
Naphthyridine Derivatives Induce Programmed Cell Death in Naegleria fowleri

 , , , and
, , , and
Abstract
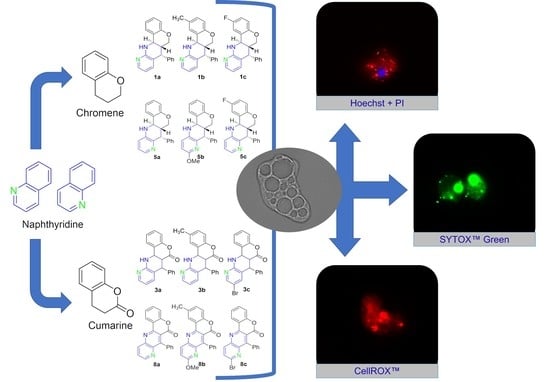
:
1. Introduction
2. Results
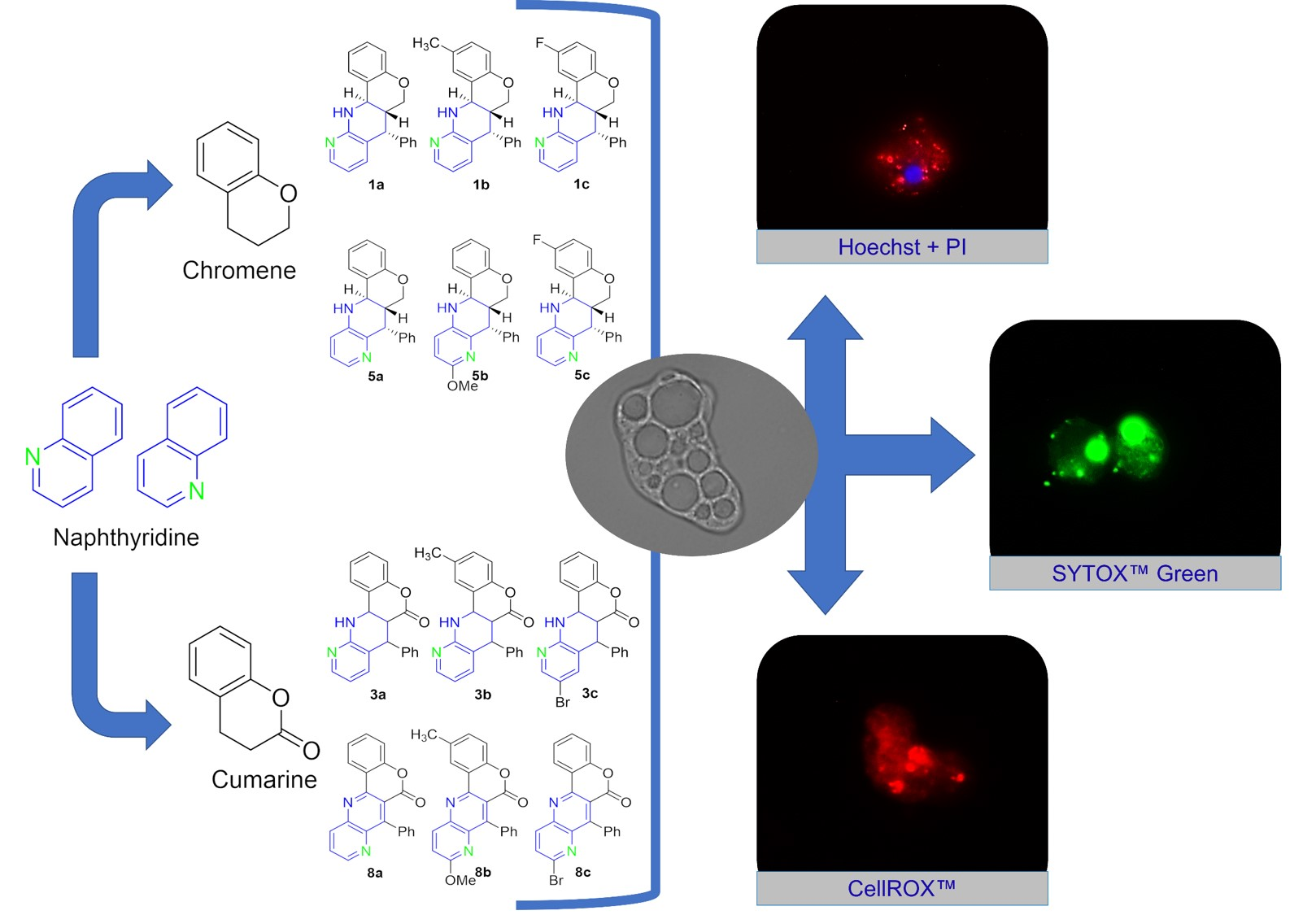
2.1. In Vitro Evaluation of Amoebicidal Activity and Cytotoxicity of Fused 1,5 Naphthyridine Derivatives
2.2. Evaluation of PCD Induction by the Selected Compounds
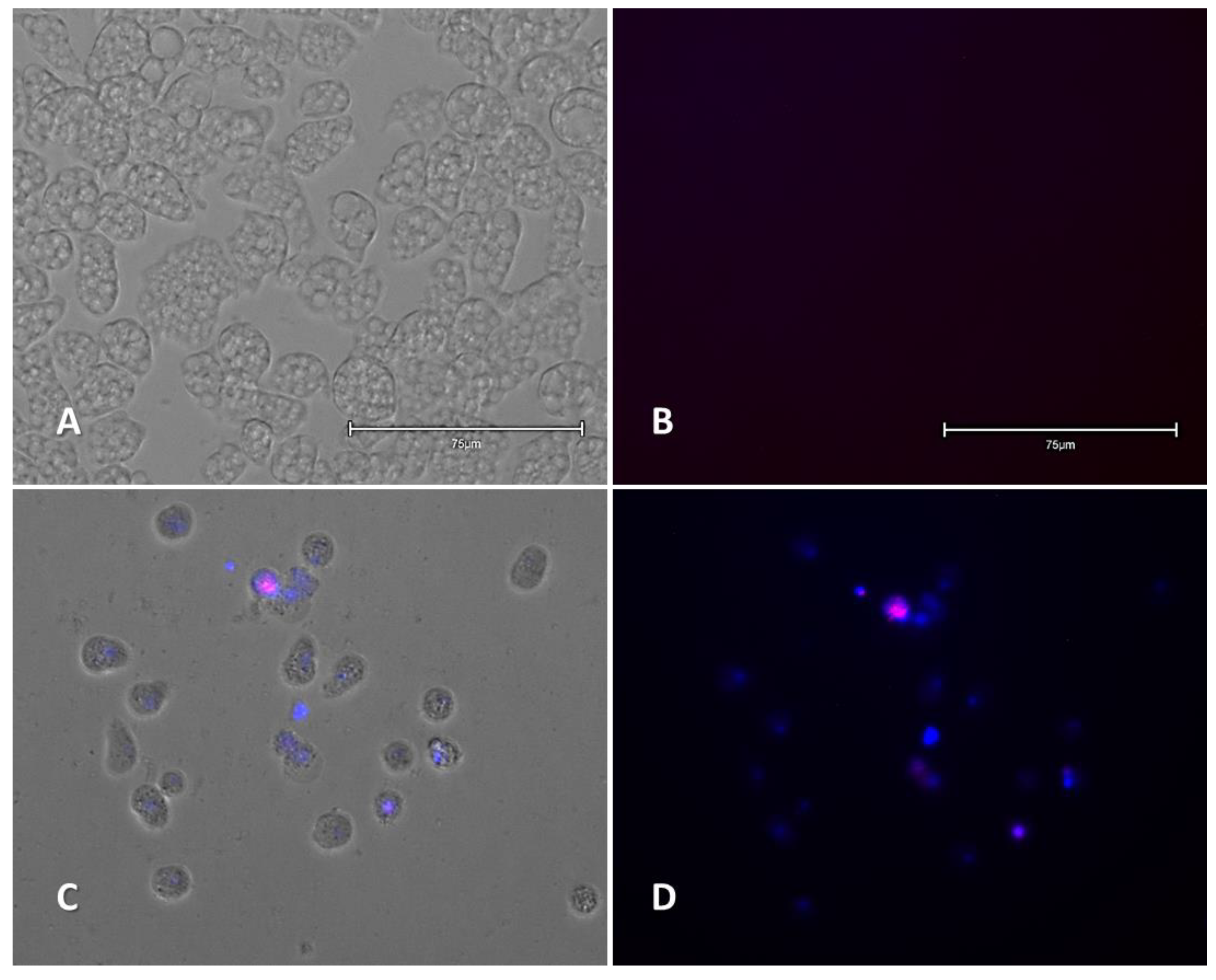
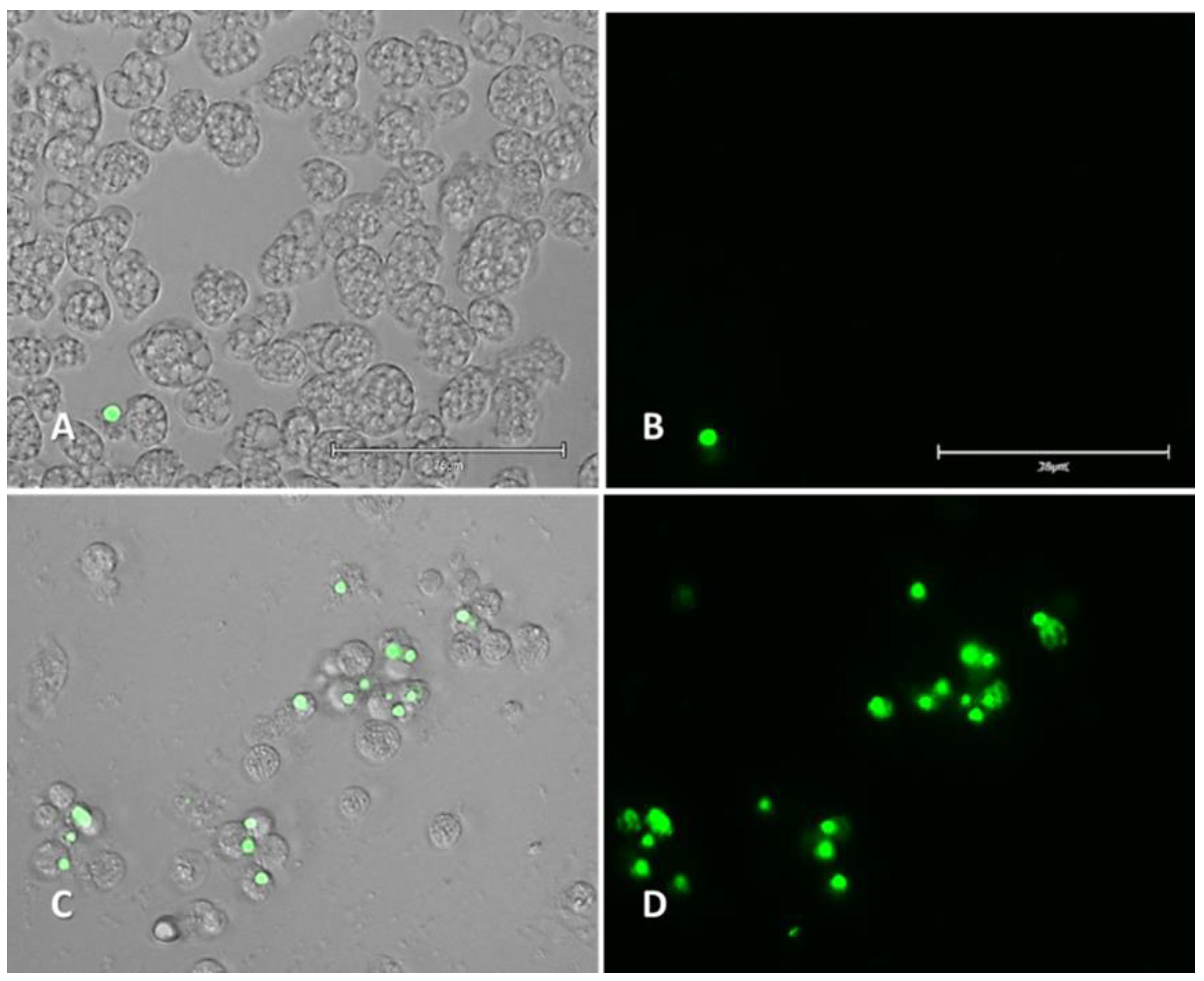
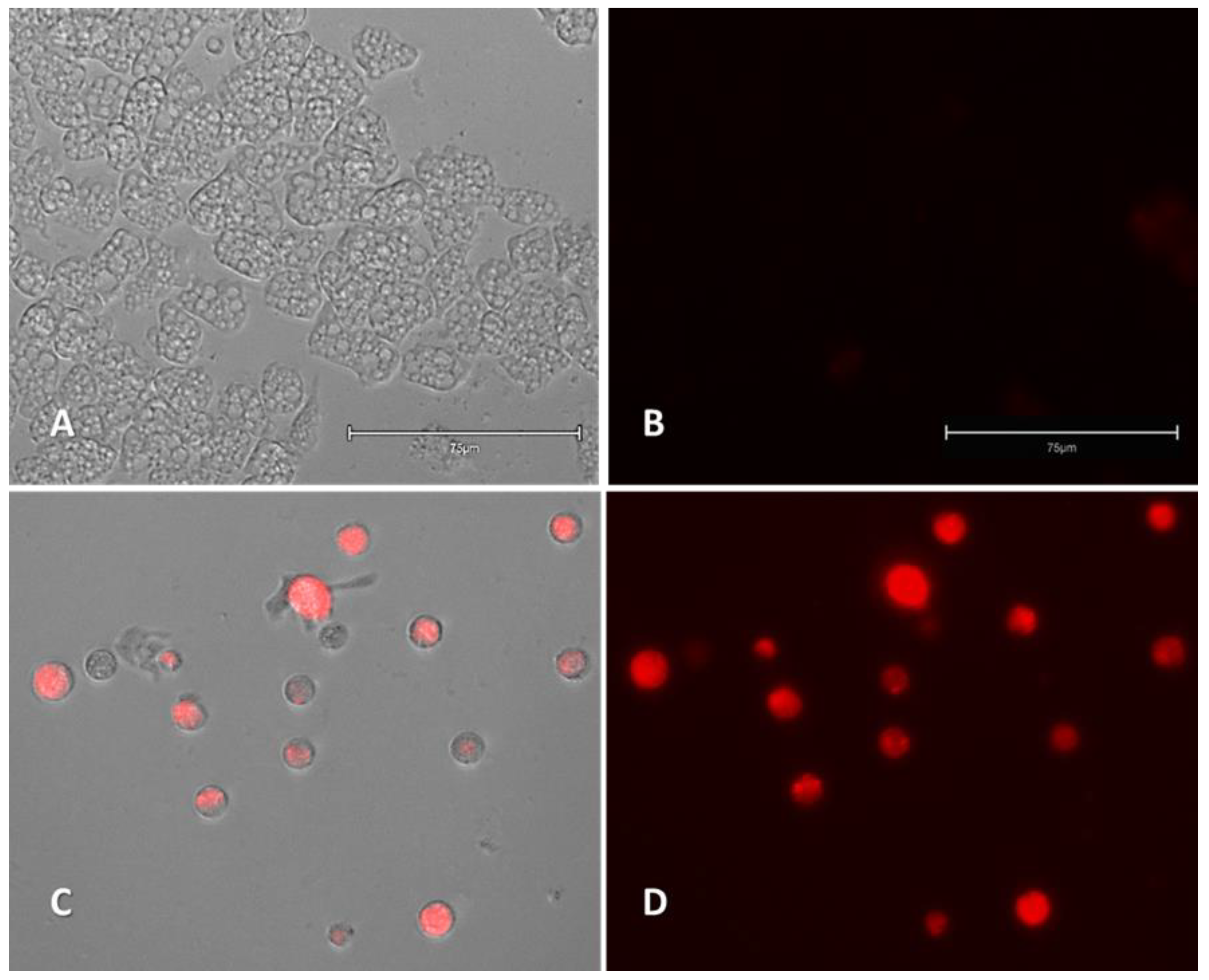
2.2.1. Double-Stain Assay for the Detection of Chromatin Condensation (Hoechst 33342/PI)
2.2.2. Plasma Membrane Permeability (SYTOX Green)
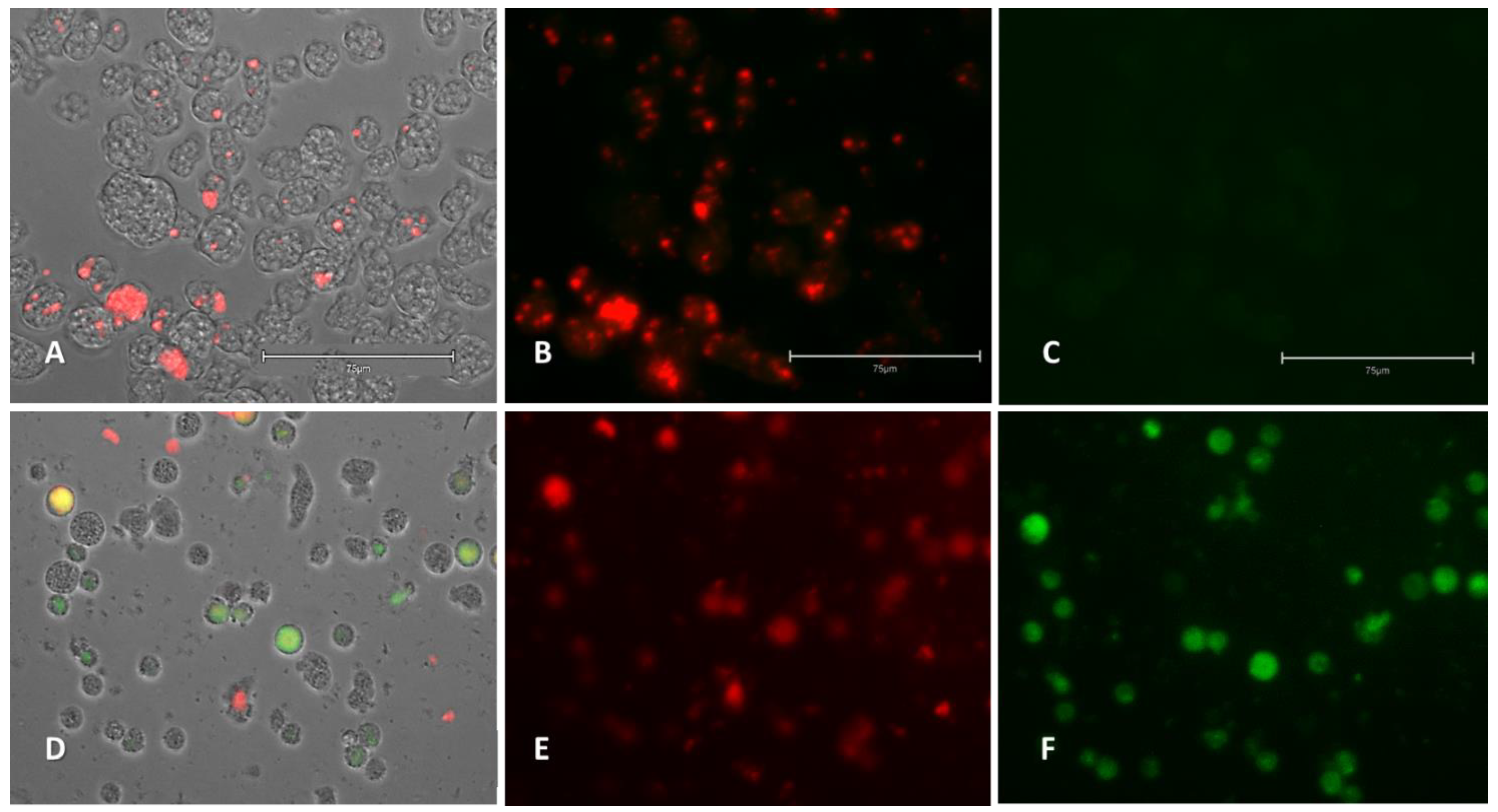
2.2.3. Generation of Intracellular Reactive Oxygen Species (ROS)
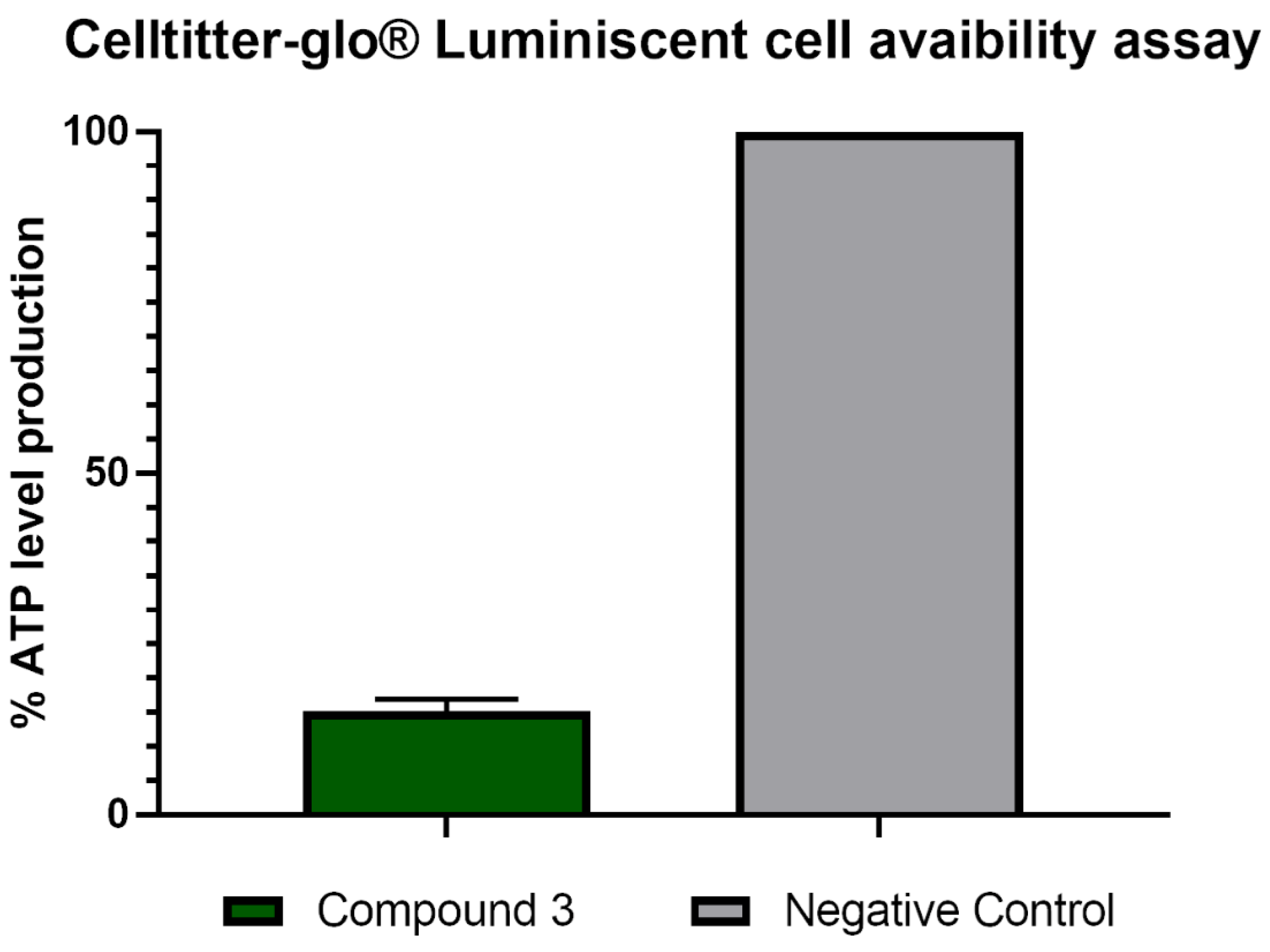
2.2.4. Analysis of Mitochondrial Function Disruption
3. Discussion
4. Materials and Methods
4.1. Amoebic Strains and Cell Line Maintenance
4.2. Chemistry General Experimental Information
4.3. Compound Purity Analysis
4.4. In Vitro Amoebicidal Activity
4.5. In Vitro Cytotoxicity
4.6. Analysis of Programmed Cell Death (PCD) Induction
4.6.1. Detection of Chromatin Condensation
4.6.2. Plasma Membrane Alteration
4.6.3. Reactive Oxygen Species (ROS) Generation
4.6.4. Analysis of Mitochondrial Function Disruption
Mitochondrial Membrane Potential
Evaluation of ATP Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tillery, L.; Barrett, K.; Goldstein, J.; Lassner, J.W.; Osterhout, B.; Tran, N.L.; Xu, L.; Young, R.M.; Craig, J.; Chun, I.; et al. Naegleria fowleri: Protein structures to facilitate drug discovery for the deadly, pathogenic free-living amoeba. PLoS ONE 2021, 16, e0241738. [Google Scholar] [CrossRef]
- Jamerson, M.; da Rocha-Azevedo, B.; Cabral, G.A.; Marciano-Cabral, F. Pathogenic Naegleria fowleri and non-pathogenic Naegleria lovaniensis exhibit differential adhesion to, and invasion of, extracellular matrix proteins. Microbiology 2012, 158, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Güémez, A.; García, E. Primary Amoebic Meningoencephalitis by Naegleria fowleri: Pathogenesis and Treatments. Biomolecules 2021, 11, 1320. [Google Scholar] [CrossRef]
- Fowler, M.; Carter, R.F. Acute Pyogenic Meningitis Probably Due to Acanthamoeba sp.: A Preliminary Report. Br. Med. J. 1965, 2, 734–742. [Google Scholar] [CrossRef] [Green Version]
- Maciver, S.K.; Piñero, J.E.; Lorenzo-Morales, J. Is Naegleria fowleri an Emerging Parasite? Trends Parasitol. 2020, 36, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Gharpure, R.; Bliton, J.; Goodman, A.; Ali, I.K.M.; Yoder, J.; Cope, J.R. Epidemiology and Clinical Characteristics of Primary Amebic Meningoencephalitis Caused by Naegleria fowleri: A Global Review. Clin. Infect. Dis. 2021, 73, e19–e27. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Jamal, S.B.; Farhat, S.M. Naegleria fowleri in Pakistan. Lancet Infect. Dis. 2020, 20, 27–28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, M.; Hu, B.; Chen, H.; Pan, J.R.; Ruan, W.; Yao, L. Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in China. Int. J. Infect. Dis. 2018, 72, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matanock, A.; Mehal, J.M.; Liu, L.; Blau, D.M.; Cope, J.R. Estimation of Undiagnosed Naegleria fowleri Primary Amebic Meningoencephalitis, United States. Emerg. Infect. Dis. 2018, 24, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Arberas-Jiménez, I.; García-Davis, S.; Rizo-Liendo, A.; Sifaoui, I.; Reyes-Batlle, M.; Chiboub, O.; Rodríguez-Expósito, R.L.; Díaz-Marrero, A.R.; Piñero, J.E.; Fernández, J.J.; et al. Laurinterol from Laurencia johnstonii eliminates Naegleria fowleri triggering PCD by inhibition of ATPases. Sci. Rep. 2020, 10, 17731. [Google Scholar] [CrossRef] [PubMed]
- Rizo-Liendo, A.; Arberas-Jiménez, I.; Sifaoui, I.; Reyes-Batlle, M.; Piñero, J.E.; Lorenzo-Morales, J. The type 2 statins, cerivastatin, rosuvastatin and pitavastatin eliminate Naegleria fowleri at low concentrations and by induction of programmed cell death (PCD). Bioorg. Chem. 2021, 110, 104784. [Google Scholar] [CrossRef] [PubMed]
- Rizo-Liendo, A.; Sifaoui, I.; Cartuche, L.; Arberas-Jiménez, I.; Reyes-Batlle, M.; Fernández, J.J.; Piñero, J.E.; Díaz-Marrero, A.R.; Lorenzo-Morales, J. Evaluation of indolocarbazoles from streptomyces sanyensis as a novel source of therapeutic agents against the brain-eating amoeba Naegleria fowleri. Microorganisms 2020, 8, 789. [Google Scholar] [CrossRef]
- Jahangeer, M.; Mahmood, Z.; Munir, N.; Waraich, U.A.; Tahir, I.M.; Akram, M.; Ali Shah, S.M.; Zulfqar, A.; Zainab, R. Naegleria fowleri: Sources of infection, pathophysiology, diagnosis, and management; a review. Clin. Exp. Pharmacol. Physiol. 2020, 47, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Cope, J.R.; Ali, I.K. Primary Amebic Meningoencephalitis: What Have We Learned in the Last 5 Years? Curr. Infect. Dis. Rep. 2016, 18, 31. [Google Scholar] [CrossRef]
- Schuster, F.L.; Visvesvara, G.S. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int. J. Parasitol. 2004, 34, 1001–1027. [Google Scholar] [CrossRef] [PubMed]
- Capewell, L.G.; Harris, A.M.; Yoder, J.S.; Cope, J.R.; Eddy, B.A.; Roy, S.L.; Visvesvara, G.S.; Fox, L.A.M.; Beach, M.J. Diagnosis, clinical course, and treatment of primary amoebic meningoencephalitis in the United States, 1937–2013. J. Pediatric Infect. Dis. Soc. 2015, 4, e68–e75. [Google Scholar] [CrossRef] [Green Version]
- Grace, E.; Asbill, S.; Virga, K. Naegleria fowleri: Pathogenesis, diagnosis, and treatment options. Antimicrob. Agents Chemother. 2015, 59, 6677–6681. [Google Scholar] [CrossRef] [Green Version]
- Anser, H.; Hasan, A. A Review on Global Distribution of Primary Amoebic Meningoencephalitis (PAM) Caused By Naegleria fowleri—The Brain Eating Amoeba. RADS J. Pharm. Pharm. Sci. 2018, 6, 95–99. [Google Scholar]
- Heggie, T.W.; Küpper, T. Surviving Naegleria fowleri infections: A successful case report and novel therapeutic approach. Travel Med. Infect. Dis. 2017, 16, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Linam, W.M.; Ahmed, M.; Cope, J.R.; Chu, C.; Visvesvara, G.S.; da Silva, A.J.; Qvarnstrom, Y.; Green, J. Successful Treatment of an Adolescent with Naegleria fowleri Primary Amebic Meningoencephalitis. Pediatrics 2015, 135, e744–e748. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.L.; Reed, T.; Stewart, C.; Levy, R.A. Naegleria fowleri That Induces Primary Amoebic Meningoencephalitis: Rapid Diagnosis and Rare Case of Survival in a 12-Year-Old Caucasian Girl. Lab. Med. 2016, 47, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Laniado-Laborín, R.; Cabrales-Vargas, M.N. Amphotericin B: Side effects and toxicity. Rev. Iberoam. Micol. 2009, 26, 223–227. [Google Scholar] [CrossRef]
- Benitez, L.L.; Carver, P.L. Adverse Effects Associated with Long-Term Administration of Azole Antifungal Agents. Drugs 2019, 79, 833–853. [Google Scholar] [CrossRef] [PubMed]
- Ellis, G.P. Chromenes, Chromanes, Chromones. In Chemistry of Heterocyclic Compounds; Ellis, G.P., Ed.; Wiley Interscience: New York, NY, USA, 1977; pp. 1–10. [Google Scholar]
- Schweizer, E.E.; Meeder-Nycz, O. 2H-and 4H-1-Benzopyrans. In Chemistry of Heterocyclic Compounds; Ellis, G.P., Ed.; Wiley Interscience: New York, NY, USA, 1977; pp. 11–139. [Google Scholar]
- Fravel, W.B.; Nedolya, N.A. Comprehensive Heterocyclic Chemistry III; Katritzky, A.R., Ramsden, C.A., Scriven, E.F.V., Taylor, R.J.K., Eds.; Elsevier: Oxford, UK, 2008; Volume 7, ISBN 9780080449913. [Google Scholar]
- Costa, M.; Dias, T.A.; Brito, A.; Proença, F. Biological importance of structurally diversified chromenes. Eur. J. Med. Chem. 2016, 123, 487–507. [Google Scholar] [CrossRef]
- Sugita, Y.; Takao, K.; Uesawa, Y.; Sakagami, H. Search for New Type of Anticancer Drugs with High Tumor Specificity and Less Keratinocyte Toxicity. Anticancer Res. 2017, 37, 5919–5924. [Google Scholar] [PubMed] [Green Version]
- Fuertes, M.; Masdeu, C.; Martin-Encinas, E.; Selas, A.; Rubiales, G.; Palacios, F.; Alonso, C. Synthetic Strategies, Reactivity and Applications of 1,5-Naphthyridines. Molecules 2020, 25, 3252. [Google Scholar] [CrossRef]
- Masdeu, C.; Fuertes, M.; Martin-Encinas, E.; Selas, A.; Rubiales, G.; Palacios, F.; Alonso, C. Fused 1,5-Naphthyridines: Synthetic Tools and Applications. Molecules 2020, 25, 3508. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Sharma, R.; Dhamodharan, V.; Pradeepkumar, P.I.; Manhas, A.; Srivastava, K.; Ahmed, S.; Kumar, N. Investigating Pharmacological Targeting of G-Quadruplexes in the Human Malaria Parasite. Biochemistry 2017, 56, 6691–6699. [Google Scholar] [CrossRef] [PubMed]
- Olepu, S.; Suryadevara, P.K.; Rivas, K.; Yokoyama, K.; Verlinde, C.L.M.J.; Chakrabarti, D.; Van Voorhis, W.C.; Gelb, M.H. 2-Oxo-tetrahydro-1,8-naphthyridines as selective inhibitors of malarial protein farnesyltransferase and as anti-malarials. Bioorg. Med. Chem. Lett. 2008, 18, 494–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundenapally, R.; Domala, R.; Bathula, S. Synthesis and Antibacterial Activity of Novel 5-arylidene-2-imino-3-(2-phenyl-1,8-naphthyridin-3-yl)thiazolidin-4-ones. Asian J. Pharm. Clin. Res. 2019, 12, 150–153. [Google Scholar] [CrossRef]
- Sato, Y.; Rokugawa, M.; Ito, S.; Yajima, S.; Sugawara, H.; Teramae, N.; Nishizawa, S. Fluorescent Trimethylated Naphthyridine Derivative with an Aminoalkyl Side Chain as the Tightest Non-aminoglycoside Ligand for the Bacterial A-site RNA. Chem. Eur. J. 2018, 24, 13862–13870. [Google Scholar] [CrossRef]
- Takahashi, N.; Hayashi, K.; Nakagawa, Y.; Furutani, Y.; Toguchi, M.; Shiozaki-Sato, Y.; Sudoh, M.; Kojima, S.; Kakeya, H. Development of an anti-hepatitis B virus (HBV) agent through the structure-activity relationship of the interferon-like small compound CDM-3008. Bioorganic Med. Chem. 2019, 27, 470–478. [Google Scholar] [CrossRef]
- Donalisio, M.; Massari, S.; Argenziano, M.; Manfroni, G.; Cagno, V.; Civra, A.; Sabatini, S.; Cecchetti, V.; Loregian, A.; Cavalli, R.; et al. Ethyl 1,8-Naphthyridone-3-carboxylates Downregulate Human Papillomavirus-16 E6 and E7 Oncogene Expression. J. Med. Chem. 2014, 57, 5649–5663. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Z.; Smith, S.J.; Métifiot, M.; Marchand, C.; Boyer, P.L.; Pommier, Y.; Hughes, S.H.; Burke, T.R. 4-Amino-1-hydroxy-2-oxo-1,8-naphthyridine-Containing Compounds Having High Potency against Raltegravir-Resistant Integrase Mutants of HIV-1. J. Med. Chem. 2014, 57, 5190–5202. [Google Scholar] [CrossRef]
- Gohil, J.D.; Patel, H.B.; Patel, M.P. Synthesis and evaluation of new chromene based [1,8]naphthyridines derivatives as potential antimicrobial agents. RSC Adv. 2016, 6, 74726–74733. [Google Scholar] [CrossRef]
- Martín-Encinas, E.; Rubiales, G.; Knudsen, B.R.; Palacios, F.; Alonso, C. Fused chromeno and quinolino[1,8]naphthyridines: Synthesis and biological evaluation as topoisomerase I inhibitors and antiproliferative agents. Bioorg. Med. Chem. 2021, 40, 116177. [Google Scholar] [CrossRef] [PubMed]
- Rizo-Liendo, A.; Sifaoui, I.; Arberas-Jiménez, I.; Reyes-Batlle, M.; Piñero, J.E.; Lorenzo-Morales, J. Fluvastatin and atorvastatin induce programmed cell death in the brain eating amoeba Naegleria fowleri. Biomed. Pharmacother. 2020, 130, 110583. [Google Scholar] [CrossRef] [PubMed]
- Tejería, A.; Pérez-Pertejo, Y.; Reguera, R.M.; Balaña-Fouce, R.; Alonso, C.; Fuertes, M.; González, M.; Rubiales, G.; Palacios, F. Antileishmanial effect of new indeno-1,5-naphthyridines, selective inhibitors of Leishmania infantum type IB DNA topoisomerase. Eur. J. Med. Chem. 2016, 124, 740–749. [Google Scholar] [CrossRef]
- Martín-Encinas, E.; Selas, A.; Tesauro, C.; Rubiales, G.; Knudsen, B.R.; Palacios, F.; Alonso, C. Synthesis of novel hybrid quinolino[4,3-b][1,5]naphthyridines and quinolino[4,3-b][1,5]naphthyridin-6(5H)-one derivatives and biological evaluation as topoisomerase I inhibitors and antiproliferatives. Eur. J. Med. Chem. 2020, 195, 112292. [Google Scholar] [CrossRef]
- Cárdenas-Zúñiga, R.; Silva-Olivares, A.; Villalba-Magdaleno, J.D.A.; Sánchez-Monroy, V.; Serrano-Luna, J.; Shibayama, M. Amphotericin B induces apoptosis-like programmed cell death in Naegleria fowleri and Naegleria gruberi. Microbiology 2017, 163, 940–949. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, G.; Zhang, P.; Fan, J. Programmed cell death and its role in inflammation. Mil. Med. Res. 2015, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullican, J.C.; Chapman, N.M.; Tracy, S. Complete genome sequence of the circular extrachromosomal element of Naegleria gruberi strain EGB ribosomal DNA. Genome Announc. 2018, 6, e00020-18. [Google Scholar] [CrossRef] [Green Version]
- Liechti, N.; Schürch, N.; Bruggmann, R.; Wittwer, M. Nanopore sequencing improves the draft genome of the human pathogenic amoeba Naegleria fowleri. Sci. Rep. 2019, 9, 16040. [Google Scholar] [CrossRef] [Green Version]
- Ali, I.K.M.; Kelley, A.; Joseph, S.J.; Park, S.; Roy, S.; Jackson, J.; Cope, J.R.; Rowe, L.A.; Burroughs, M.; Sheth, M.; et al. Draft Chromosome Sequences of a Clinical Isolate of the Free-Living Ameba Naegleria fowleri. Microbiol. Resour. Announc. 2021, 10, e01034-20. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, O.; Viayna, E.; Vicente-García, E.; Bartolini, M.; Ramón, R.; Juárez-Jiménez, J.; Clos, M.V.; Pérez, B.; Andrisano, V.; Luque, F.J.; et al. 1,2,3,4-Tetrahydrobenzo[h][1,6]naphthyridines as a new family of potent peripheral-to-midgorge-site inhibitors of acetylcholinesterase: Synthesis, pharmacological evaluation and mechanistic studies. Eur. J. Med. Chem. 2014, 73, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeouk, I.; Sifaoui, I.; Rizo-Liendo, A.; Arberas-Jiménez, I.; Reyes-Batlle, M.; Bazzocchi, I.; Bekhti, K.; Piñero, J.; Jiménez, I.A.; Lorenzo-Morales, J. Exploring the Anti-Infective Value of Inuloxin A Isolated from Inula viscosa against the Brain-Eating Amoeba (Naegleria fowleri) by Activation of Programmed Cell Death. ACS Chem. Neurosci. 2021, 12, 195–202. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (μM) ATCC® 30808 | IC50 (μM) ATCC® 30215 | CC50 (μM) ATCC® TIB-67 | SI (CC50/IC50) ATCC® TIB-67/ATCC® 30808 | IC90 (μM) ATCC® 30808 | IC90 (μM) ATCC® 30215 |
|---|---|---|---|---|---|---|
| 1 | >100 | N/D | N/D | N/D | N/D | N/D |
| 2 | >100 | N/D | N/D | N/D | N/D | N/D |
| 3 | 61.45 ± 5.27 | 76.61 ± 10.84 | >300 | >4.88 | 199.02 | >200 |
| 4 | >100 | N/D | N/D | N/D | N/D | N/D |
| 5 | >100 | N/D | N/D | N/D | N/D | N/D |
| 6 | >100 | N/D | N/D | N/D | N/D | N/D |
| 7 | >100 | N/D | N/D | N/D | N/D | N/D |
| 8 | >100 | N/D | N/D | N/D | N/D | N/D |
| 9 | >100 | N/D | N/D | N/D | N/D | N/D |
| 10 | >100 | N/D | N/D | N/D | N/D | N/D |
| 11 | >100 | N/D | N/D | N/D | N/D | N/D |
| 12 | >100 | N/D | N/D | N/D | N/D | N/D |
| 13 | >100 | N/D | N/D | N/D | N/D | N/D |
| 14 | >100 | N/D | N/D | N/D | N/D | N/D |
| 15 | >100 | N/D | N/D | N/D | N/D | N/D |
| 16 | >100 | N/D | N/D | N/D | N/D | N/D |
| 17 | >100 | N/D | N/D | N/D | N/D | N/D |
| 18 | >100 | N/D | N/D | N/D | N/D | N/D |
| 19 | >100 | N/D | N/D | N/D | N/D | N/D |
| 20 | >100 | N/D | N/D | N/D | N/D | N/D |
| 21 | >100 | N/D | N/D | N/D | N/D | N/D |
| 22 | >100 | N/D | N/D | N/D | N/D | N/D |
| 23 | >100 | N/D | N/D | N/D | N/D | N/D |
| 24 | >100 | N/D | N/D | N/D | N/D | N/D |
| Amphotericin B | 0.12 ± 0.03 | 0.16 ± 0.02 | ≥200 | ≥1652.89 | 0.35 ± 0.02 | 0.41 ± 0.12 |
| Miltefosine | 38.74 ± 4.23 | 81.57 ± 7.23 | 127.89 ± 8.85 | 3.301 | 89.47 ± 17.37 | >200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizo-Liendo, A.; Arberas-Jiménez, I.; Martin-Encinas, E.; Sifaoui, I.; Reyes-Batlle, M.; Chao-Pellicer, J.; Alonso, C.; Palacios, F.; Piñero, J.E.; Lorenzo-Morales, J. Naphthyridine Derivatives Induce Programmed Cell Death in Naegleria fowleri. Pharmaceuticals 2021, 14, 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101013
Rizo-Liendo A, Arberas-Jiménez I, Martin-Encinas E, Sifaoui I, Reyes-Batlle M, Chao-Pellicer J, Alonso C, Palacios F, Piñero JE, Lorenzo-Morales J. Naphthyridine Derivatives Induce Programmed Cell Death in Naegleria fowleri. Pharmaceuticals. 2021; 14(10):1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101013
Chicago/Turabian StyleRizo-Liendo, Aitor, Iñigo Arberas-Jiménez, Endika Martin-Encinas, Ines Sifaoui, María Reyes-Batlle, Javier Chao-Pellicer, Concepción Alonso, Francisco Palacios, José E. Piñero, and Jacob Lorenzo-Morales. 2021. "Naphthyridine Derivatives Induce Programmed Cell Death in Naegleria fowleri" Pharmaceuticals 14, no. 10: 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14101013