Targeting Oxidative Stress, NLRP3 Inflammasome, and Autophagy by Fraxetin to Combat Doxorubicin-Induced Cardiotoxicity


Abstract
:1. Introduction
2. Methods and Materials
2.1. Chemicals and Drugs
2.2. Animals and Groups
2.3. Assessment of Cardiac Function Tests in Blood Samples
2.4. Processing and Preparation of Cardiac Tissues
2.5. Evaluation of Oxidative Stress Parameters and Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2) Content in Cardiac Tissues
2.6. Determination of Cardiac Tissue Transforming Growth Factor Beta 1 (TGF-β1), Interleukin 10 (IL-10), Interferon Alpha (IFN-α), and Nucleotide-Binding Domain-Like Receptor Family, Pyrin Domain-Containing 3 (NLRP3) Inflammasome
2.7. Assay of p38 Mitogen-Activated Protein (p-MAP) Kinase, Phosphoprotein Kinase B (p-AKT), and Phospho-Mammalian Target of Rapamycin (p-mTOR) in Cardiac Tissues
2.8. Quantification of Beclin-1, Nerve Growth Factor Beta (NGF-β) and c-Jun NH2-Terminal Kinase (JNK) Activity in Cardiac Tissues
2.9. In Vivo Echocardiographic Examination of Rats of the Different Studied Groups
2.10. Evaluation of the Histopathological Changes in Cardiac Tissues
2.11. Immunohistochemical Staining of Cardiac Tissues for Assessment of Caspase-3
2.12. Assessment of the Electron Microscopic Changes of the Cardiac Tissues
2.13. Statistical Analysis of the Obtained Data
3. Results
3.1. Fraxetin, in a Dose-Dependent Manner, Combatted the Changes Induced by Doxorubicin in Cardiac Function Tests
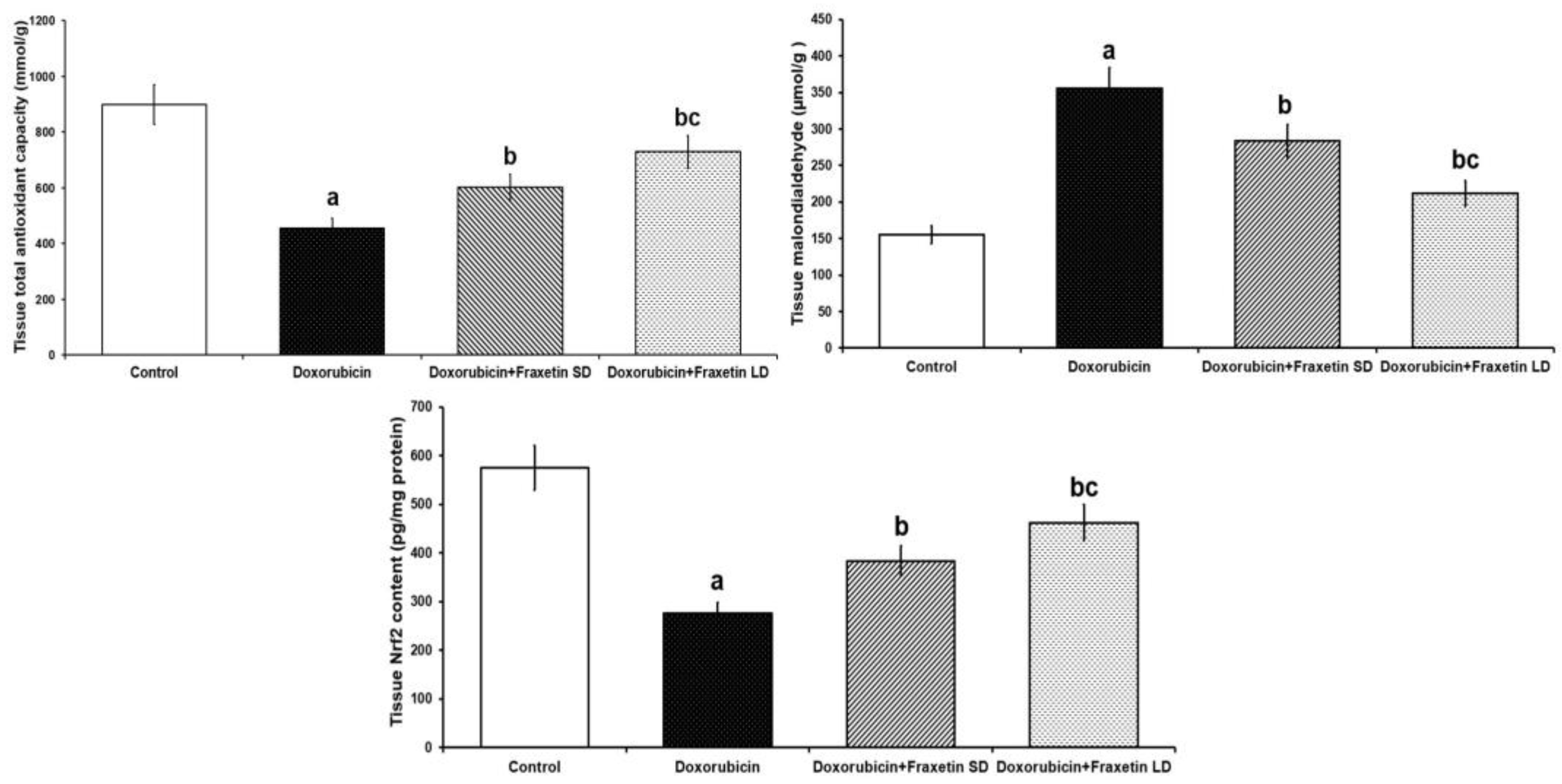
3.2. Fraxetin Augmented the Antioxidant Defense Mechanisms and Restored Nrf2 Content of Cardiac Tissues in Doxorubicin-Treated Rats
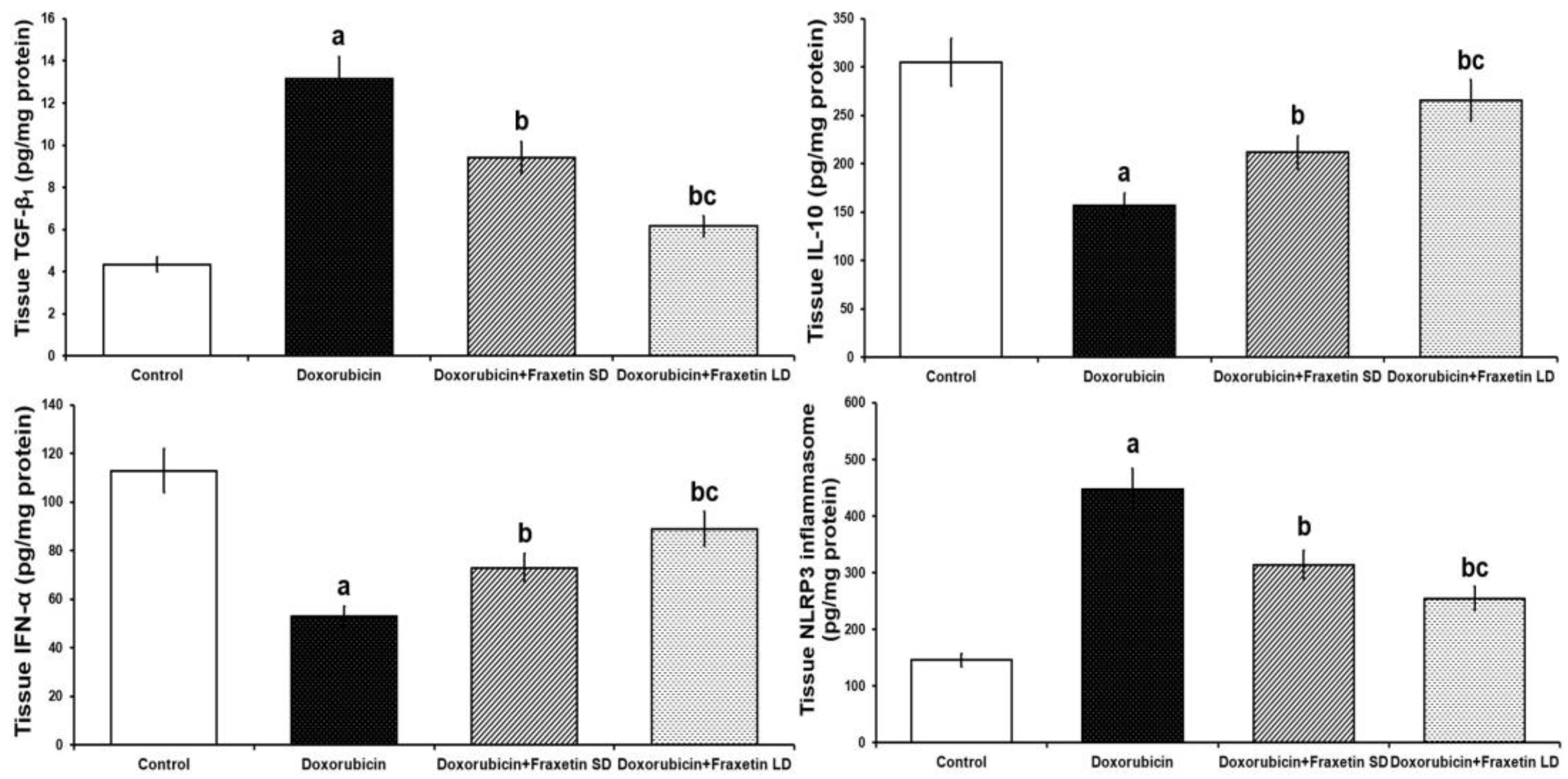
3.3. Fraxetin Mitigated the Changes Induced by Doxorubicin in Cardiac Tissue TGF-β1, IL-10, IFN-α, and NLRP3 Inflammasome
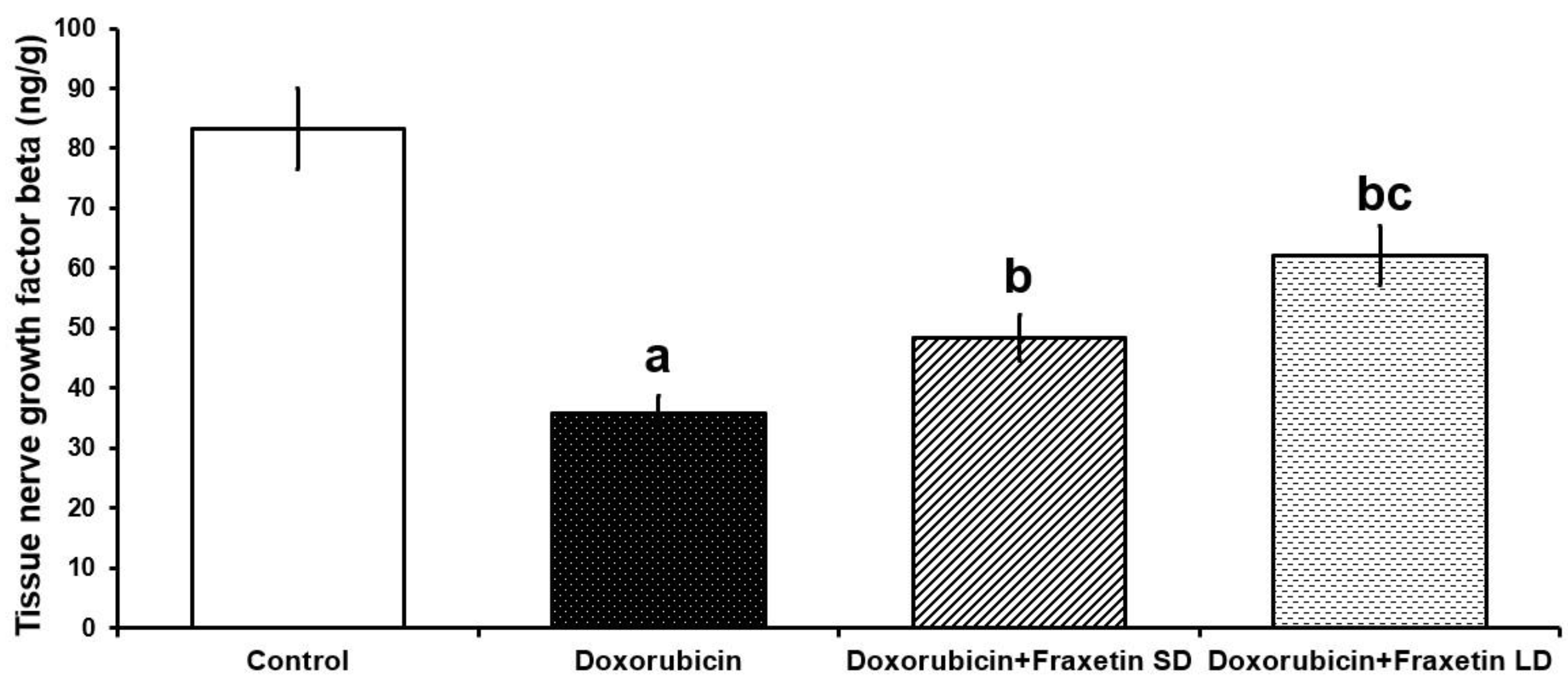
3.4. Fraxetin Augmented the Autophagy-Associated Pathways in the Cardiac Tissues of Doxorubicin-Treated Rats
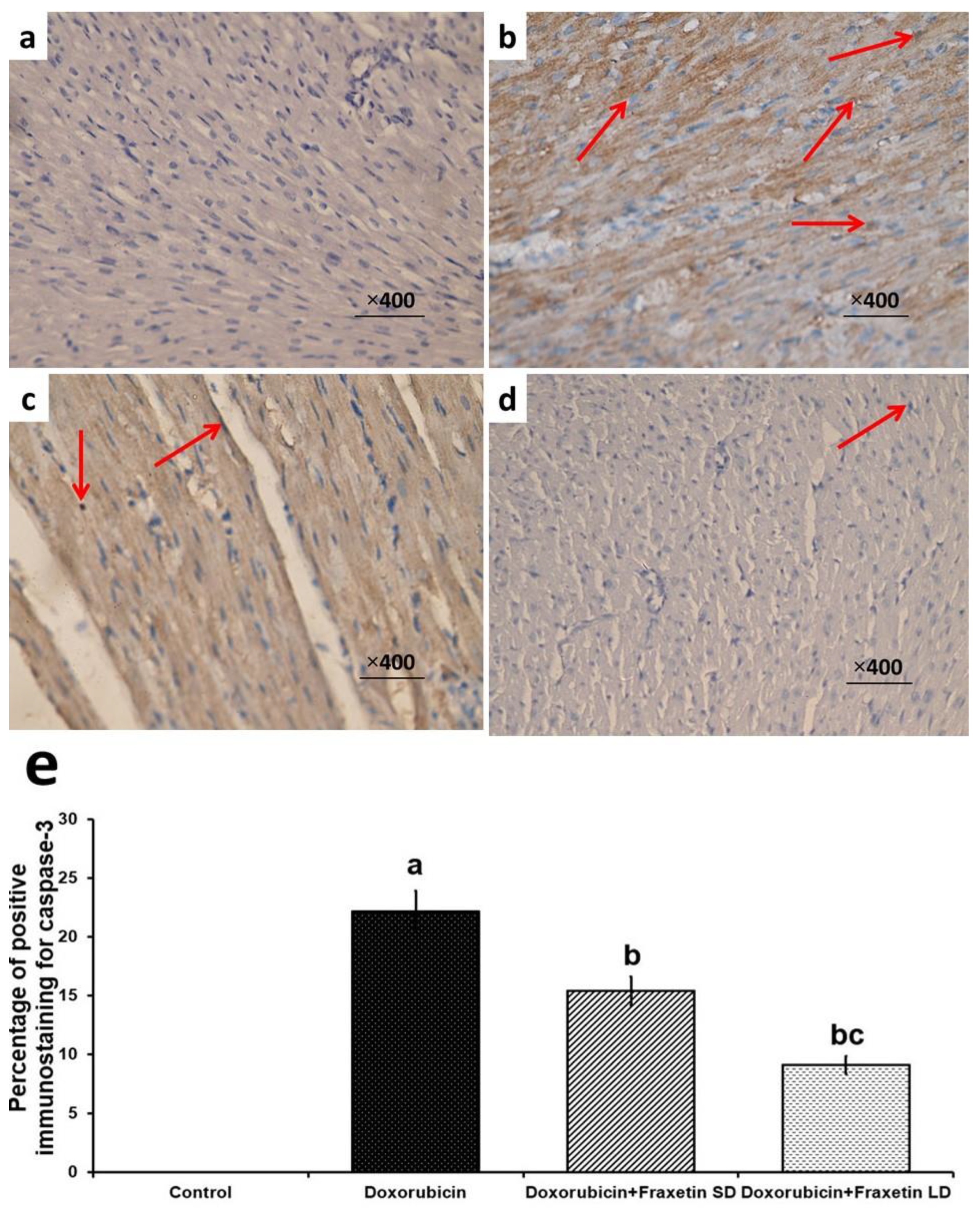
3.5. Fraxetin Abrogated the Apoptotic Changes in Cardiac Tissues of Doxorubicin-Treated Rats
3.6. Effect of Doxorubicin with or without Fraxetin on Echocardiographic Indices in the Studied Groups
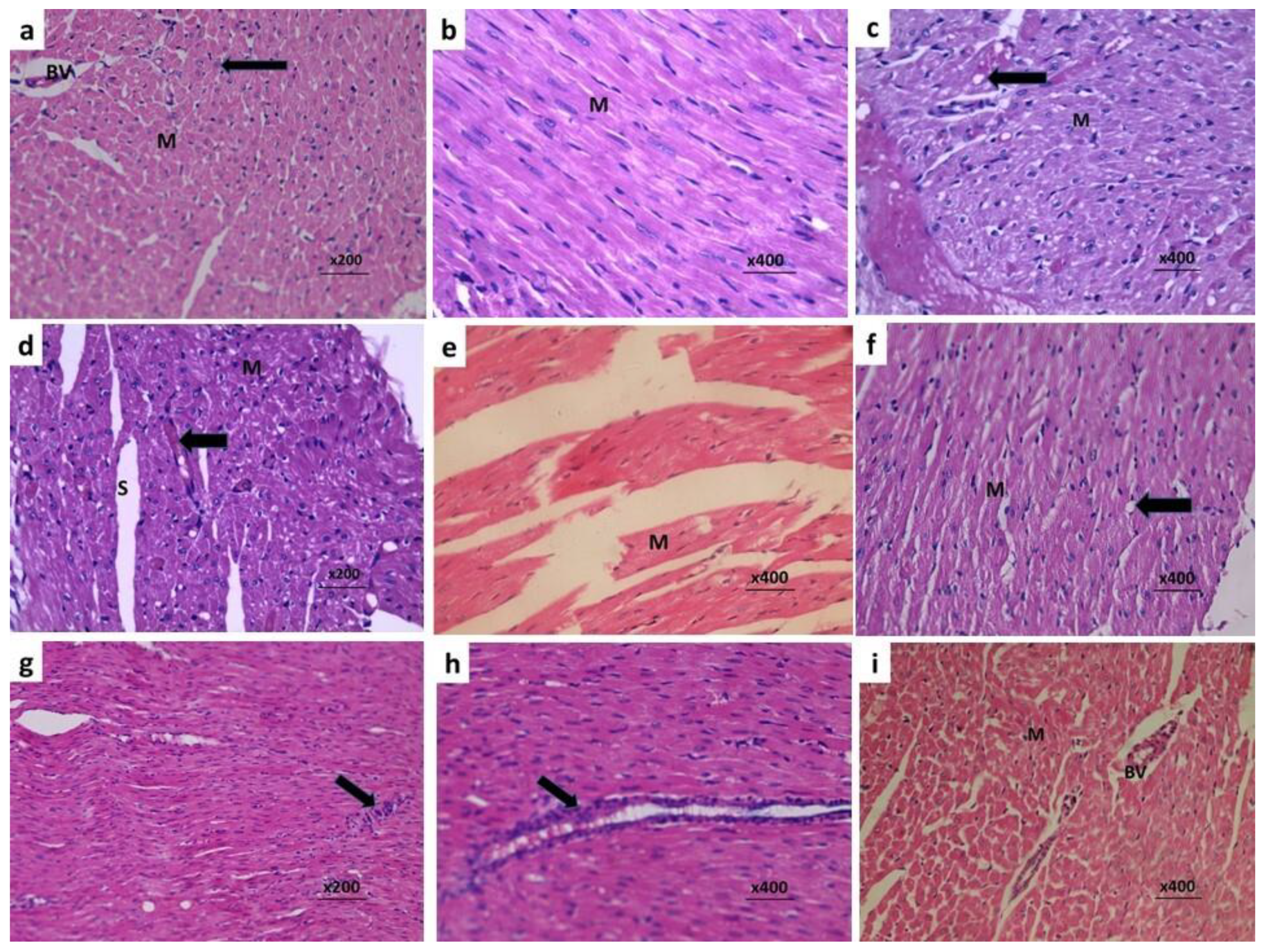
3.7. Fraxetin Combatted the Histopathological Changes Induced by Doxorubicin in Cardiac Tissues
3.8. Fraxetin Abrogated the Electron Microscopic Changes Induced by Doxorubicin in Cardiac Tissues
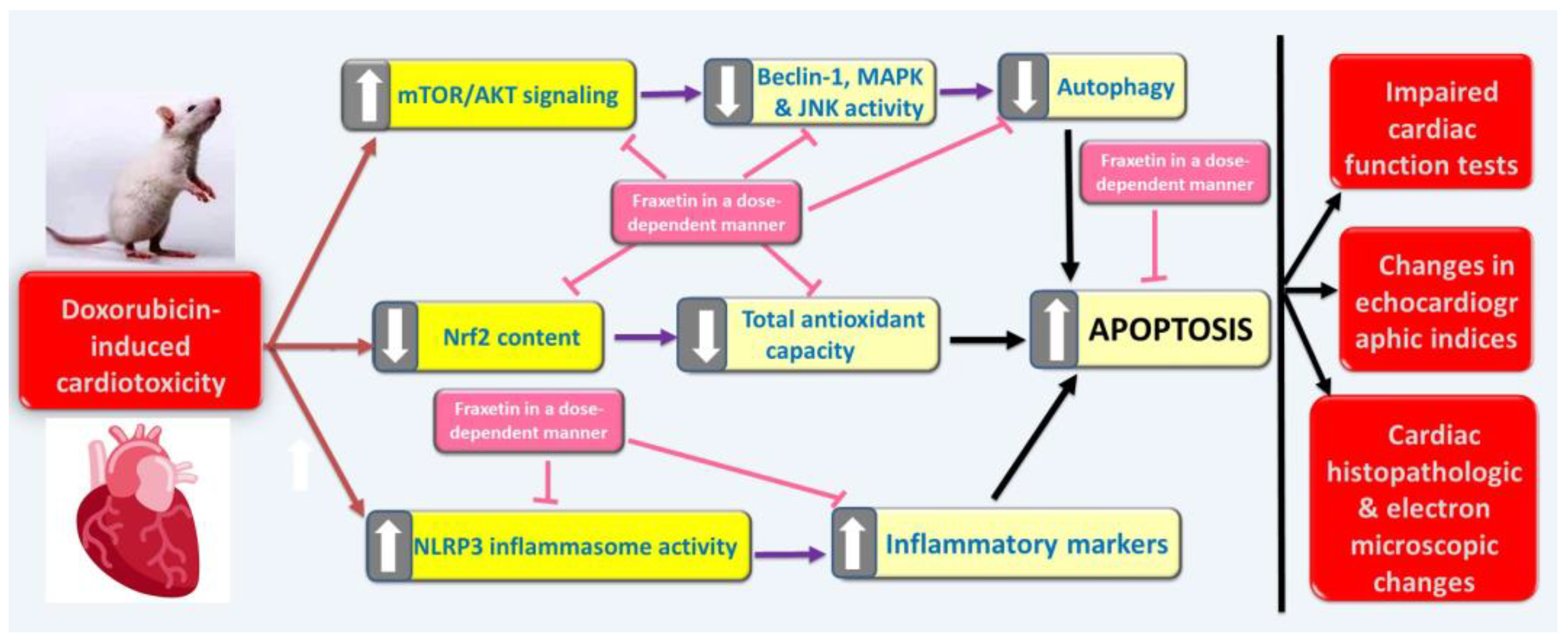
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability statement
Acknowledgments
Conflicts of Interest
References
- Hevener, K.; Verstak, T.A.; Lutat, K.E.; Riggsbee, D.L.; Mooney, J.W. Recent developments in topoisomerase-targeted cancer chemotherapy. Acta. Pharm. Sin. B 2018, 8, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, O.; Shapira, M.; Hershko, D.D. Differential effects of doxorubicin treatment on cell cycle arrest and Skp2 expression in breast cancer cells. Anticancer Drugs 2007, 18, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M.; Alzahrani, A.A.; Bawazir, N.M.; Khawtani, R.O.; Arab, H.H. Targeting the proinflammatory cytokines, oxidative stress, apoptosis and TGF-β1/STAT-3 signaling by irbesartan to ameliorate doxorubicin-induced hepatotoxicity. J. Infect. Chemother. 2018, 24, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, Z.R.; Tanaka, M.; Pampo, C.; Lepler, S.; Rice, L.; Guingab-Cagmat, J.; Garrett, T.J.; Siemann, D.W. Normal tissue and tumor microenvironment adaptations to aerobic exercise enhance doxorubicin anti-tumor efficacy and ameliorate its cardiotoxicity in retired breeder mice. Oncotarget 2021, 12, 1737–1748. [Google Scholar] [CrossRef]
- Wang, C.; Hu, L.; Guo, S.; Yao, Q.; Liu, X.; Zhang, B.; Meng, X.; Yang, X. Phosphocreatine attenuates doxorubicin-induced cardiotoxicity by inhibiting oxidative stress and activating TAK1 to promote myocardial survival in vivo and in vitro. Toxicology 2021, 460, 152881. [Google Scholar] [CrossRef]
- Yao, L.; Gui, M.; Li, J.; Lu, B.; Wang, J.; Zhou, X.; Fu, D. Shengxian decoction decreases doxorubicin-induced cardiac apoptosis by regulating the TREM1/NF-κB signaling pathway. Mol. Med. Rep. 2021, 23, 219. [Google Scholar] [CrossRef]
- Vasti, C.; Hertig, C.M. Neuregulin-1/erbB activities with focus on the susceptibility of the heart to anthracyclines. World J. Cardiol. 2014, 6, 653–662. [Google Scholar] [CrossRef]
- Maayah, Z.H.; Takahara, S.; Dyck, J.R.B. The beneficial effects of reducing NLRP3 inflammasome activation in the cardiotoxicity and the anti-cancer effects of doxorubicin. Arch. Toxicol. 2021, 95, 1–9. [Google Scholar] [CrossRef]
- Sun, Z.; Lu, W.; Lin, N.; Lin, H.; Zhang, J.; Ni, T.; Meng, L.; Zhang, C.; Guo, H. Dihydromyricetin alleviates doxorubicin-induced cardiotoxicity by inhibiting NLRP3 inflammasome through activation of SIRT1. Biochem. Pharmacol. 2020, 175, 113888. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Shi, H.; Yu, Y.; Yu, Y.; Li, M.; Chen, R. NLRP3 inflammasome, an immune-inflammatory target in pathogenesis and treatment of cardiovascular diseases. Clin. Transl. Med. 2020, 10, 91–106. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Sayed, A.M.; Hussein, O.E.; Mahmoud, A.M. Coumarins as Modulators of the Keap1/Nrf2/ARE Signaling Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 1675957. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Xiao, Y.; Guo, H.; Zhu, H.; Chen, D.; Wang, J.; Deng, J.; Lan, J.; Liu, X.; Zhang, Q.; et al. The anti-dysenteric drug fraxetin enhances anti-tumor efficacy of gemcitabine and suppresses pancreatic cancer development by antagonizing STAT3 activation. Aging 2021, 13, 18545–18563. [Google Scholar] [CrossRef]
- González-Cabo, P.; Vázquez-Manrique, R.P.; García-Gimeno, M.A.; Sanz, P.; Palau, F. Frataxin interacts functionally with mitochondrial electron transport chain proteins. Hum. Mol. Genet. 2005, 14, 2091–2098. [Google Scholar] [CrossRef]
- Tsai, H.H.; Rodríguez-Celma, J.; Lan, P.; Wu, Y.C.; Vélez-Bermúdez, I.C.; Schmidt, W. Scopoletin 8-Hydroxylase-Mediated Fraxetin Production Is Crucial for Iron Mobilization. Plant Physiol. 2018, 177, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Erdei, J.; Tóth, A.; Balogh, E.; Nyakundi, B.B.; Bányai, E.; Ryffel, B.; Paragh, G.; Cordero, M.D.; Jeney, V. Induction of NLRP3 Inflammasome Activation by Heme in Human Endothelial Cells. Oxid. Med. Cell. Longev. 2018, 2018, 4310816. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, M.; He, X.; Ouyang, D. A mini-review on ion fluxes that regulate NLRP3 inflammasome activation. Acta. Biochim. Biophys. Sin. 2021, 53, 131–139. [Google Scholar] [CrossRef]
- Argun, M.; Üzüm, K.; Sönmez, M.F.; Özyurt, A.; Derya, K.; Çilenk, K.T.; Unalmış, S.; Pamukcu, Ö.; Baykan, A.; Narin, F.; et al. Cardioprotective effect of metformin against doxorubicin cardiotoxicity in rats. Anatol. J. Cardiol. 2016, 16, 234–241. [Google Scholar] [CrossRef]
- Ekinci-Akdemi, R.F.N.; Bi Ngöl, Ç.; Yıldırım, S.; Kandemi, R.F.M.; Küçükler, S.; Sağlam, Y.S. The investigation of the effect of fraxin on hepatotoxicity induced by cisplatin in rats. Iran. J. Basic. Med. Sci. 2020, 23, 1382–1387. [Google Scholar] [CrossRef]
- Murali, R.; Srinivasan, S.; Ashokkumar, N. Antihyperglycemic effect of fraxetin on hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Biochimie 2013, 95, 1848–1854. [Google Scholar] [CrossRef]
- Laferriere, C.A.; Pang, D.S. Review of Intraperitoneal Injection of Sodium Pentobarbital as a Method of Euthanasia in Laboratory Rodents. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 254–263. [Google Scholar] [CrossRef]
- Walker, J.R.; Sharma, A.; Lytwyn, M.; Bohonis, S.; Thliveris, J.; Singal, P.K.; Jassal, D.S. The cardioprotective role of probucol against anthracycline and trastuzumab-mediated cardiotoxicity. J. Am. Soc. Echocardiogr. 2011, 24, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Marinello, J.; Delcuratolo, M.; Capranico, G. Anthracyclines as Topoisomerase II Poisons: From Early Studies to New Perspectives. Int. J. Mol. Sci. 2018, 19, 3480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabel, A.M.; Omar, M.S.; Balaha, M.F.; Borg, H.M. Effect of metformin and adriamycin on transplantable tumor model. Tissue Cell. 2015, 47, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, F.; He, B.; Fang, C. Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage. Open Life Sci. 2021, 16, 856–866. [Google Scholar] [CrossRef]
- Arunachalam, S.; Nagoor Meeran, M.F.; Azimullah, S.; Sharma, C.; Goyal, S.N.; Ojha, S. Nerolidol Attenuates Oxidative Stress, Inflammation, and Apoptosis by Modulating Nrf2/MAPK Signaling Pathways in Doxorubicin-Induced Acute Cardiotoxicity in Rats. Antioxidants 2021, 10, 984. [Google Scholar] [CrossRef]
- Gu, J.; Huang, H.; Liu, C.; Jiang, B.; Li, M.; Liu, L.; Zhang, S. Pinocembrin inhibited cardiomyocyte pyroptosis against doxorubicin-induced cardiac dysfunction via regulating Nrf2/Sirt3 signaling pathway. Int. Immunopharmacol. 2021, 95, 107533. [Google Scholar] [CrossRef]
- El-Agamy, D.S.; El-Harbi, K.M.; Khoshhal, S.; Ahmed, N.; Elkablawy, M.A.; Shaaban, A.A.; Abo-Haded, H.M. Pristimerin protects against doxorubicin-induced cardiotoxicity and fibrosis through modulation of Nrf2 and MAPK/NF-kB signaling pathways. Cancer Manag. Res. 2018, 11, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Liu, D.; Xing, R.; Song, H.; Tian, X.; Yan, C.; Han, Y. Orosomucoid 1 Attenuates Doxorubicin-Induced Oxidative Stress and Apoptosis in Cardiomyocytes via Nrf2 Signaling. Biomed. Res. Int. 2020, 2020, 5923572. [Google Scholar] [CrossRef]
- Zhang, W.B.; Lai, X.; Guo, X.F. Activation of Nrf2 by miR-152 Inhibits Doxorubicin-Induced Cardiotoxicity via Attenuation of Oxidative Stress, Inflammation, and Apoptosis. Oxid. Med. Cell. Longev. 2021, 2021, 8860883. [Google Scholar] [CrossRef]
- Najmanová, I.; Doseděl, M.; Hrdina, R.; Anzenbacher, P.; Filipský, T.; Říha, M.; Mladěnka, P. Cardiovascular effects of coumarins besides their antioxidant activity. Curr. Top. Med. Chem. 2015, 15, 830–849. [Google Scholar] [CrossRef]
- Kundu, J.; Chae, I.G.; Chun, K.S. Fraxetin Induces Heme Oxygenase-1 Expression by Activation of Akt/Nrf2 or AMP-activated Protein Kinase α/Nrf2 Pathway in HaCaT Cells. J. Cancer Prev. 2016, 21, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K.; Cheema, H.S.; Saxena, A.; Jyotshana Singh, S.; Darokar, M.P.; Bawankule, D.U.; Shanker, K.; Luqman, S. Fraxetin and ethyl acetate extract from Lawsonia inermis L. ameliorate oxidative stress in P. berghei infected mice by augmenting antioxidant defence system. Phytomedicine 2017, 36, 262–272. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Alyaseer, A.A.A.; de Lima, M.H.S.; Braga, T.T. The Role of NLRP3 Inflammasome Activation in the Epithelial to Mesenchymal Transition Process during the Fibrosis. Front. Immunol. 2020, 11, 883. [Google Scholar] [CrossRef]
- Wei, S.; Ma, W.; Li, X.; Jiang, C.; Sun, T.; Li, Y.; Zhang, B.; Li, W. Involvement of ROS/NLRP3 Inflammasome Signaling Pathway in Doxorubicin-Induced Cardiotoxicity. Cardiovasc. Toxicol. 2020, 20, 507–519. [Google Scholar] [CrossRef]
- Chen, G.; Song, X.; Lin, D.; Xu, P. Isofraxidin Alleviates Myocardial Infarction through NLRP3 Inflammasome Inhibition. Inflammation 2020, 43, 712–721. [Google Scholar] [CrossRef]
- Di Stasi, L.C. Coumarin Derivatives in Inflammatory Bowel Disease. Molecules 2021, 26, 422. [Google Scholar] [CrossRef]
- Wu, B.; Wang, R.; Li, S.; Wang, Y.; Song, F.; Gu, Y.; Yuan, Y. Antifibrotic effects of Fraxetin on carbon tetrachloride-induced liver fibrosis by targeting NF-κB/IκBα, MAPKs and Bcl-2/Bax pathways. Pharmacol. Rep. 2019, 71, 409–416. [Google Scholar] [CrossRef]
- Miao, Y.; Yang, J.; Yun, Y.; Sun, J.; Wang, X. Synthesis and anti-rheumatoid arthritis activities of 3-(4-aminophenyl)-coumarin derivatives. J. Enzyme. Inhib. Med. Chem. 2021, 36, 450–461. [Google Scholar] [CrossRef]
- Mishra, S.; Pandey, A.; Manvati, S. Coumarin: An emerging antiviral agent. Heliyon 2020, 6, e03217. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.H.; Hung, T.W.; Chen, Y.S.; Huang, Y.N.; Chiou, H.L.; Lee, C.C.; Tsai, J.P. In Vitro and In Vivo Antifibrotic Effects of Fraxetin on Renal Interstitial Fibrosis via the ERK Signaling Pathway. Toxins 2021, 13, 474. [Google Scholar] [CrossRef]
- Xiao, B.; Hong, L.; Cai, X.; Mei, S.; Zhang, P.; Shao, L. The true colors of autophagy in doxorubicin-induced cardiotoxicity. Oncol. Lett. 2019, 18, 2165–2172. [Google Scholar] [CrossRef] [Green Version]
- Christidi, E.; Brunham, L.R. Regulated cell death pathways in doxorubicin-induced cardiotoxicity. Cell Death Dis. 2021, 12, 339. [Google Scholar] [CrossRef]
- Liao, D.; Zhang, C.; Liu, N.; Cao, L.; Wang, C.; Feng, Q.; Yao, D.; Long, M.; Jiang, P. Involvement of neurotrophic signaling in doxorubicin-induced cardiotoxicity. Exp. Ther. Med. 2020, 19, 1129–1135. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, C.; Liu, C.; Wei, F. Luteolin attenuates doxorubicin-induced cardiotoxicity by modulating the PHLPP1/AKT/Bcl-2 signalling pathway. PeerJ 2020, 8, e8845. [Google Scholar] [CrossRef]
- Sánchez-Reus, M.I.; Peinado, I.I.; Molina-Jiménez, M.F.; Benedí, J. Fraxetin prevents rotenone-induced apoptosis by induction of endogenous glutathione in human neuroblastoma cells. Neurosci. Res. 2005, 53, 48–56. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, J.; Wang, Q.; Li, Y.; Zhang, B. Fraxetin inhibits the proliferation of RL95-2 cells through regulation of metabolism. Int. J. Clin. Exp. Pathol. 2020, 13, 1500–1505. [Google Scholar]
- Sumorek-Wiadro, J.; Zając, A.; Langner, E.; Skalicka-Woźniak, K.; Maciejczyk, A.; Rzeski, W.; Jakubowicz-Gil, J. Antiglioma Potential of Coumarins Combined with Sorafenib. Molecules 2020, 25, 5192. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Doxorubicin | Doxorubicin + Fraxetin Small Dose | Doxorubicin + Fraxetin Large Dose |
|---|---|---|---|---|
| Serum CK-MB (U/L) | 75.21 ± 8.13 | 169.6 ± 18.23 a | 130.15 ± 14.38 b | 105.02 ± 11.7 bc |
| Serum LDH (U/L) | 742.3 ± 80.6 | 1994.6 ± 180.9 a | 1412.8 ± 148.5 b | 968.8±101.2 bc |
| Serum troponin I (ng/mL) | 1.73 ± 0.18 | 3.87 ± 0.41 a | 2.72±0.32 b | 2.18 ±0.24 bc |
| Serum NT-pro BNP (ng/L) | 8.68 ± 1.06 | 18.23 ± 1.95 a | 13.92 ± 1.53 b | 10.27 ± 1.21 bc |
| Parameters | Control | Doxorubicin | Doxorubicin + Fraxetin Small Dose | Doxorubicin + Fraxetin Large Dose |
|---|---|---|---|---|
| Tissue p-mTOR (% change from the control) | 100.00 ± 10.95 | 191.72 ± 20.22 a | 161.1 ± 18.34 b | 130.84 ± 15.18 bc |
| Tissue p-AKT (% change from the control) | 100.00 ± 9.89 | 183.49 ± 19.8 a | 155.95 ± 17.47 b | 124.17 ± 14.4 bc |
| Tissue p38 MAPK (% change from the control) | 100.00 ± 11.45 | 56.17 ± 6.91 a | 73.84 ± 8.25 b | 86.38 ± 9.16 bc |
| Tissue JNK activity (% change of control) | 100.0 ± 12.3 | 54.3 ± 5.8 a | 70.2 ± 7.32 b | 81.52 ± 8.82 bc |
| Tissue beclin-1 (ng/g protein) | 8.36 ± 0.94 | 3.89 ± 0.45 a | 5.75 ± 0.61 b | 6.97 ± 0.82 bc |
| Parameters | Control | Doxorubicin | Doxorubicin + Fraxetin Small Dose | Doxorubicin + Fraxetin Large Dose |
|---|---|---|---|---|
| Heart Rate (bpm) | 229 ± 24 | 225 ± 23 | 234 ± 26 | 228 ± 24 |
| LVEDD (cm) | 0.59 ± 0.07 | 0.57±0.06 | 0.56 ± 0.06 | 0.61 ± 0.07 |
| LVESD (cm) | 0.32 ± 0.04 | 0.45 ± 0.05 a | 0.38 ± 0.04 b | 0.34 ± 0.04 bc |
| LVEF (%) | 78.4 ± 8.92 | 39.4 ± 4.21 a | 55.4 ± 5.81 b | 68.9 ± 7.12 bc |
| LVFS (%) | 50.3 ± 6.1 | 24.76 ± 2.6 a | 34.5 ± 3.6 b | 41.15 ± 4.41 bc |
| MPI | 0.38 ± 0.04 | 0.21 ± 0.03 a | 0.27 ± 0.03 b | 0.34 ± 0.04 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabel, A.M.; Salama, S.A.; Adwas, A.A.; Estfanous, R.S. Targeting Oxidative Stress, NLRP3 Inflammasome, and Autophagy by Fraxetin to Combat Doxorubicin-Induced Cardiotoxicity. Pharmaceuticals 2021, 14, 1188. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111188
Kabel AM, Salama SA, Adwas AA, Estfanous RS. Targeting Oxidative Stress, NLRP3 Inflammasome, and Autophagy by Fraxetin to Combat Doxorubicin-Induced Cardiotoxicity. Pharmaceuticals. 2021; 14(11):1188. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111188
Chicago/Turabian StyleKabel, Ahmed M., Samir A. Salama, Almokhtar A. Adwas, and Remon S. Estfanous. 2021. "Targeting Oxidative Stress, NLRP3 Inflammasome, and Autophagy by Fraxetin to Combat Doxorubicin-Induced Cardiotoxicity" Pharmaceuticals 14, no. 11: 1188. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14111188