
Rhaponticum uniflorum and Serratula centauroides Extracts Attenuate Emotional Injury in Acute and Chronic Emotional Stress


 , ,
, ,
Abstract
:1. Introduction
2. Results
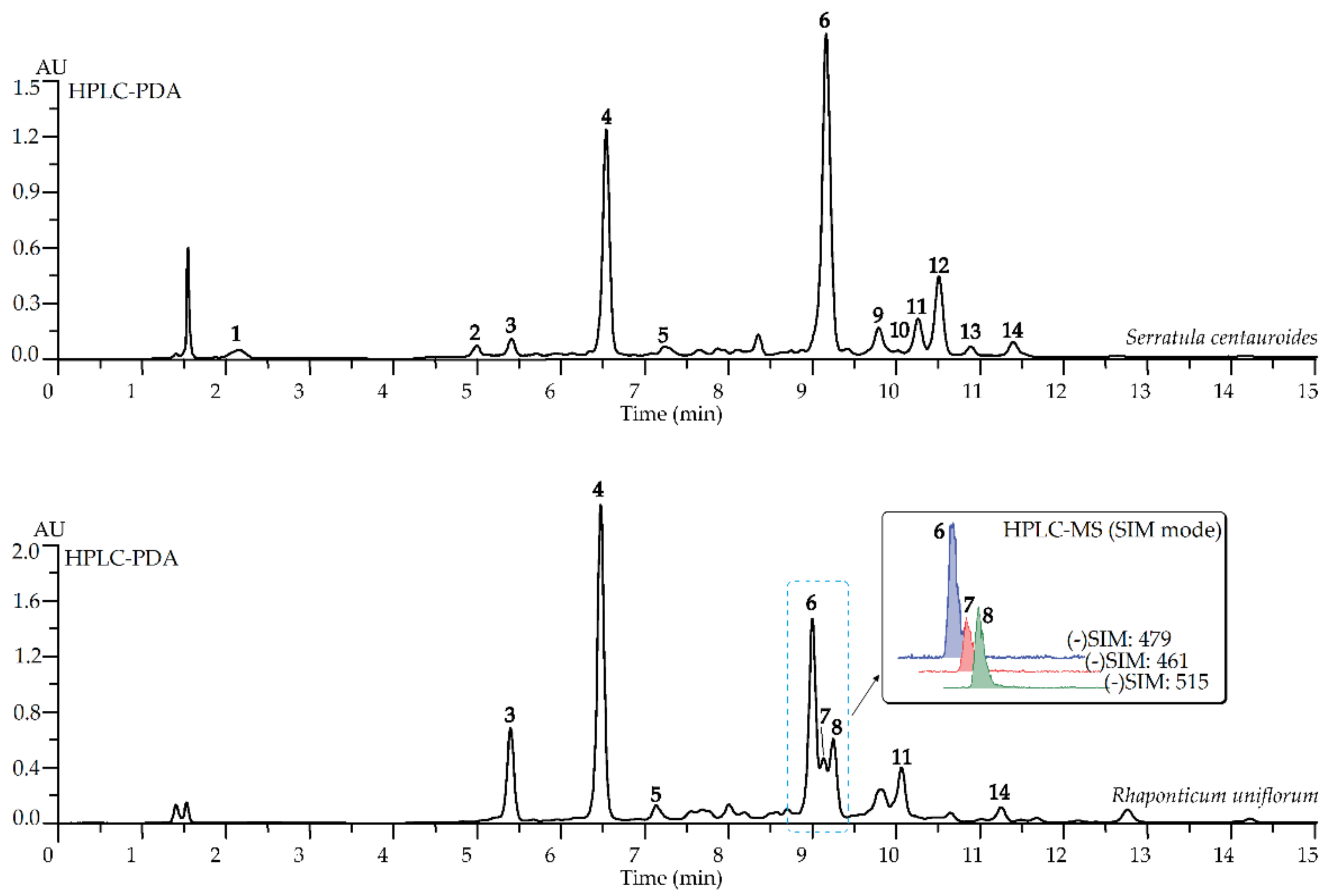
2.1. HPLC-DAD-ESI-MS Profiles of Rhaponticum uniflorum and Serratula centauroides Extracts
2.2. Acute Emotional Stress
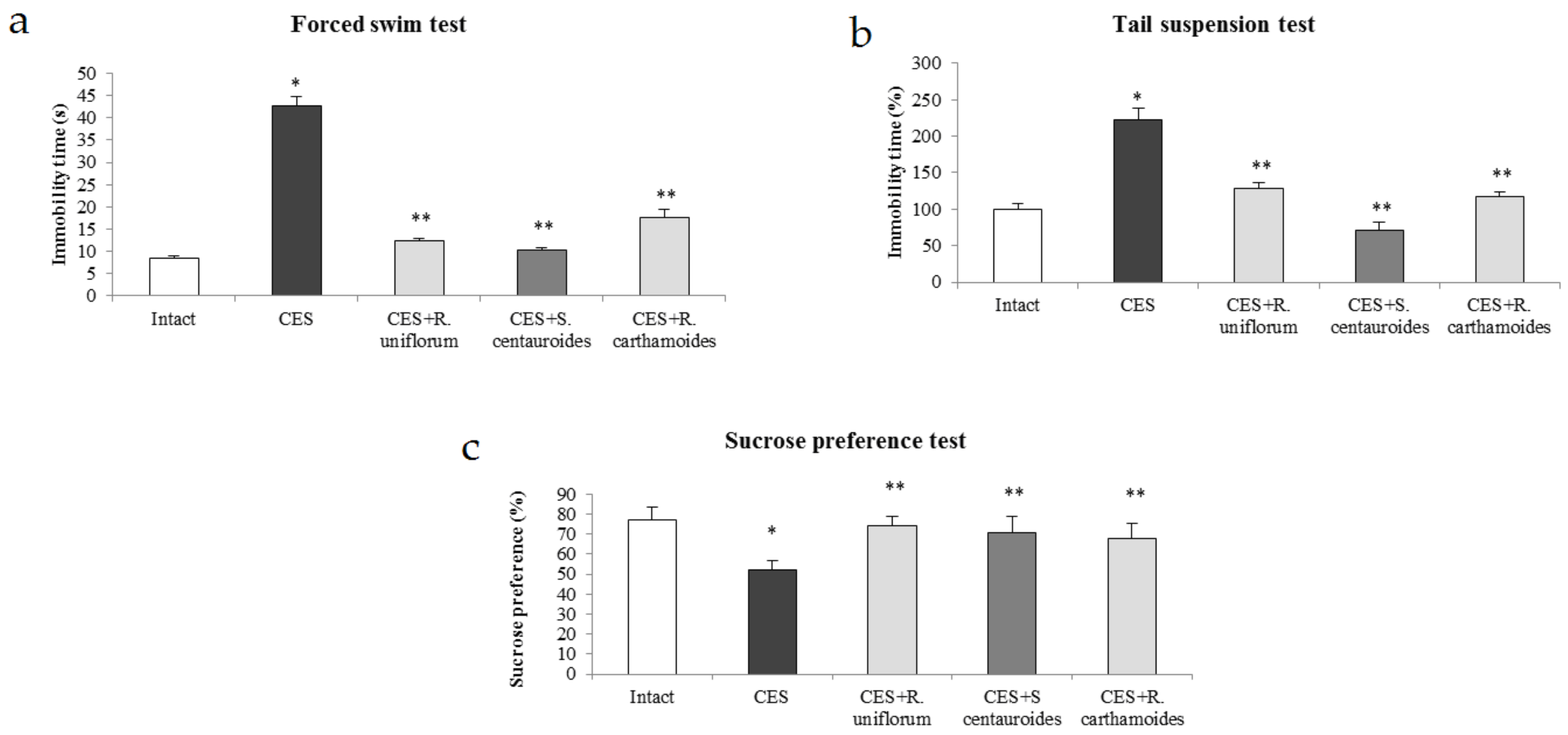
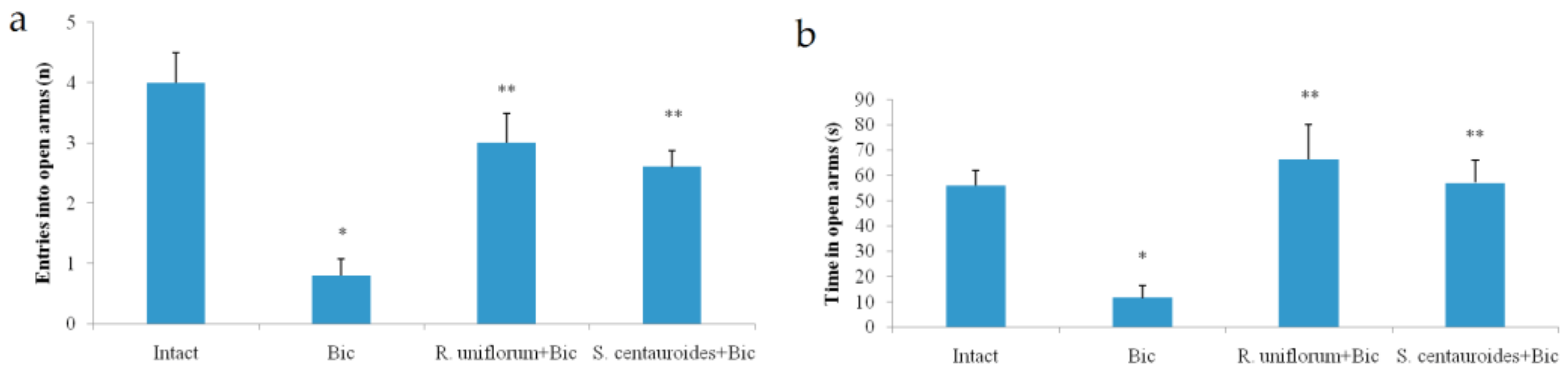
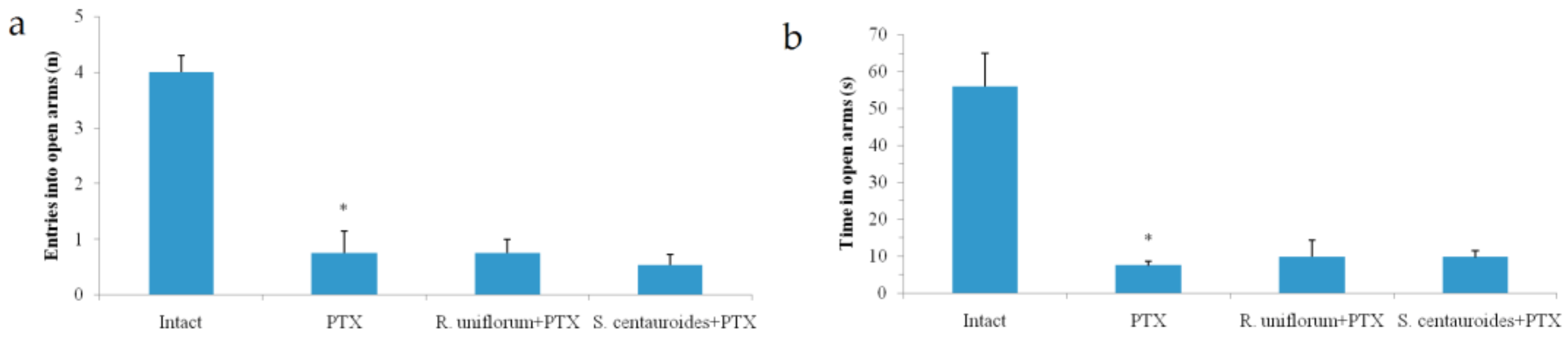
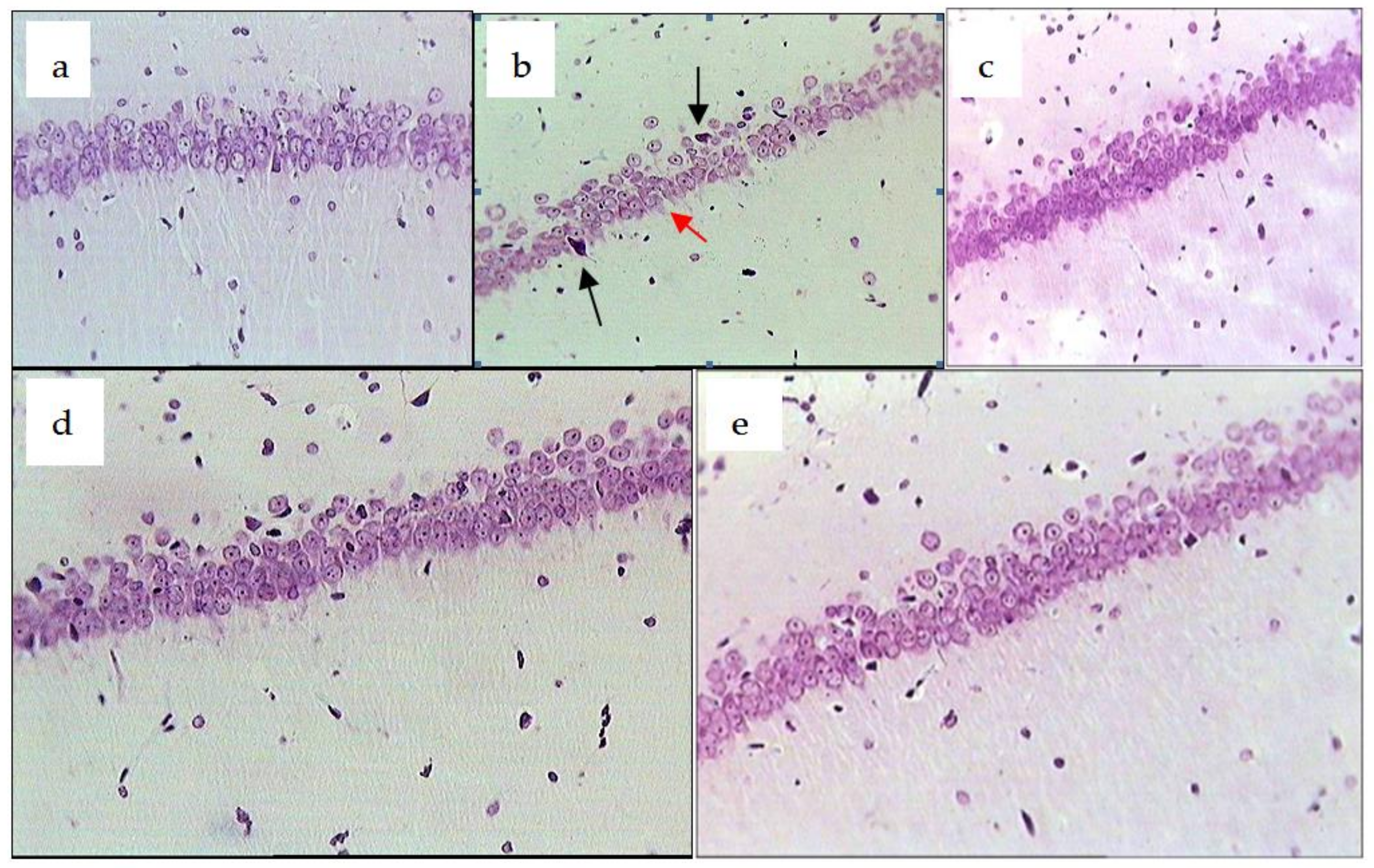
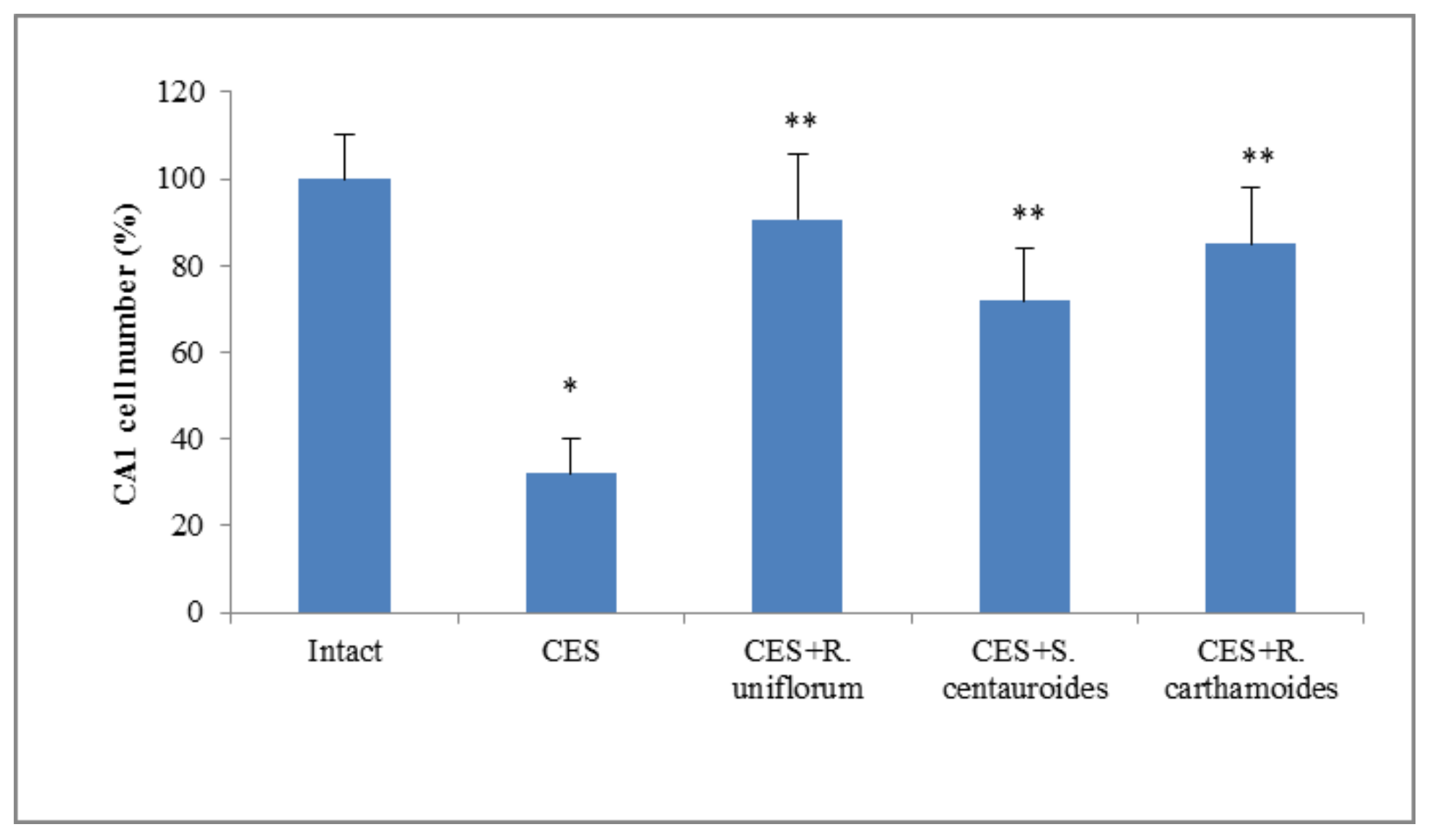
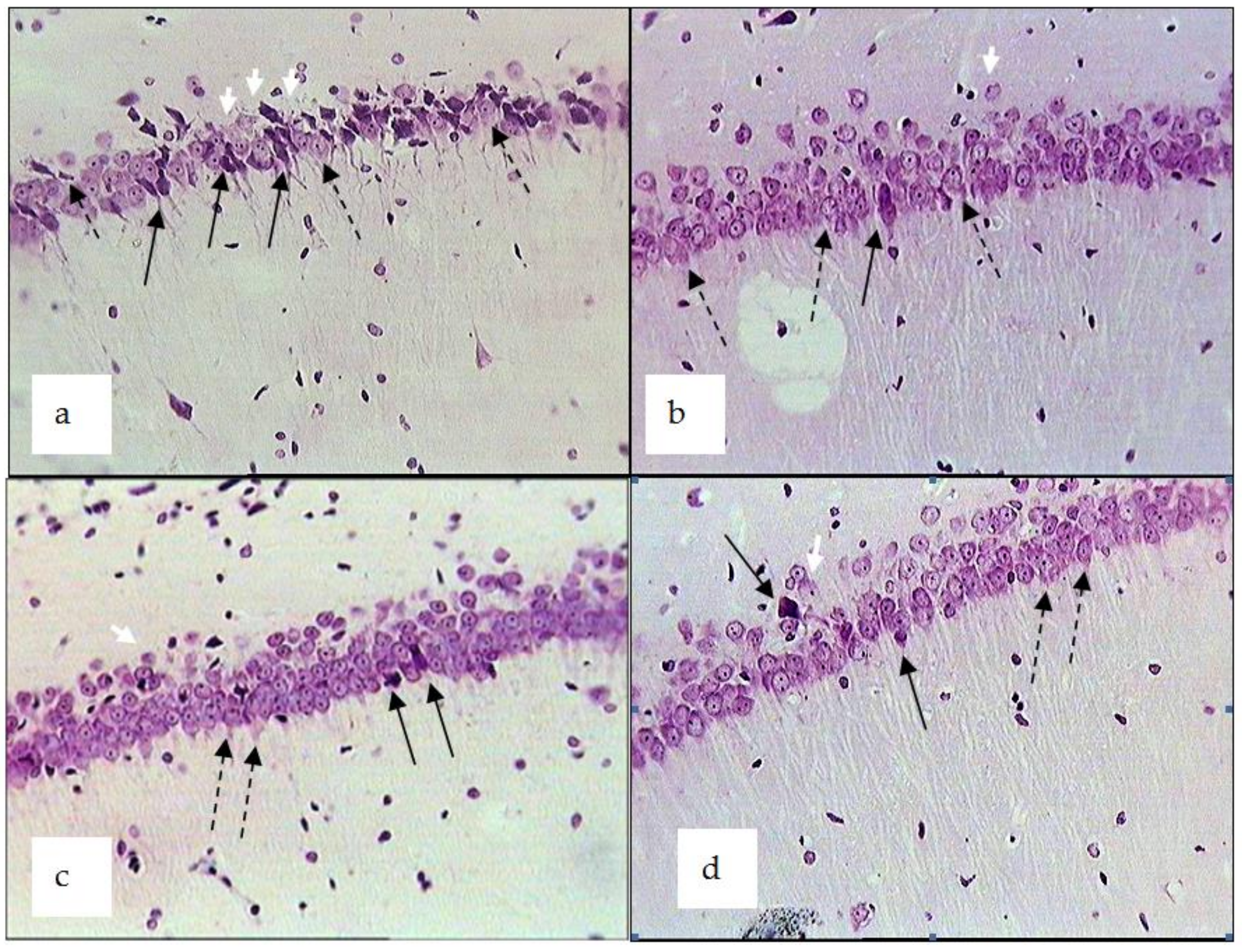
2.3. Chronic Emotional Stress
3. Discussion
4. Materials and Methods
4.1. Preparation of R. uniflorum and S. centauroides Extracts
4.2. High-Performance Liquid Chromatography with Photodiode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC-PDA-ESI-tQ-MS)
4.3. Laboratory Animals
4.4. Models, Methods and Experimental Protocols
4.4.1. Acute Emotional Stress
4.4.2. Chronic Emotional Stress
- day 1 of the week—placing of the rats one-by-one on a small elevated, round and dry platform (d 8 cm) located in a bath with cold water at 8–10 °C for 1 h;
- day 2—immobilisation of the rats in a small container for 1 h;
- day 3—nuchal fold suspension for 5 min.;
- day 4—placing of the rats in a cage with the floor awash with cold water for an hour;
- day 5—deprivation of water and food from 6 p.m. till 8 a.m.;
- day 6—placing of the rats in the home cage with a slope of 45° from 6 p.m. till 8 a.m.;
- day 7—no stress.
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panossian, A.; Seo, E.J.; Efferth, T. Novel molecular mechanisms for the adaptogenic effects of herbal extracts on isolated brain cells using systems biology. Phytomedicine 2018, 50, 257–284. [Google Scholar] [CrossRef] [PubMed]
- Addo, J.; Ayerbe, L.; Mohan, K.M.; Crichton, S.; Sheldenkar, A.; Chen, R.; Wolfe, C.B.; McKevitt, C. Socioeconomic status and stroke: An updated review. Stroke 2012, 43, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, J.D.; Ding, J.; Baum, L.; Sham, P.C. Systemic neuro-dysregulation in depression: Evidence from genome-wide association. Eur. Neuropsychopharmacol. 2020, 39, 1–18. [Google Scholar] [CrossRef]
- Fancourt, D.; Steptoe, A.; Bu, F. Trajectories of anxiety and depressive symptoms during enforced isolation due to COVID-19 in England: A longitudinal observational study. Lancet Psychiatry 2021, 8, 141–149. [Google Scholar] [CrossRef]
- Raihan, M.M.H. Mental health consequences of COVID-19 pandemic on adult population: A systematic review. Mental Health Rev. J. 2021, 26, 42–54. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G. Effects of Adaptogens on the central nervous system and the molecular mechanisms associated with their stress-protective activity. Pharmaceuticals 2010, 3, 188–224. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A. Understanding adaptogenic activity: Specificity of the pharmacological action of adaptogens and other phytochemicals. Ann. N. Y. Acad. Sci. 2017, 1401, 49–64. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Soleymani, A.; Cheng, Q. Traditional herbal medicines to overcome stress, anxiety and improve mental health in outbreaks of human coronaviruses. Phytother. Res. 2021, 35, 1237–1247. [Google Scholar] [CrossRef]
- Lafont, R. Recent progress in ecdysteroid pharmacology. Theor. Appl. Ecol. 2012, 1, 6–12. [Google Scholar] [CrossRef]
- Laekeman, G.; Vlietinck, A.J. Phytoecdysteroids: Phytochemistry and Pharmacological Activity. In Natural Products; Springer: Berlin, Heidelberg, Germany, 2013; pp. 3827–3849. [Google Scholar] [CrossRef]
- Volodin, V.; Chadin, I.; Whiting, P.; Dinan, L. Screening plants of European North-East Russia for ecdysteroids. Biochem. Syst. Ecol. 2002, 30, 525–578. [Google Scholar] [CrossRef]
- Lee, J.-H.; Hwang, K.H.; Kim, G.H. Inhibition of wild herb Rhaponticum uniflorum on synthesis of inflammatory mediators in macrophage cells. Food Sci. Biotechnol. 2013, 22, 567–572. [Google Scholar] [CrossRef]
- Chen, H.; Wang, C.; Qi, M.; Ge, L.; Tian, Z.; Li, J.; Zhang, M.; Wang, M.; Huang, L.; Tang, X. Anti-tumor effect of Rhaponticum uniflorum ethyl acetate extract by regulation of peroxiredoxin1 and epithelial-to-mesenchymal transition in oral cancer. Front. Pharmacol. 2017, 8, 870. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.P.; Jing, Z.; Dong, M.; Zhang, M.-L.; Huo, C.-H.; Shi, Q.-W.; Gu, Y.-C. Chemical Constituents of Plants from the Genus Rhaponticum. Chem. Biodiv. 2010, 7, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva, I.G.; Tsybiktarova, L.P.; Garmaeva, L.L.; Nikolaeva, G.G.; Olennikov, D.N.; Matkhanov, I.E. Determination of ecdysteroids in Fornicium unflorum (L.) and Serratula centauroides (L.) raw materials by chromatography-UV spectrophotometry. J. Anal. Chem. 2017, 72, 854–861. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. New flavonoids and turkesterone-2-O-cinnamate from leaves of Rhaponticum uniflorum. Chem. Nat. Compd. 2019, 55, 256–264. [Google Scholar] [CrossRef]
- Piao, L.; Zhang, X.W.; Jin, X.Z. Anti-senile effect of Rhaponticum uniflorm (L.) DC. extract on D-galactose induced senile in rats. Lishizhen Med. Mater. Med. Res. 2006, 10, 1918–1919. [Google Scholar]
- Olennikov, D.N.; Kashchenko, N.I. Phytoecdysteroids of Serratula centauroides Herb from Cisbaikalia. Russ. J. Bioorg. Chem. 2019, 45, 913–919. [Google Scholar] [CrossRef]
- Nikolaev, S.M.; Nikolaeva, I.G.; Razuvaeva, Y.G.; Matkhanov, I.E.; Tsybiktarova, L.P.; Shantanova, L.N.; Nikolaeva, G.G. Phenolic compounds of Serratula centauroides and anxiolytic effect. Farmacia 2019, 67, 504–510. [Google Scholar] [CrossRef]
- Paoletti, F.; Mocali, A. Determination of superoxide dismutase activity by purely chemical system based on NAD(P)H oxidation. Methods Enzymol. 1990, 186, 209–220. [Google Scholar] [CrossRef]
- Liao, L.Y.; He, Y.F.; Li, L.; Meng, H.; Dong, Y.M.; Yi, F.; Xiao, P.G. A preliminary review of studies on adaptogens: Comparison of their bioactivity in TCM with that of ginseng-like herbs used worldwide. Chin. Med. 2018, 13, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.; et al. Evolution of the adaptogenic concept from traditional use to medical systems: Pharmacology of stress- and aging-related diseases. Med. Res. Rev. 2021, 41, 630–703. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.M.; Alsawah, F.; McNeill, P.C.; Galloway, M.P.; Perrine, S.A. Neurochemical, hormonal, and behavioral effects of chronic unpredictable stress in the rat. Behav. Brain Res. 2011, 220, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Pshennikova, M.G. The phenomenon of stress: Emotional stress and its role in pathology. In Aktual’nye Problemy Patofiziologii: Izbrannye Leklii; Moioza, B.B., Ed.; Meditsina: Moscow, Russia, 2001; pp. 220–353. [Google Scholar]
- Arushanyan, E.B.; Beyer, E.V. Plant Adaptogens; Izd-vo SGMA: Stavropol, Russia, 2009; Volume 232. [Google Scholar]
- Wu, J.M.; Gao, L.; Shang, L.; Wang, G.H.; Wei, N.N.; Chu, T.T.; Chen, S.P.; Zhang, Y.J.; Huang, J.; Wang, J.H.; et al. Ecdysterones from Rhaponticum carthamoides (Willd.) Iljin reduce hippocampal excitotoxic cell loss and upregulate mTOR signaling in rats. Fitoterapia 2017, 119, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, B. Stress preferentially increases extraneuronal levels of excitatory amino acids in the prefrontal cortex: Comparison to hippocampus and basal ganglia. J. Neurochem. 1993, 60, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Heuser, I.; Lammers, C.-H. Stress and the brain. Neurobiol. Aging 2003, 24, S69–S76. [Google Scholar] [CrossRef]
- Hu, J.; Luo, C.X.; Chu, W.H.; Shan, Y.A.; Qian, Z.M.; Zhu, G.; Yu, Y.B.; Feng, H. 20-Hydroxyecdysone protects against oxidative stress-induced neuronal injury by scavenging free radicals and modulating NF-kappa and JNK pathways. PLoS ONE 2012, 7, e50764. [Google Scholar] [CrossRef]
- Miyamae, Y.; Kurisu, M.; Han, J.; Isoda, H.; Shigemori, H. Structure–activity relationship of caffeoylquinic acids on the accelerating activity on ATP production. Chem. Pharm. Bull. 2011, 59, 502–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef] [PubMed]
- Overmier, J.; Murison, R.; Ursin, H. The ulcerogenic effect of a rest period after exposure to water–restraint stress. Behav. Neural. Biol. 1986, 46, 372–386. [Google Scholar] [CrossRef]
- Kikugawa, K.; Kojima, T.; Yamaki, S.; Kosugi, H. Interpretation of the thiobarbituric acid reactivity of rat liver and brain homogenates in the presence of ferric ion and ethylenediaminetetraacetic acid. Anal. Biochem. 1992, 202, 249–255. [Google Scholar] [CrossRef]
- Akerboom, T.P.; Sies, H. Assay of glutathione, glutathione disulfide, and glutathione mixed disulfides in biological samples. Methods Enzymol. 1981, 77, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Okhlopkova, Z.M.; Zulfugarov, I.S. Chemical composition and antioxidant activity of Tánara Ótó (Dracocephalum palmatum Stephan), a medicinal plant used by the North-Yakutian nomads. Molecules 2013, 18, 14105–14121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyut, Z.; Beydemir, S.; Gulcin, I. Antioxidant and antiradical properties of selected flavonoids and phenolic compounds. Biochem. Res. Int. 2017, 2017, 7616791. [Google Scholar] [CrossRef]
- Ozen, T.; Demirtas, I.; Aksit, H. Determination of antioxidant activities of various extracts and essential oil compositions of Thymus praecox subsp. skorpilii var. skorpilii. Food Chem. 2011, 124, 58–64. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Agafonova, S.V.; Stolbikova, A.V.; Rokhin, A.V. Melanin of Laetiporus sulphureus (Bull: Fr.) Murr sterile form. Appl. Biochem. Microbiol. 2011, 47, 298–303. [Google Scholar] [CrossRef]
- Papp, M.; Willner, P.; Muscat, R. An animal model of anhedonia: Attenuation of sucrose consumption and place preference conditioning by chronic unpredictable emotional stress. Psychopharmacology 1991, 104, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The open field test. In Mood and Anxiety Related Phenotypes in Mice. Neuromethods; Gould, T.D., Ed.; Humana Press Inc.: New York, NY, USA, 2009; pp. 1–20. [Google Scholar] [CrossRef]
- Pellow, S.; File, S.E. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: A novel test of anxiety in the rat. Pharmacol. Biochem. Behav. 1986, 24, 525–529. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Anton, G.; Blavet, N.; Jalfre, M. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharmacol. 1978, 47, 379–391. [Google Scholar] [CrossRef]
- Avtandilov, G.G. Medical Morphometry: Handbook; Meditsina: Moscow, Russia, 1990; pp. 115–117. [Google Scholar]
- Jung, Y.H.; Ha, R.R.; Kwon, S.H.; Hong, S.I.; Lee, K.H.; Kim, S.Y.; Lee, S.Y.; Jang, C.G. Anxiolytic effects of julibroside C1 isolated from Albizzia julibrissin in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 44, 184–192. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | tR, min | Compound 1 | UV, λmax, nm | ESI-MS, m/z | Content in Extract, mg/g ± S.D. | |
|---|---|---|---|---|---|---|
| R. uniflorum | S. centauroides | |||||
| 1 | 2.11 | Arbutin S | 283 | 271 [M–H]− | n.d. | 24.78 ± 0.49 |
| 2 | 5.02 | 1-O-Caffeoylquinic acid S | 327 | 353 [M–H]− | n.d. | 1.82 ± 0.03 |
| 3 | 5.48 | 4-O-Caffeoylquinic acid S | 327 | 353 [M–H] − | 25.11 ± 0.50 | 4.19 ± 0.08 |
| 4 | 6.52 | 5-O-Caffeoylquinic acid S | 327 | 353 [M–H]− | 63.59 ± 1.27 | 40.85 ± 0.82 |
| 5 | 7.18 | 1,3-Di-O-caffeoylquinic acid S | 328 | 515 [M–H]− | 2.97 ± 0.06 | 2.16 ± 0.04 |
| 6 | 9.11 | 20-Hydroxyecdysone S | 246 | 519 [M+K]+, 503 [M+Na]+, 481 [M+H]+, 463 [(M+H)–H2O]+, 445 [(M+H)–2 × H2O]+, 427 [(M+H)–3 × H2O]+, 409 [(M+H)–4 × H2O]+ | 17.83 ± 0.35 | 27.43 ± 0.55 |
| 7 | 9.22 | Luteolin-7-O-glucuronide S | 347 | 461 [M–H]−, 285 [(M–GlcA)–H]− | 12.18 ± 0.24 | n.d. |
| 8 | 9.42 | 3,4-Di-O-caffeoylquinic acid S | 328 | 515 [M–H]− | 14.15 ± 0.28 | n.d. |
| 9 | 9.82 | Inokosterone S | 245 | 519 [M+K]+, 503 [M+Na]+, 481 [M+H]+, 463 [(M+H)–H2O]+, 445 [(M+H)–2×H2O]+, 427 [(M+H)–3×H2O]+, 409 [(M+H)–4×H2O]+ | n.d. | 5.68 ± 0.11 |
| 10 | 9.92 | 3,5-Di-O-caffeoylquinic acid S | 328 | 515 [M–H]− | n.d. | n.d. |
| 11 | 10.18 | Apigenin-7-O-glucuronide S | 337 | 445 [M–H]−, 269 [(M–GlcA)–H]- | 16.93 ± 0.37 | 18.27 ± 0.36 |
| 12 | 10.49 | Chrysoeriol-7-O-glucuronide L | 347 | 475 [M–H]−, 299 [(M–GlcA)–H]- | n.d. | 16.90 ± 0.31 |
| 13 | 10.92 | 4,5-Di-O-caffeoylquinic acid S | 328 | 515 [M–H]− | n.d. | 1.53 ± 0.03 |
| 14 | 11.28 | 1,5-Di-O-caffeoylquinic acid S | 328 | 515 [M–H]− | 2.35 ± 0.05 | 1.93 ± 0.04 |
| Parameter | Experimental Groups | ||||
|---|---|---|---|---|---|
| Intact (no AES) | AES + Saline | AES + R. uniflorum | AES + S. centauroides | AES + R. carthamoides | |
| Thymus weight, mg/100g | 57.31 ± 5.23 | 40.54 ± 2.10 * | 56.10 ± 3.78 ** | 55.52 ± 3.61 ** | 49.50 ± 2.53 ** |
| Adrenal glands weight, mg/100g | 16.04 ± 1.08 | 25.02 ± 2.51 * | 20.31 ± 1.66 | 15.81 ± 1.98 ** | 16.33 ± 1.62 ** |
| TBARS, µM/L | 12.20 ± 1.03 | 24.71 ± 1.41 * | 15.30 ± 0.78 ** | 16.63 ± 1.37 ** | 14.30 ± 0.92 ** |
| Reduced glutathione, mM/L | 3.12 ± 0.16 | 0.80 ± 0.73 * | 2.33 ± 0.17 ** | 2.74 ± 0.04 ** | 1.52 ± 0.08 ** |
| Catalase, U/mL | 8.31 ± 0.61 | 5.92 ± 0.48 * | 7.31 ± 0.46 ** | 7.50 ± 0.11 ** | 6.51 ± 0.72 ** |
| Superoxide dismutase, U/mL | 15.64 ± 1.08 | 6.21 ± 0.57 * | 9.72 ± 0.05 ** | 11.32 ± 0.79 ** | 6.84 ± 0.27 ** |
| Extract, Compound | DPPH˙, IC50, µg/mL | ABTS•+, IC50, µg/mL | O2•-, IC50, µg/mL | FeCA, IC50, µg/mL |
|---|---|---|---|---|
| R. uniflorum | 22.80 ± 1.32 * | 15.50 ± 1.11 * | 55.40 ± 4.10 * | 546.51 ± 24.20 * |
| S. centauroides | 30.12 ± 1.61 * | 25.14 ± 1.92 * | 77.31 ± 5.22 * | 819.20 ± 41.32 * |
| Ascorbic acid | 4.84 ± 0.11 | 9.11 ± 0.92 | 101.10 ± 5.14 | 150.25 ± 10.11 |
| Parameter | Experimental Groups | ||||
|---|---|---|---|---|---|
| Intact (no AES) | AES + Saline | AES + R. Uniflorum | AES + S. Centauroides | AES + R. Carthamoides | |
| Epinephrine, nM | 8.51 ± 0.59 | 37.81 ± 0.35 * | 26.50 ± 1.24 ** | 24.33 ± 0.89 ** | 31.61 ± 0.51 ** |
| Norepinephrine, nM | 64.12 ± 0.27 | 120.63 ± 4.71 | 87.71 ± 5.35 ** | 103.80 ± 3.15 | 111.32 ± 3.77 |
| ACTH, pg/mL | 15.84 ± 1.69 | 51.03 ± 4.27 * | 31.22 ± 0.86 ** | 37.42 ± 0.69 ** | 42.60 ± 2.12 ** |
| Corticosterone, nM | 44.30 ± 3.74 | 65.72 ± 3.80 * | 50.52 ± 2.45 ** | 50.61 ± 3.83 ** | 54.74 ± 4.32 |
| Aldosterone, pg/mL | 271.11 ± 10.42 | 296.10 ± 11.74 * | 226.13 ± 12.4 ** | 278.04 ± 17.6 | 257.05 ± 16.61 |
| Parameter | Experimental Groups | |||||
|---|---|---|---|---|---|---|
| Intact (no CES) | CES + saline | CES + R. uniflorum | CES + S. centauroides | CES + R. carthamoides | ||
| Open Field | ||||||
| Crossing | 29.91 ± 3.01 | 17.93 ± 1.44 * | 27.90 ± 4.02 ** | 31.85 ± 4.80 ** | 24.33 ± 2.31 | |
| Rearing | 6.10 ± 0.82 | 3.40 ± 0.05 * | 14.35 ± 0.81 ** | 12.63 ± 2.50 ** | 4.12 ± 0.31 | |
| Peeking in the holes | 4.05 ± 0.16 | 0.61 ± 0.01 * | 2.32 ± 0.13 ** | 3.10 ± 0.08 ** | 2.55 ± 0.17 ** | |
| Boli | 1.33 ± 0.10 | 2.21 ± 0.12 * | 2.10 ± 0.14 | 1.53 ± 0.15 ** | 1.84 ± 0.10 ** | |
| Elevated plus maze test | ||||||
| Number of entries | open arm | 1.90 ± 0.27 | 0.25 ± 0.13 * | 1.75 ± 0.20 ** | 2.11 ± 0.41 ** | 0.53 ± 0.12 ** |
| closed arm | 2.83 ± 0.34 | 0.72 ± 0.21 * | 1.84 ± 0.12 ** | 2.20 ± 0.25 ** | 2.21 ± 0.04 ** | |
| Duration of stay | open arm | 70.32 ± 5.61 | 13.21 ± 2.12 * | 39.11 ± 5.63 ** | 48.71 ± 2.91 ** | 22.80 ± 2.05 ** |
| closed arm | 216.1 ± 30.10 | 280.2 ± 5.60* | 239.1 ± 15.05 ** | 250.0 ± 10.04 ** | 259.2 ± 6.10 ** | |
| Experimental Groups | Hippocampus Neurons (CA1), LH (%) | Hippocampus Neurons (Ca1), RH (%) | ||||
|---|---|---|---|---|---|---|
| Hyperchromic | Dystrophic | Normal | Hyperchromic | Dystrophic | Normal | |
| Intact | 1.32 ± 0.41 | 2.33 ± 0.70 | 96.71 ± 0.62 | 1.44 ± 0.74 | 2.40 ± 0.64 | 96.25 ± 8.12 |
| Sham | 3.50 ± 0.51 | 17.51 ± 1.53 | 79.03 ± 3.01 | 4.81 ± 0.50 | 12.10 ± 0.91 | 82.13 ± 1.71 |
| Control (CES + ischemia) | 64.11 ± 8.52 * | 15.80 ± 2.04 | 20.12 ± 4.11 * | 21.41 ± 2.02 * | 17.43 ± 3.25 | 60.81 ± 8.15 * |
| CES + ischemia + R. uniflorum | 44.12 ± 8.87 ** | 25.92 ± 5.33 ** | 29.14 ± 4.05 ** | 23.06 ± 2.41 ** | 5.44 ± 3.21 | 71.53 ± 10.31 |
| CES + ischemia + S. centauroides | 41.11 ± 9.80 ** | 21.94 ± 2.37 | 37.10 ± 5.06 ** | 13.42 ± 2.04 ** | 17.41 ± 3.24 | 69.22 ± 10.21 |
| CES + ischemia + R. carthamoides | 39.20 ± 7.10 ** | 19.90 ± 4.18 ** | 40.95 ± 3.72 ** | 15.51 ± 3.01 | 2.85 ± 4.72 ** | 81.70 ± 9.90 |
| Compound | Ionization a | Optimised MRM Transitions, m/z | CE b (eV) | Regression Equation c | r2 | SYX | LOD/ LOQ (µg/mL) | Linear Range (µg/mL) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Precursor | Quantifier | a | b∙106 | |||||||
| Arbutin | N | 271 | 107 | −10 | 0.1756 | −0.0144 | 0.9967 | 3.01∙10−2 | 0.56/1.71 | 2.00–850.0 |
| 1-O-Caffeoylquinic acid | N | 353 | 127 | −15 | 2.5394 | −1.2360 | 0.9994 | 0.45∙10−2 | 0.006/0.02 | 0.02–300.0 |
| 4-O-Caffeoylquinic acid | N | 353 | 173 | −15 | 2.7365 | −1.0690 | 0.9996 | 0.51∙10−2 | 0.006/0.02 | 0.02–300.0 |
| 5-O-Caffeoylquinic acid | N | 353 | 165 | −15 | 2.9021 | −1.4184 | 0.9998 | 0.39∙10−2 | 0.004/0.01 | 0.02–300.0 |
| 1,3-O-Caffeoylquinic acid | N | 515 | 179 | −15 | 2.4176 | −1.5647 | 0.9994 | 0.40∙10−2 | 0.005/0.02 | 0.02–300.0 |
| 20-Hydroxyecdysone | P | 481 | 463 | +20 | 1.6705 | −0.4374 | 0.9988 | 12.79∙10−2 | 0.25/0.77 | 0.8–100.0 |
| Luteolin-7-O-glucuronide | N | 461 | 285 | −20 | 1.4412 | −0.6211 | 0.9930 | 11.25∙10−2 | 0.26/0.78 | 0.80–100.0 |
| 3,4-Di-O-caffeoylquinic acid | N | 515 | 191 | −15 | 1.1541 | −0.4691 | 0.9987 | 1.06∙10−2 | 0.03/0.10 | 0.10–350.0 |
| 3,5-Di-O-caffeoylquinic acid | N | 515 | 173 | −20 | 0.9562 | −0.0521 | 0.9971 | 7.79∙10−2 | 0.27/0.82 | 0.90–100.0 |
| Apigenin-7-O-glucuronide | N | 445 | 269 | −15 | 7.064 | −1.533 | 0.9992 | 1.92∙10−2 | 0.009/0.03 | 0.03–500.0 |
| Chrysoeriol-7-O-glucoside | N | 461 | 299 | −20 | 2.0384 | −0.3640 | 0.9975 | 2.02∙10−2 | 0.03/0.10 | 0.10–350.0 |
| 4,5-Di-O-caffeoylquinic acid | N | 515 | 203 | −15 | 3.6748 | −0.7069 | 0.9987 | 0.90∙10−2 | 0.008/0.02 | 0.02–400.0 |
| 1,5-Di-O-caffeoylquinic acid | N | 515 | 191 | −20 | 1.4689 | −0.3641 | 0.9990 | 5.69∙10−2 | 0.12/0.38 | 0.40–400.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shantanova, L.N.; Olennikov, D.N.; Matkhanov, I.E.; Gulyaev, S.M.; Toropova, A.A.; Nikolaeva, I.G.; Nikolaev, S.M. Rhaponticum uniflorum and Serratula centauroides Extracts Attenuate Emotional Injury in Acute and Chronic Emotional Stress. Pharmaceuticals 2021, 14, 1186. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111186
Shantanova LN, Olennikov DN, Matkhanov IE, Gulyaev SM, Toropova AA, Nikolaeva IG, Nikolaev SM. Rhaponticum uniflorum and Serratula centauroides Extracts Attenuate Emotional Injury in Acute and Chronic Emotional Stress. Pharmaceuticals. 2021; 14(11):1186. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111186
Chicago/Turabian StyleShantanova, Larisa N., Daniil N. Olennikov, Irinchey E. Matkhanov, Sergey M. Gulyaev, Anyuta A. Toropova, Irina G. Nikolaeva, and Sergey M. Nikolaev. 2021. "Rhaponticum uniflorum and Serratula centauroides Extracts Attenuate Emotional Injury in Acute and Chronic Emotional Stress" Pharmaceuticals 14, no. 11: 1186. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111186