Impairment of the Developing Human Brain in Iron Deficiency: Correlations to Findings in Experimental Animals and Prospects for Early Intervention Therapy


Abstract
:1. Introduction
1.1. Transport of Iron into the Brain
1.2. Transport of Iron into the Developing and Iron-Deficient Brain
1.3. The Significance of Iron for Precursor Cells of the Developing Brain
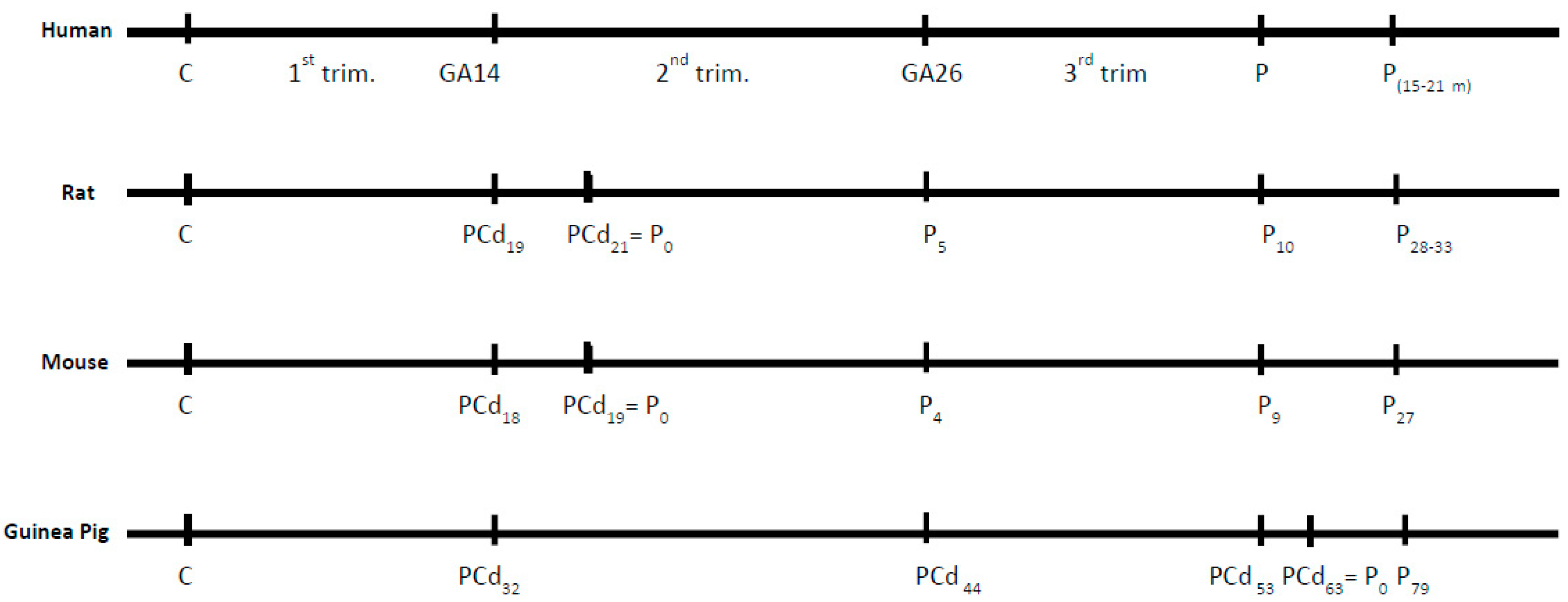
1.4. Translational Models of the Brain Development
1.5. Evidence of Deleterious Effects of Iron Deficiency on the Developing Brain
1.6. Experimental Animals
1.7. The Developing Human Brain
2. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BBB | blood–brain barrier |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
References
- Zimmermann, M.B.; Hurrell, R.F. Nutritional iron deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef]
- Breymann, C.; Bian, X.M.; Blanco-Capito, L.R.; Chong, C.; Mahmud, G.; Rehman, R. Expert recommendations for the diagnosis and treatment of iron-deficiency anemia during pregnancy and the postpartum period in the Asia-Pacific region. J. Perinat. Med. 2011, 39, 113–121. [Google Scholar] [CrossRef]
- Hernandez-Martinez, C.; Canals, J.; Aranda, N.; Ribot, B.; Escribano, J.; Arija, V. Effects of iron deficiency on neonatal behavior at different stages of pregnancy. Early Hum. Dev. 2011, 87, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Khalafallah, A.; Dennis, A.; Bates, J.; Bates, G.; Robertson, I.K.; Smith, L.; Ball, M.J.; Seaton, D.; Brain, T.; Rasko, J.E. A prospective randomized, controlled trial of intravenous versus oral iron for moderate iron deficiency anaemia of pregnancy. J. Int. Med. 2010, 268, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Milman, N. Serum ferritin in Danes: Studies of iron status from infancy to old age, during blood donation and pregnancy. Int. J. Hematol. 1996, 63, 103–135. [Google Scholar] [CrossRef]
- Camaschella, C. Iron deficiency. Blood 2019, 133, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashiri, A.; Burstein, E.; Sheiner, E.; Mazor, M. Anemia during pregnancy and treatment with intravenous iron: Review of the literature. Eur. J. Obs. Gyn. Reprod. Biol. 2003, 110, 2–7. [Google Scholar] [CrossRef]
- Shao, J.; Lou, J.; Rao, R.; Georgieff, M.K.; Kaciroti, N.; Felt, B.T.; Zhao, Z.Y.; Lozoff, B. Maternal serum ferritin concentration is positively associated with newborn iron stores in women with low ferritin status in late pregnancy. J. Nutr. 2012, 142, 2004–2049. [Google Scholar] [CrossRef] [PubMed]
- Duck, K.A.; Connor, J.R. Iron uptake and transport across physiological barriers. Biometals 2016, 29, 573–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallman, P.R.; Spirito, R.A. Brain iron in the rat: Extremely slow turnover in normal rats may explain long-lasting effects of early iron deficiency. J. Nutr. 1977, 107, 1075–1081. [Google Scholar] [CrossRef]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J. Nutr. 2001, 131, 568S–579S. [Google Scholar] [CrossRef] [PubMed]
- Beard, J. Iron deficiency alters brain development and functioning. J. Nutr. 2003, 133 (Suppl. 1), 1468s–1472s. [Google Scholar] [CrossRef] [PubMed]
- Moos, T.; Morgan, E.H. A morphological study of the developmentally regulated transport of iron into the brain. Dev. Neurosci. 2002, 24, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, A.M.; Rao, R.; Long, J.D.; Widness, J.A.; Georgieff, M.K. The assessment of newborn iron stores at birth: A review of the literature and standards for ferritin concentrations. Neonatology 2007, 92, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, C.; Henderson, D.J.; Matthews, D.J.; Copp, A.J. Transferrin and its receptor in the development of genetically determined neural tube defects in the mouse embryo. Dev. Dyn. 1996, 207, 35–46. [Google Scholar] [CrossRef]
- Matak, P.; Matak, A.; Moustafa, S.; Aryal, D.K.; Benner, E.J.; Wetsel, W.; Andrews, N. Disrupted iron homeostasis causes dopaminergic neurodegeneration in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 3428–3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferies, W.A.; Brandon, M.R.; Hunt, S.V.; Williams, A.F.; Gatter, K.C.; Mason, D.Y. Transferrin receptor on endothelium of brain capillaries. Nature 1984, 312, 162–163. [Google Scholar] [CrossRef]
- Meyron-Holtz, E.G.; Cohen, L.A.; Fahoum, L.; Haimovich, Y.; Lifshitz, L.; Magid-Gold, I.; Stuemler, T.; Truman-Rosentsvit, M. Ferritin polarization and iron transport across monolayer epithelial barriers in mammals. Front. Pharmacol. 2014, 5, 194. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.; Jia, X.; Zhou, M.; Wang, K.; Conde, J.; He, J.; Tian, J.; Yan, X. Ferritin nanocarrier traverses the blood brain barrier and kills glioma. ACS Nano 2018, 12, 4105–4115. [Google Scholar] [CrossRef]
- Fiandra, L.; Mazzucchelli, S.; Truffi, M.; Bellini, M.; Sorrentino, L.; Corsi, F. In vitro permeation of FITC-loaded ferritins across a rat blood-brain barrier: A model to study the delivery of nanoformulated molecules. J. Vis. Exp. 2016, 114. [Google Scholar] [CrossRef]
- Chiou, B.; Neal, E.H.; Bowman, A.B.; Lippmann, E.S.; Simpson, I.A.; Connor, J.R. Endothelial cells are critical regulators of iron transport in a model of the human blood-brain barrier. J. Cereb. Blood Flow Metab. 2018. [Google Scholar] [CrossRef]
- Skjørringe, T.; Burkhart, A.; Johnsen, K.B.; Moos, T. Divalent metal transporter 1 (DMT1) in the brain: Implications for a role in iron transport at the blood-brain barrier, and neuronal and glial pathology. Front. Mol. Neurosci. 2015, 8, 19. [Google Scholar] [PubMed]
- Burkhart, A.; Skjørringe, T.; Johnsen, K.B.; Siupka, P.; Thomsen, L.B.; Nielsen, M.S.; Thomsen, L.L.; Moos, T. Expression of iron-related proteins at the neurovascular unit supports reduction and reoxidation of iron for transport through the blood-brain barrier. Mol. Neurobiol. 2016, 53, 7237–7253. [Google Scholar] [CrossRef] [PubMed]
- Skjørringe, T.; Møller, L.B.; Moos, T. Impairment of interrelated iron- and copper homeostatic mechanisms in brain contributes to the pathogenesis of neurodegenerative disorders. Front. Pharmacol. 2012, 3, 169. [Google Scholar] [CrossRef] [PubMed]
- Theil, E.T. Ferritin: Structure, gene regulation, and cellular function in animals, plants, and microorganisms. Ann. Rev. Biochem. 1987, 56, 289–315. [Google Scholar] [CrossRef] [PubMed]
- d’Estaintot, B.L.; Santambrogio, P.; Granier, T.; Gallois, B.; Chevalier, J.M.; Precigoux, G.; Levi, S.; Arosio, P. Crystal structure and biochemical properties of the human mitochondrial ferritin and its mutant Ser144Ala. J. Mol. Biol. 2004, 340, 277–293. [Google Scholar]
- Møllgård, K.; Dziegielewska, K.M.; Saunders, N.R.; Zakut, H.; Soreq, H. Synthesis and localization of plasma proteins in the developing human brain. Integrity of the fetal blood-brain barrier to endogenous proteins of hepatic origin. Dev. Biol. 1988, 128, 207–221. [Google Scholar] [CrossRef]
- Moos, T.; Oates, P.S.; Morgan, E.H. Expression of the neuronal transferrin receptor is age dependent and susceptible to iron deficiency. J. Comp. Neurol. 1998, 398, 420–430. [Google Scholar] [CrossRef]
- Knutson, M.D. Non-transferrin-bound iron transporters. Free Radic. Biol. Med. 2019, 133, 101–111. [Google Scholar] [CrossRef]
- Taylor, E.M.; Morgan, E.H. Developmental changes in transferrin and iron uptake by the brain in the rat. Dev. Brain Res. 1990, 55, 35–42. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Restricted transport of anti-transferrin receptor antibody [OX26] through the blood-brain barrier in the rat. J. Neurochem. 2001, 79, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Connor, J.R.; Beard, J.L. Brain iron, transferrin and ferritin concentrations are altered in developing iron-deficient rats. J. Nutr. 1995, 125, 1529–1535. [Google Scholar] [PubMed]
- Laskey, J.; Webb, I.; Schulman, H.M.; Ponka, P. Evidence that transferrin supports cell proliferation by supplying iron for DNA synthesis. Exp. Cell Res. 1988, 176, 87–95. [Google Scholar] [CrossRef]
- Taylor, E.M.; Crowe, A.; Morgan, E.H. Transferrin and iron uptake by the brain: Effects of altered iron status. J. Neurochem. 1991, 57, 1584–1592. [Google Scholar] [CrossRef]
- Richard, C.; John, M.; Wrigglesworth, H.B. Iron-dependent enzymes in mammalian systems. In Transport and Storage; Chapter: Iron-Dependent Enzymes in Mammalian Systems; Ponka, P., Schulman, H.M., Woodworth, R.C., Richter, G.W., Eds.; CRC Press: Stockholm, Sweden, 1990; pp. 17–39. [Google Scholar]
- Lozoff, B.; Georgieff, M.K. Iron deficiency and brain development. Semin. Pediatr. Neurol. 2006, 13, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Nutrition and the developing brain: Nutrient priorities and measurement. Am. J. Clin. Nutr. 2007, 85, 614–620. [Google Scholar]
- Wright, J.A.; Chan, A.K.; Choy, B.K.; Hurta, R.A.; McClarty, G.A.; Tagger, A.Y. Regulation and drug resistance mechanisms of mammalian ribonucleotide reductase, and the significance to DNA synthesis. Biochem. Cell Biol. 1990, 68, 1364–1371. [Google Scholar]
- Moos, T.; Skjorringe, T.; Thomsen, L.L. Iron deficiency and iron treatment in the fetal developing brain—A pilot study introducing an experimental rat model. Reprod. Health 2018, 15 (Suppl. 1), 93. [Google Scholar] [CrossRef]
- Oloyede, O.B.; Folayan, A.T.; Odutuga, A.A. Effects of low-iron status and deficiency of essential fatty acids on some biochemical constituents of rat brain. Biochem. Int. 1992, 27, 913–922. [Google Scholar]
- Möller, H.E.; Bossoni, L.; Connor, J.R.; Crichton, R.R.; Does, M.D.; Ward, R.J.; Zecca, L.; Zucca, F.A.; Ronen, I. Iron, myelin, and the brain: Neuroimaging meets neurobiology. Trends Neurosci. 2019, 42, 384–401. [Google Scholar] [CrossRef]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling transformations of neurodevelopmental sequences across mammalian species. J. Neurosci. 2013, 33, 7368–7383. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgenson, L.A.; Sun, M.; O’Connor, M.; Georgieff, M.K. Fetal iron deficiency disrupts the maturation of synaptic function and efficacy in area CA1 of the developing rat hippocampus. Hippocampus 2005, 15, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Eseh, R.; Zimmerberg, B. Age-dependent effects of gestational and lactational iron deficiency on anxiety behavior in rats. Behav. Brain Res. 2005, 164, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Felt, B.T.; Beard, J.L.; Schallert, T.; Shao, J.; Aldridge, J.W.; Connor, J.R.; Georgieff, M.K.; Lozoff, B. Persistent neurochemical and behavioral abnormalities in adulthood despite early iron supplementation for perinatal iron deficiency anemia in rats. Behav. Brain Res. 2006, 171, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unger, E.L.; Paul, T.; Murray-Kolb, L.E.; Felt, B.; Jones, B.C.; Beard, J.L. Early iron deficiency alters sensorimotor development and brain monoamines in rats. J. Nutr. 2007, 137, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.V.; Carlson, E.S.; Fretham, S.J.; Georgieff, M.K. Early-life iron deficiency anemia alters neurotrophic factor expression and hippocampal neuron differentiation in male rats. J. Nutr. 2008, 138, 2495–2501. [Google Scholar] [CrossRef]
- Bourque, S.L.; Iqbal, U.; Reynolds, J.N.; Adams, M.A.; Nakatsu, K. Perinatal iron deficiency affects locomotor behavior and water maze performance in adult male and female rats. J. Nutr. 2008, 138, 931–937. [Google Scholar] [CrossRef]
- Wu, L.L.; Zhang, L.; Shao, J.; Qin, Y.F.; Yang, R.W.; Zhao, Z.Y. Effect of perinatal iron deficiency on myelination and associated behaviors in rat pups. Behav. Brain Res. 2008, 188, 263–270. [Google Scholar] [CrossRef]
- McEchron, M.D.; Cheng, A.Y.; Liu, H.; Connor, J.R.; Gilmartin, M.R. Perinatal nutritional iron deficiency permanently impairs hippocampus-dependent trace fear conditioning in rats. Nutr. Neurosci. 2005, 8, 195–206. [Google Scholar] [CrossRef]
- Schmidt, A.T.; Ladwig, E.K.; Wobken, J.D.; Grove, W.M.; Georgieff, M.K. Delayed alternation performance in rats following recovery from early iron deficiency. Physiol. Behav. 2010, 101, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Brunette, K.E.; Tranm, P.V.; Wobken, J.D.; Carlson, E.S.; Georgieff, M.K. Gestational and neonatal iron deficiency alters apical dendrite structure of CA1 pyramidal neurons in adult rat hippocampus. Dev. Neurosci. 2010, 32, 238–248. [Google Scholar] [CrossRef]
- Bastian, T.W.; Prohaska, J.R.; Georgieff, M.K.; Anderson, G.W. Perinatal iron and copper deficiencies alter neonatal rat circulating and brain thyroid hormone concentrations. Endocrinology 2010, 151, 4055–4065. [Google Scholar] [CrossRef]
- Mihaila, C.; Schramm, J.; Strathmann, F.G.; Lee, D.L.; Gelein, R.M.; Luebke, A.E.; Mayer-Pröschel, M. Identifying a window of vulnerability during fetal development in a maternal iron restriction model. PLoS ONE 2011, 6, e17483. [Google Scholar] [CrossRef]
- Rao, R.; Tkac, I.; Schmidt, A.T.; Georgieff, M.K. Fetal and neonatal iron deficiency causes volume loss and alters the neurochemical profile of the adult rat hippocampus. Nutr. Neurosci. 2011, 14, 59–65. [Google Scholar] [CrossRef]
- Callahan, L.S.; Thibert, K.A.; Wobken, J.D.; Georgieff, M.K. Early-life iron deficiency anemia alters the development and long-term expression of parvalbumin and perineuronal nets in the rat hippocampus. Dev. Neurosci. 2013, 35, 427–436. [Google Scholar] [CrossRef]
- Greminger, A.R.; Lee, D.L.; Shrager, P.; Mayer-Proschel, M. Gestational iron deficiency differentially alters the structure and function of white and gray matter brain regions of developing rats. J. Nutr. 2014, 144, 1058–1066. [Google Scholar] [CrossRef]
- Greminger, A.R.; Mayer-Proschel, M. Identifying the threshold of iron deficiency in the central nervous system of the rat by the auditory brainstem response. ASN Neuro 2015, 7. [Google Scholar] [CrossRef]
- Bastian, T.W.; Santarriaga, S.; Nguyen, T.A.; Prohaska, J.R.; Georgieff, M.K.; Anderson, G.W. Fetal and neonatal iron deficiency but not copper deficiency increases vascular complexity in the developing rat brain. Nutr. Neurosci. 2015, 18, 365–375. [Google Scholar] [CrossRef]
- Hu, X.; Wang, R.; Shan, Z.; Dong, Y.; Zheng, H.; Jesse, F.F.; Rao, E.; Takahashi, E.; Li, W.; Teng, W.; et al. Perinatal iron deficiency-induced hypothyroxinemia impairs early brain development regardless of normal iron levels in the neonatal brain. Thyroid 2016, 26, 891–900. [Google Scholar] [CrossRef]
- Bastian, T.W.; von Hohenberg, W.C.; Mickelson, D.J.; Lanier, L.M.; Georgieff, M.K. Iron deficiency impairs developing hippocampal neuron gene expression, energy metabolism, and dendrite complexity. Dev. Neurosci. 2016, 38, 264–276. [Google Scholar] [CrossRef]
- Horiquini-Barbosa, E.; Gibb, R.; Kolb, B.; Bray, D.; Lachat, J.J. Tactile stimulation partially prevents neurodevelopmental changes in visual tract caused by early iron deficiency. Brain Res. 2017, 1657, 130–139. [Google Scholar] [CrossRef]
- Vieyra-Reyes, P.; Millan-Aldaco, D.; Palomero-Rivero, M.; Jimenez-Garces, C.; Hernandez-Gonzalez, M.; Caballero-Villarraso, J. An iron-deficient diet during development induces oxidative stress in relation to age and gender in Wistar rats. J. Physiol. Biochem. 2017, 73, 99–110. [Google Scholar] [CrossRef]
- Rosato-Siri, M.V.; Marziali, L.; Guitart, M.E.; Badaracco, M.E.; Puntel, M.; Pitossi, F.; Correale, J.; Pasquini, J.M. Iron availability compromises not only oligodendrocytes but also astrocytes and microglial cells. Mol. Neurobiol. 2018, 55, 1068–1081. [Google Scholar] [CrossRef]
- Hubbard, A.C.; Bandyopadhyay, S.; Wojczyk, B.S.; Spitalnik, S.L.; Hod, E.A.; Prestia, K.A. Effect of dietary iron on fetal growth in pregnant mice. Comp. Med. 2013, 63, 127–135. [Google Scholar]
- Unger, E.L.; Earley, C.J.; Thomsen, L.L.; Jones, B.C.; Allen, R.P. Effects of IV iron isomaltoside-1000 treatment on regional brain iron status in an iron-deficient animal. Neuroscience 2013, 246, 179–185. [Google Scholar] [CrossRef]
- Jougleux, J.L.; Rioux, F.M.; Church, M.W.; Fiset, S.; Jacques, H.; Surette, M.E. Dietary LC-PUFA in iron-deficient anaemic pregnant and lactating guinea pigs induce minor defects in the offsprings’ auditory brainstem responses. Nutr. Neurosci. 2016, 19, 447–460. [Google Scholar] [CrossRef]
- Jougleux, J.L.; Rioux, F.M.; Church, M.W.; Fiset, S.; Surette, M.E. Mild maternal iron deficiency anemia during pregnancy and lactation in guinea pigs causes abnormal auditory function in the offspring. J. Nutr. 2011, 141, 1390–1395. [Google Scholar] [CrossRef]
- Jougleux, J.L.; Rioux, F.M.; Church, M.W.; Fiset, S.; Surette, M.E. Mild iron deficiency anaemia during pregnancy and lactation in guinea pigs alters amplitudes and auditory nerve velocity, but not brainstem transmission times in the offspring’s auditory brainstem response. Nutr. Neurosci. 2014, 17, 37–47. [Google Scholar] [CrossRef]
- Antonides, A.; van Laarhoven, S.; van der Staay, F.J.; Nordquist, R.E. Non-anemic iron deficiency from birth to weaning does not impair growth or memory in piglets. Front. Behav. Neurosci. 2016, 10, 112. [Google Scholar] [CrossRef]
- Mudd, A.T.; Fil, J.E.; Knight, L.C.; Dilger, R.N. Dietary iron repletion following early-life dietary iron deficiency does not correct regional volumetric or diffusion tensor changes in the developing pig brain. Front. Neurol. 2017, 8, 735. [Google Scholar] [CrossRef]
- Schachtschneider, K.M.; Liu, Y.; Rund, L.A.; Madsen, O.; Johnson, R.W.; Groenen, M.A.; Schook, L.B. Impact of neonatal iron deficiency on hippocampal DNA methylation and gene transcription in a porcine biomedical model of cognitive development. BMC Genom. 2016, 17, 856. [Google Scholar] [CrossRef]
- Leyshon, B.J.; Radlowski, E.C.; Mudd, A.T.; Steelman, A.J.; Johnson, R.W. Postnatal iron deficiency alters brain development in piglets. J. Nutr. 2016, 146, 1420–1427. [Google Scholar] [CrossRef]
- Rao, R.; Ennis, K.; Oz, G.; Lubach, G.R.; Georgieff, M.K.; Coe, C.L. Metabolomic analysis of cerebrospinal fluid indicates iron deficiency compromises cerebral energy metabolism in the infant monkey. Neurochem. Res. 2013, 38, 573–580. [Google Scholar] [CrossRef]
- Rao, R.; Ennis, K.; Lubach, G.R.; Lock, E.F.; Georgieff, M.K.; Coe, C.L. Metabolomic analysis of CSF indicates brain metabolic impairment precedes hematological indices of anemia in the iron-deficient infant monkey. Nutr. Neurosci. 2018, 21, 40–48. [Google Scholar] [CrossRef]
- Patton, S.M.; Coe, C.L.; Lubach, G.R.; Connor, J.R. Quantitative proteomic analyses of cerebrospinal fluid using iTRAQ in a primate model of iron deficiency anemia. Dev. Neurosci. 2012, 34, 354–365. [Google Scholar] [CrossRef]
- Lubach, G.R.; Coe, C.L. Selective impairment of cognitive performance in the young monkey following recovery from iron deficiency. J. Dev. Behav. Pediatr. JDBP 2008, 29, 11–17. [Google Scholar] [CrossRef]
- Golub, M.S.; Hogrefe, C.E.; Germann, S.L.; Capitanio, J.P.; Lozoff, B. Behavioral consequences of developmental iron deficiency in infant rhesus monkeys. Neurotox. Teratol. 2006, 28, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Guitart, M.E.; Vence, M.; Correale, J.; Pasquini, J.M.; Rosato-Siri, M.V. Ontogenetic oligodendrocyte maturation through gestational iron deprivation: The road not taken. Glia 2019, 67, 1760–1774. [Google Scholar] [CrossRef]
- Szudzik, M.; Starzyński, R.R.; Jończy, A.; Mazgaj, R.; Lenartowicz, M.; Lipiński, P. Iron supplementation in suckling piglets: An ostensibly easy therapy of neonatal iron deficiency anemia. Pharmaceuticals 2018, 11, 128. [Google Scholar] [CrossRef]
- Lozoff, B. Iron deficiency and child development. Food Nutr. Bull. 2007, 28 (Suppl. 4), S560–S671. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, D.; Anupurba, S.; Verma, A.; Kumar, A. Effect of maternal iron deficiency anemia on fetal neural development. J. Perinatol. 2018, 38, 233–239. [Google Scholar] [CrossRef]
- Berglund, S.K.; Torres-Espinola, F.J.; Garcia-Valdes, L.; Segura, M.; Martinez-Zaldivar, C.; Padilla, C.; Rueda, R.; Pérez García, M.; McArdle, H.J.; Campoy, C. The impacts of maternal iron deficiency and being overweight during pregnancy on neurodevelopment of the offspring. Br. J. Nutr. 2017, 118, 533–540. [Google Scholar] [CrossRef]
- Carpenter, K.L.H.; Li, W.; Wei, H.; Wu, B.; Xiao, X.; Liu, C.; Worley, G.; Egger, H.L. Magnetic susceptibility of brain iron is associated with childhood spatial IQ. Neuro Image 2016, 132, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Algarin, C.; Karunakaran, K.D.; Reyes, S.; Morales, C.; Lozoff, B.; Peirano, P.; Biswal, B. Differences on brain connectivity in adulthood are present in subjects with iron deficiency anemia in infancy. Front. Aging Neurosci. 2017, 9, 54. [Google Scholar] [CrossRef]
- Insel, B.J.; Schaefer, C.A.; McKeague, I.W.; Susser, E.S.; Brown, A.S. Maternal iron deficiency and the risk of schizophrenia in offspring. Arch. Gen. Psychiatry 2008, 65, 1136–1144. [Google Scholar] [CrossRef]
- Chen, M.H.; Su, T.P.; Chen, Y.S.; Hsu, J.W.; Huang, K.L.; Chang, W.H.; Chen, T.J.; Bai, Y.M. Association between psychiatric disorders and iron deficiency anemia among children and adolescents: A nationwide population-based study. BMC Psychiatry 2013, 13, 161. [Google Scholar] [CrossRef]
- Lane, R.; Kessler, R.; Buckley, A.W.; Rodriguez, A.; Farmer, C.; Thurm, A.; Swedo, S.; Felt, B. Evaluation of periodic limb movements in sleep and iron status in children with autism. Pediatr. Neurol. 2015, 53, 343–349. [Google Scholar] [CrossRef]
- Berglund, S.K.; Westrup, B.; Haraldsson, E.; Engstrom, B.; Domellof, M. Effects of iron supplementation on auditory brainstem response in marginally LBW infants. Pediatr. Res. 2011, 70, 601–606. [Google Scholar] [CrossRef]
- ElAlfy, M.S.; El-Farrash, R.A.; Taha, H.M.; Ismail, E.A.; Mokhtar, N.A. Auditory brainstem response in full-term neonates born to mothers with iron deficiency anemia: Relation to disease severity. J. Matern.-Fetal Neonatal Med. 2018, 4, 1–8. [Google Scholar] [CrossRef]
- Sundagumaran, H.; Seethapathy, J. Auditory brainstem response in infants with iron deficiency anemia. Int. J. Pediatr. Otorhinolaryngol. 2019, 117, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, V.; Amin, S.B.; Agarwal, A.; Srivastava, L.M.; Soni, A.; Saluja, S. Latent iron deficiency at birth influences auditory neural maturation in late preterm and term infants. Am. J. Clin. Nutr. 2015, 102, 1030–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monga, M.; Walia, V.; Gandhi, A.; Chandra, J.; Sharma, S. Effect of iron deficiency anemia on visual evoked potential of growing children. Brain Dev. 2010, 32, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Armony-Sivan, R.; Kaciroti, N.; Jing, Y.; Golub, M.; Jacobson, S.W. Eye-blinking rates are slower in infants with iron-deficiency anemia than in nonanemic iron-deficient or iron-sufficient infants. J. Nutr. 2010, 140, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Amony-Sivan, R.; Zhu, B.; Clark, K.M.; Richards, B.; Ji, C.; Kaciroti, N.; Shao, J.; Lozoff, B. Iron deficiency (ID) at both birth and 9 months predicts right frontal EEG asymmetry in infancy. Dev. Psychobiol. 2016, 58, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Otero, G.A.; Pliego-Rivero, F.B.; Porcayo-Mercado, R.; Mendieta-Alcantara, G. Working memory impairment and recovery in iron deficient children. Clin. Neurophysiol. 2008, 119, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, A.F.; Koss, M.; Burden, M.J.; Jonides, J.; Nelson, C.A.; Kaciroti, N.; Jimenez, E.; Lozoff, B. Iron deficiency in infancy and neurocognitive functioning at 19 years: Evidence of long-term deficits in executive function and recognition memory. Nutr. Neurosci. 2010, 13, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Congdon, E.L.; Westerlund, A.; Algarin, C.R.; Peirano, P.D.; Gregas, M.; Lozoff, B.; Nelson, C.A. Iron deficiency in infancy is associated with altered neural correlates of recognition memory at 10 years. J. Pediatr. 2012, 160, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Clark, K.M.; Jing, Y.; Armony-Sivan, R.; Angelilli, M.L.; Jacobson, S.W. Dose-response relationships between iron deficiency with or without anemia and infant social-emotional behavior. J. Pediatr. 2008, 152, 696–702. [Google Scholar] [CrossRef]
- Parsons, A.G.; Zhou, S.J.; Spurrier, N.J.; Makrides, M. Effect of iron supplementation during pregnancy on the behaviour of children at early school age: Long-term follow-up of a randomised controlled trial. Br. J. Nutr. 2008, 99, 1133–1139. [Google Scholar] [CrossRef]
- Qubty, W.; Renaud, D.L. Cognitive impairment associated with low ferritin responsive to iron supplementation. Pediatr. Neurol. 2014, 51, 831–833. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Cui, N.; Liu, J. Neurocognitive function is associated with serum iron status in early adolescents. Biol. Res. Nurs. 2017, 19, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Mireku, M.O.; Davidson, L.L.; Boivin, M.J.; Zoumenou, R.; Massougbodji, A.; Cot, M.; Bodeau-Livinec, F. Prenatal iron deficiency, neonatal ferritin, and infant cognitive function. Pediatrics 2016, 138. [Google Scholar] [CrossRef] [PubMed]
- Doom, J.R.; Georgieff, M.K.; Gunnar, M.R. Institutional care and iron deficiency increase ADHD symptomology and lower IQ 2.5–5 years post-adoption. Dev. Sci. 2015, 18, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.; Seth, S.; Saha, S.; Roy, A.; Das, R.; Datta, A.K. A study on association of iron deficiency with attention deficit hyperactivity disorder in a tertiary care center. Ind. J. Psychiatry 2018, 60, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Holm, C.; Thomsen, L.L.; Norgaard, A.; Langhoff-Roos, J. Single-dose intravenous iron infusion or oral iron for treatment of fatigue after postpartum haemorrhage: A randomized controlled trial. Vox Sang. 2017, 112, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Markova, V.; Norgaard, A.; Jørgensen, K.J.; Langhoff-Roos, J. Treatment for women with postpartum iron deficiency anaemia. Cochrane Database Syst. Rev. 2015, 13, CD010861. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Study Design | Method | Offspring Age | Conclusion | Reference |
|---|---|---|---|---|---|
| Rat | ID before conception + Gestational ID | Electrophysiological recordings | P15–P30, P65 | Late term effects on synapses in hippocampus in spite of cerebral iron repletion | [44] |
| ID before conception | Behavior | P10–Adult | Some persistent effects in spite of iron repletion | [45] | |
| ID before conception + Gestational ID | Brain iron Neurotransmitters | P35 | Behavioral impairments related to persistent loss in dopamine in spite of brain iron reversal | [46] | |
| Gestational ID + Lactational ID | mRNA | P6–P21 | ID from E15 leads to alteration in tyrosine hydroxylase and reversibility in behavior | [47] | |
| Gestational ID + Lactational ID | mRNA, proteins morphology | P7–P15; P30 | Lower BDNF, impaired neuronal differentiation | [48] | |
| ID before conception + Gestational ID | Behavior | Adult | Detrimental effects of behavioral tasks, sex dependency | [49] | |
| Gestational ID | Myelination | P25 | Impaired myelination with correlation to impairment | [50] | |
| Gestational ID | mRNA, proteins | P32–P69 | Effect of behavior, no effects on motor skills in hippocampus | [51] | |
| Gestational ID + Lactational ID | Behavior | P65 | Permanent changes in behavioral tasks | [52] | |
| Gestational ID + Lactational ID | mRNA morphology | P7–P65 | Permanent changes in mRNA of neuronal markers and dendritic branching in spite of postnatal reversal to normal diet | [53] | |
| Gestational ID | mRNA, T3, T4 | P12 | Marked reduction in T3, T4 | [54] | |
| Rat | ID before conception + Gestational ID | ABR, DPOAE | P0–P45 | First trimester displays profound changes in auditory brain stem response | [55] |
| Gestational ID + Lactational ID | MRI, NMR | P7–56 | Restoration of brain iron, permanent size reduction in hippocampus and neurochemical hall-markers in spite of postnatal reversal to normal diet | [56] | |
| Gestational ID | mRNA | P7–P56 | Impaired formation of neuronal network and impaired neuronal plasticity in spite of postnatal reversal to normal diet | [57] | |
| Gestational ID | Morphology | P21–P40 | 25% reduction in dendritic length 20% reduction in axonal diameter | [58] | |
| ID before conception + Gestational ID | ABR | P40 | Increased ABR latencies in ID depending on stage of ID | [59] | |
| Gestational ID + Lactational ID | mRNA | P10–P15 | Elevated angiogenic/vasculogenic signaling with increased blood vessel complexity | [60] | |
| ID before conception + gestational ID | mRNA, T3, T4 | E13–P10 | Marked reduction in T3, T4 Lowering of thyroid hormone responsive genes | [61] | |
| Embryonic brain | mRNA | Not available (Cultures at E16) | DFO-induced ID lowers expression of series of markers of dendritic and synaptic development, and mitochondrial function | [62] | |
| Gestational ID | Tactile stimuli | P1–P32 | Tactile stimuli reverse defect myelination and alteration in oligodendrocytes and microglia, but not astrocytes | [63] | |
| Rat | Gestational ID | Pro/anti-oxidant | P0–P70 | Age- and iron-dependent levels of oxidative stress profiling | [64] |
| Gestational ID | mRNA, IHC | P21, P35 | Defect myelination, alteration in glial cells | [65] | |
| Mouse | Gestational ID Brain iron | Hematology | E17–E18 | Effect of brain weight, lower brain iron | [66] |
| ID in offspring | Brain iron | Adult | Correction of cerebral ID with parenteral iron | [67] | |
| Guinea Pig | Gestational ID + Lactational ID | ABR | P9–P24 | Effect of ABR in ID Part restoration with PUFA treatment | [68] |
| Gestational ID + Lactational ID | ABR | P24 | Effect of ABR in ID | [69,70] | |
| Domestic Pig | Gestational ID + Lactational ID | Cognitive tasks | 0–4 weeks after birth | No cognitive deficits | [71] |
| ID in offspring | MRI | 0–6 weeks after birth | Cerebral ID, alteration in brain tissue composition persists in spite of iron repletion | [72] | |
| Lactational ID | RNA analysis | 4 weeks after birth | Change in hippocampal DNA methylation and gene regulation | [73] | |
| Gestational ID | MRI, IHC | PD 2–30 | ID after PD 14 detriments white matter | [74] | |
| Monkey | ID in offspring | 1H NMR | Infancy | Change in metabolomic profile in CSF | [75] |
| ID in offspring | Proteomic | Infancy | Persistent change in proteomic profile in CSF | [76] | |
| ID in offspring | 1H NMR | Infancy | Metabolomic profile in CSF predicts effects of ID on brain iron metabolism | [77] | |
| ID in offspring | Cognitive tasks | Infancy | Only initial cognitive + behavioral deficits | [78] | |
| Gestational ID + Lactational ID | Cognitive tasks | Infancy | Cognitive and emotional effects present, but vary with protocol | [79] |
| Study Objective | Evidence of ID | Infant Age | No. F/O | Conclusion | Reference |
|---|---|---|---|---|---|
| Fetal brain development | |||||
| Normal development | Maternal IDA | PND 3–5 days | /70 | Maternal IDA adversely affects l hippocampal morphogenesis and fetal production of BDNF | [83] |
| Normal development | Maternal IDA | 18 months | 331/ | Maternal ID at 34 weeks associated with lower motor scores | [84] |
| Normal development | Normal iron status | 7–11 years | /39 | MRI iron content in basal ganglia influences spatial intelligence | [85] |
| Brain connectivity | Infant IDA | Mean 21.5 years | /31 | Different patterns of functional connectivity between former IDA and control young adults | [86] |
| Risk of schizophrenia | Maternal IDA | Prospective study | /6872 | Maternal ID as risk factor for schizophrenia in offspring | [87] |
| Cerebral functions | IDA in adults | Adult | /2957 | IDA associated with increases in psychiatric disorders | [88] |
| Autism | Infant IDA | 2–7 years | /102 | No evidence between IDA and autism | [89] |
| Basic cerebral functions | |||||
| ABR | LBW | PND 42–6 months | /285 | Iron supplements did not improve ABR, but ABR was discarded as measure of impairment in ID | [90] |
| ABR | Maternal IDA | PND 2, 3 months | ABR closely related to severity of maternal and neonatal iron status | [91] | |
| ABR | Infant IDA | 6–24 months | Prolonged latencies in ABR traces in IDA | [92] | |
| ABR | Infant IDA | <48 h | /90 | Latent iron deficiency associated with abnormal ABR | [93] |
| VEP | Infant IDA | 6–24 months | /50 | Negative correlation between severity of IDA and VEP latencies | [94] |
| Eye-blinking rates | Infant IDA | 9–10 months | 61 | Increased eye-blink rats consistent with low dopamine function in IDA | [95] |
| EEG recordings | Infant IDA | 0, 9 months | /80 | ID associated with EEG asymmetry | [96] |
| Memory | |||||
| Memory | Infant IDA | 8–10 years | /201 | Iron supplementation substantially restores cognitive capabilities | [97] |
| Execution, memory | Infant IDA | 19 years | /114 | Chronic impairment of functions related to frontostriatal-connections (executive functions), and hippocampus (recognition memory) | [98] |
| Recognition memory | Infant IDA | 6–18 months | /209 | Sustained effects on memory in 10-year follow-up in spite of oral supplement in early life | [99] |
| Higher cerebral functions | |||||
| Social-emotional behavior | Infant IDA | 9–10 years | /77 | Social-emotional behavior associated with ID | [100] |
| Behavior | Normal | 6–8 years | /264 | Fe supplementation in pregnancy without consistent effect on behavior | [101] |
| Cognition | Infant IDA | 1–3 years | /3 | Improvement in cognition once iron stores were restored | [102] |
| Cognition | Infant IDA | Mean age 12.0 | Reduced cognitive performance | [103] | |
| Cognition | Infant IDA | 12 months | 828/828 | No effect of IDA on cognition or motor development | [104] |
| ADHD symptomology, IQ | Infant IDA | 2.5–5 years | /123 | Effects of early deprivation and ID on ADHD symptoms and IQ years after adoption | [105] |
| ADHD symptomology | Infant IDA | Mean age 11.0 | IDA associated with ADHD | [106] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markova, V.; Holm, C.; Pinborg, A.B.; Thomsen, L.L.; Moos, T. Impairment of the Developing Human Brain in Iron Deficiency: Correlations to Findings in Experimental Animals and Prospects for Early Intervention Therapy. Pharmaceuticals 2019, 12, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030120
Markova V, Holm C, Pinborg AB, Thomsen LL, Moos T. Impairment of the Developing Human Brain in Iron Deficiency: Correlations to Findings in Experimental Animals and Prospects for Early Intervention Therapy. Pharmaceuticals. 2019; 12(3):120. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030120
Chicago/Turabian StyleMarkova, Veronika, Charlotte Holm, Anja Bisgaard Pinborg, Lars Lykke Thomsen, and Torben Moos. 2019. "Impairment of the Developing Human Brain in Iron Deficiency: Correlations to Findings in Experimental Animals and Prospects for Early Intervention Therapy" Pharmaceuticals 12, no. 3: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/ph12030120