J.; Greinert, J.; Achterberg, E.P. Spread, behavior, and ecosystem consequences of conventional munitions compounds in coastal marine waters. Front. Mar. Sci. 2018, 5, 141. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, C.; Knobloch, T.; Rühl, N.P.; Sternheim, J.; Wichert, U.; Wöhler, J. Munitionsbelastung der deutschen meeresgewässer-bestandsaufnahme und empfehlungen. Available online: https://www.munition-im-meer.de (accessed on 10 November 2011).
- Pfeiffer, F. Changes in properties of explosives due to prolonged seawater exposure. Mar. Technol. Soc. J. 2012, 46, 102–110. [Google Scholar] [CrossRef]
- Beck, A.J.; van der Lee, E.M.; Eggert, A.; Stamer, B.; Gledhill, M.; Schlosser, C.; Achterberg, E.P. In situ measurements of explosive compound dissolution fluxes from exposed munition material in the Baltic Sea. Environ. Sci. Technol. 2019, 53, 5652–5660. [Google Scholar] [CrossRef]
- Bünning, T.H.; Strehse, J.S.; Hollmann, A.C.; Bötticher, T.; Maser, E. A toolbox for the determination of nitroaromatic explosives in marine water, sediment, and biota samples on femtogram levels by GC-MS/MS. Toxics 2021, 9, 60. [Google Scholar] [CrossRef]
- Lotufo, G.R.; Rosen, G.; Wild, W.; Carton, G. Summary Review of the Aquatic Toxicology of Munitions Constituents. 2013. Available online: https://www.researchgate.net/publication/267337013 (accessed on 1 June 2013).
- Rieger, P.G.; Sinnwell, V.; Preuss, A.; Franke, W.; Knackmuss, H.J. Hydride-Meisenheimer complex formation and protonation as key reactions of 2,4,6-trinitrophenol biodegradation by Rhodococcus erythropolis. J. Bacteriol. 1999, 181, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Strehse, J.S.; Appel, D.; Geist, C.; Martin, H.J.; Maser, E. Biomonitoring of 2,4,6-trinitrotoluene and degradation products in the marine environment with transplanted blue mussels (M. edulis). Toxicology 2017, 390, 117–123. [Google Scholar] [CrossRef]
- Appel, D.; Strehse, J.S.; Martin, H.J.; Maser, E. Bioaccumulation of 2,4,6-trinitrotoluene (TNT) and its metabolites leaking from corroded munition in transplanted blue mussels (M. edulis). Mar. Pollut. Bull. 2018, 135, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Maser, E.; Strehse, J.S. “Don’t Blast”: Blast-in-place (BiP) operations of dumped World War munitions in the oceans significantly increase hazards to the environment and the human seafood consumer. Arch. Toxicol. 2020, 94, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Schuster, R.; Strehse, J.S.; Ahvo, A.; Turja, R.; Maser, E.; Bickmeyer, U.; Lehtonen, K.K.; Brenner, M. Exposure to dissolved TNT causes multilevel biological effects in Baltic mussels (Mytilus spp.). Mar. Environ. Res. 2021, 167, 105264. [Google Scholar] [CrossRef]
- Bolt, H.M.; Degen, G.H.; Dorn, S.B.; Plöttner, S.; Harth, V. Genotoxicity and potential carcinogenicity of 2,4,6-TNT trinitrotoluene: Structural and toxicological considerations. Rev. Environ. Health 2006, 21, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Koske, D.; Goldenstein, N.I.; Kammann, U. Nitroaromatic compounds damage the DNA of zebrafish embryos (Danio rerio). Aquat. Toxicol. 2019, 217, 105345. [Google Scholar] [CrossRef] [PubMed]
- Marinovic, V.; Ristic, M.; Dostanic, M. Dynamic adsorption of trinitrotoluene on granular activated carbon. J. Hazard Mater. 2005, 117, 121–128. [Google Scholar] [CrossRef]
- An, F.Q.; Gao, B.J.; Feng, X.Q. Adsorption performance and mechanism of 2,4,6-trinitrotoluene on a novel adsorption material polyvinylbenzyl acid/SiO2. Appl. Surf. Sci. 2009, 255, 5031–5035. [Google Scholar] [CrossRef]
- Chen, W.S.; Chiang, W.C.; Lai, C.C. Recovery of nitrotoluenes in wastewater by solvent extraction. J. Hazard Mater. 2007, 145, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Duijm, N.J.; Markert, F. Assessment of technologies for disposing explosive waste. J. Hazard Mater. 2002, 90, 137–153. [Google Scholar] [CrossRef]
- Lee, B.J.; Jeong, S.W. Effects of additives on 2,4,6-trinitrotoluene (TNT) removal and its mineralization in aqueous solution by gamma irradiation. J. Hazard Mater. 2009, 165, 435–440. [Google Scholar] [CrossRef]
- Matta, R.; Hanna, K.; Chiron, S. Fenton-like oxidation of 2,4,6-trinitrotoluene using different iron minerals. Sci. Total Environ. 2007, 385, 242–251. [Google Scholar] [CrossRef]
- Huang, J.; Tang, W.Y.; Zhou, S.F. Preliminary study on the treatment of TNT redwater by wet peroxide oxidation under normal pressure. Chin. J. Explos. Propellants 1998, 21, 48–50. [Google Scholar]
- Gomez-Gardenes, J.; Lotero, L.; Taraskin, S.N.; Perez-Reche, F.J. Explosive contagion in networks. Sci. Rep. 2016, 6, 19767. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Gonzalez, M.Y.; Chandra, R.; Castillo-Zacarias, C.; Robledo-Padilla, F.; Rostro-Alanis, M.D.J.; Parra-Saldivar, R. Biotransformation and degradation of 2,4,6-trinitrotoluene by microbial metabolism and their interaction. Def. Technol. 2018, 14, 151–164. [Google Scholar] [CrossRef]
- Maksimova, Y.G.; Maksimov, A.Y.; Demakov, V.A. Biotechnological approaches to the bioremediation of an environment polluted with trinitrotoluene. Appl. Biochem. Microbiol. 2018, 54, 767–779. [Google Scholar] [CrossRef]
- Arora, P.K.; Kumar, M.; Chauhan, A.; Raghava, G.P.; Jain, R.K. OxDBase: A database of oxygenases involved in biodegradation. Bio. Med. Cent. Res. Notes. 2009, 2, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, H.S. Comparative biochemistry of the phenolase complex. Adv. Enzymol. Relat. Subj. Biochem. 1955, 16, 105–184. [Google Scholar] [PubMed]
- Choi, H.J.; Seo, J.Y.; Hwang, S.M.; Lee, Y.-I.; Jeong, Y.K.; Moon, J.Y.; Joo, W.H. Isolation and characterization of BTEX tolerant and degrading Pseudomonas putida BCNU 106. Biotechnol. Bioprocess Eng. 2013, 18, 1000–1007. [Google Scholar] [CrossRef]
- Fessner, N.D.; Grimm, C.; Kroutil, W.; Glieder, A. Late-stage functionalisation of polycyclic (N-Hetero-) aromatic hydrocarbons by detoxifying CYP5035S7 monooxygenase of the white-rot fungus Polyporus arcularius. Biomolecules 2021, 11, 1708. [Google Scholar] [CrossRef]
- Guo, Y.; Li, D.F.; Ji, H.N.; Zheng, J.T.; Zhou, N.Y. Hexachlorobenzene monooxygenase substrate selectivity and catalysis: Structural and biochemical insights. Appl. Environ. Microbiol. 2020, 87, e01965-20. [Google Scholar] [CrossRef]
- Smets, B.F.; Yin, H.; Esteve-Nunez, A. TNT biotransformation: When chemistry confronts mineralization. Appl. Microbiol. Biotechnol. 2007, 76, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Keenan, B.G.; Wood, T.K. Orthric Rieske dioxygenases for degrading mixtures of 2,4-dinitrotoluene/naphthalene and 2-amino-4,6-dinitrotoluene/4-amino-2,6-dinitrotoluene. Appl. Microbiol. Biotechnol. 2006, 73, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Liu, D.; Sun, P.; Li, Y.N.; Wu, M.; Liu, W.C.; Maser, E.; Xiong, G.M.; Guo, L.Q. Degradation of 2,4,6-trinitrotoluene (TNT): Involvement of protocatechuate 3,4-dioxygenase (P34O) in Buttiauxella sp. S19-1. Toxics 2021, 9, 231. [Google Scholar] [CrossRef]
- Hsu, D.W.; Wang, T.I.; Huang, D.J.; Pao, Y.J.; Lin, Y.A.; Cheng, T.W.; Liang, S.H.; Chen, C.Y.; Kao, C.M.; Sheu, Y.T.; et al. Copper promotes E. coli laccase-mediated TNT biotransformation and alters the toxicity of TNT metabolites toward Tigriopus japonicus. Ecotox. Environ. Saf. 2019, 173, 452–460. [Google Scholar] [CrossRef] [PubMed]
- McClay, K.; Fox, B.G.; Steffan, R.J. Toluene monooxygenase catalyzed epoxidation of alkenes. Appl. Environ. Microbiol. 2000, 66, 1877–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, H.V.; Hodgson, J.R.; Hwang, S.W.; Halpap, L.M.; Kelton, D.O.; Andersen, B.S.; VanGoethem, D.L.; Lee, C.C. Mammalian toxicity of munitions compounds. Phase I. acute oral toxicity, primary skin and eye irritation, dermal sensitization, disposition and metabolism and ames tests of additional compounds. Prog. Rep. 1978, 6, 1–37. [Google Scholar]
- Esteve-Núnez, A.; Caballero, A.; Ramos, J.L. Biological degradation of 2,4,6-trinitrotoluene. Microbiol. Mol. Biol. Rev. 2001, 65, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenhuis, R.; Vink, R.L.; Janssen, D.B.; Witholt, B. Degradation of chlorinated aliphatic hydrocarbons by Methylosinus trichosporium OB3b expressing soluble methane monooxygenase. Appl. Environ. Microbiol. 1989, 55, 2819–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wackett, L.P.; Brusseau, G.A.; Householder, S.R.; Hanson, R.S. Survey of microbial oxygenases: Trichloroethylene degradation by propane-oxidizing bacteria. Appl. Environ. Microbiol. 1989, 55, 2960–2964. [Google Scholar] [CrossRef] [Green Version]
- Hamamura, N.; Page, C.; Long, T.; Semprini, L.; Arp, D.J. Chloroform cometabolism by butane-grown CF8, Pseudomonas butanovora, and Mycobacterium vaccae JOB5 and methane grown Methylosinus trichosporium OB3b. Appl. Environ. Microbiol. 1997, 63, 3607–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, N.V.; Mattes, T.E.; Gossett, J.M.; Spain, J.C. Phylogenetic and kinetic diversity of aerobic vinyl chloride assimilating bacteria from contaminated sites. Appl. Environ. Microbiol. 2002, 68, 6162–6171. [Google Scholar] [CrossRef] [Green Version]
- Stenuit, B.A.; Agathos, S.N. Biodegradation and bioremediation of TNT and other nitro explosives. Compr. Biotechnol. 2011, 6, 167–181. [Google Scholar] [CrossRef]
- Oh, B.T.; Shea, P.J.; Drijber, R.A.; Vasilyeva, G.K.; Sarath, G. TNT biotransformation and detoxification by a Pseudomonas aeruginosa strain. Biodegradation 2003, 14, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1983; Volume 33, pp. 721–722. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; He, L.; Sun, P.; Wu, M.; Cui, X.; Liu, D.; Adomako-Bonsu, A.G.; Geng, M.; Xiong, G.; Guo, L.; et al. Critical Role of Monooxygenase in Biodegradation of 2,4,6-Trinitrotoluene by Buttiauxella sp. S19-1. Molecules 2023, 28, 1969. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041969
Xu M, He L, Sun P, Wu M, Cui X, Liu D, Adomako-Bonsu AG, Geng M, Xiong G, Guo L, et al. Critical Role of Monooxygenase in Biodegradation of 2,4,6-Trinitrotoluene by Buttiauxella sp. S19-1. Molecules. 2023; 28(4):1969. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041969
Chicago/Turabian StyleXu, Miao, Lei He, Ping Sun, Ming Wu, Xiyan Cui, Dong Liu, Amma G. Adomako-Bonsu, Min Geng, Guangming Xiong, Liquan Guo, and et al. 2023. "Critical Role of Monooxygenase in Biodegradation of 2,4,6-Trinitrotoluene by Buttiauxella sp. S19-1" Molecules 28, no. 4: 1969. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules28041969