
Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Results
2.1. Phytochemicals Analysis
2.1.1. Quantitative Analysis of Extracts’ Components
2.1.2. Total Polyphenols Content
2.2. Biological Activity
2.2.1. Anti-Tumor Activity
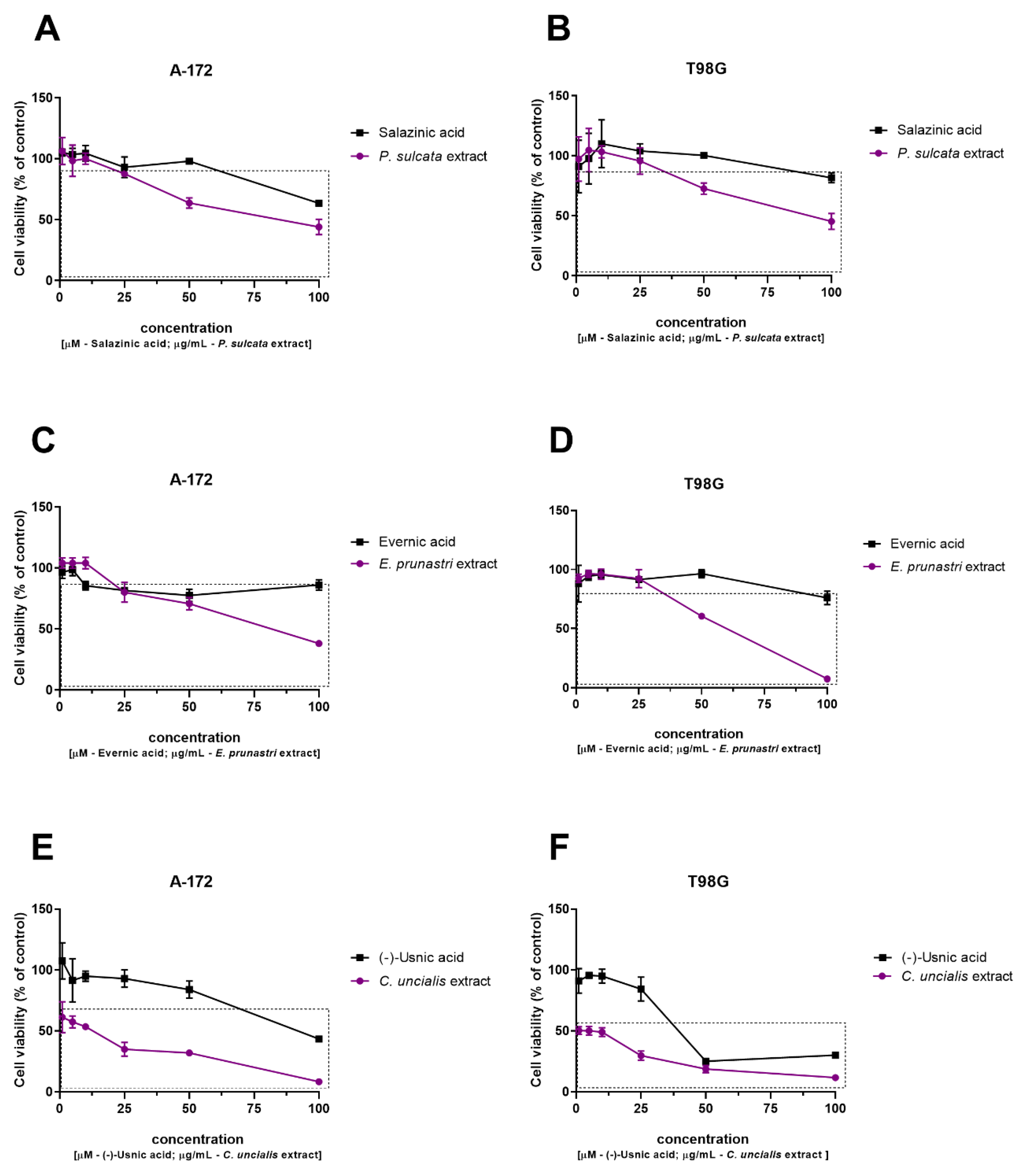
Cytotoxic Activity against GBM Cells
Inhibition of the Kynurenine Pathway Enzymes: Indoleamine 2,3-Dioxygenases 1 (IDO1), Indoleamine 2,3-Dioxygenases 2 (IDO2), and Tryptophan 2,3-Dioxygenase (TDO)
2.2.2. Anti-Inflammatory Activity
Inhibition of Cyclooxygenase-2 (COX-2)
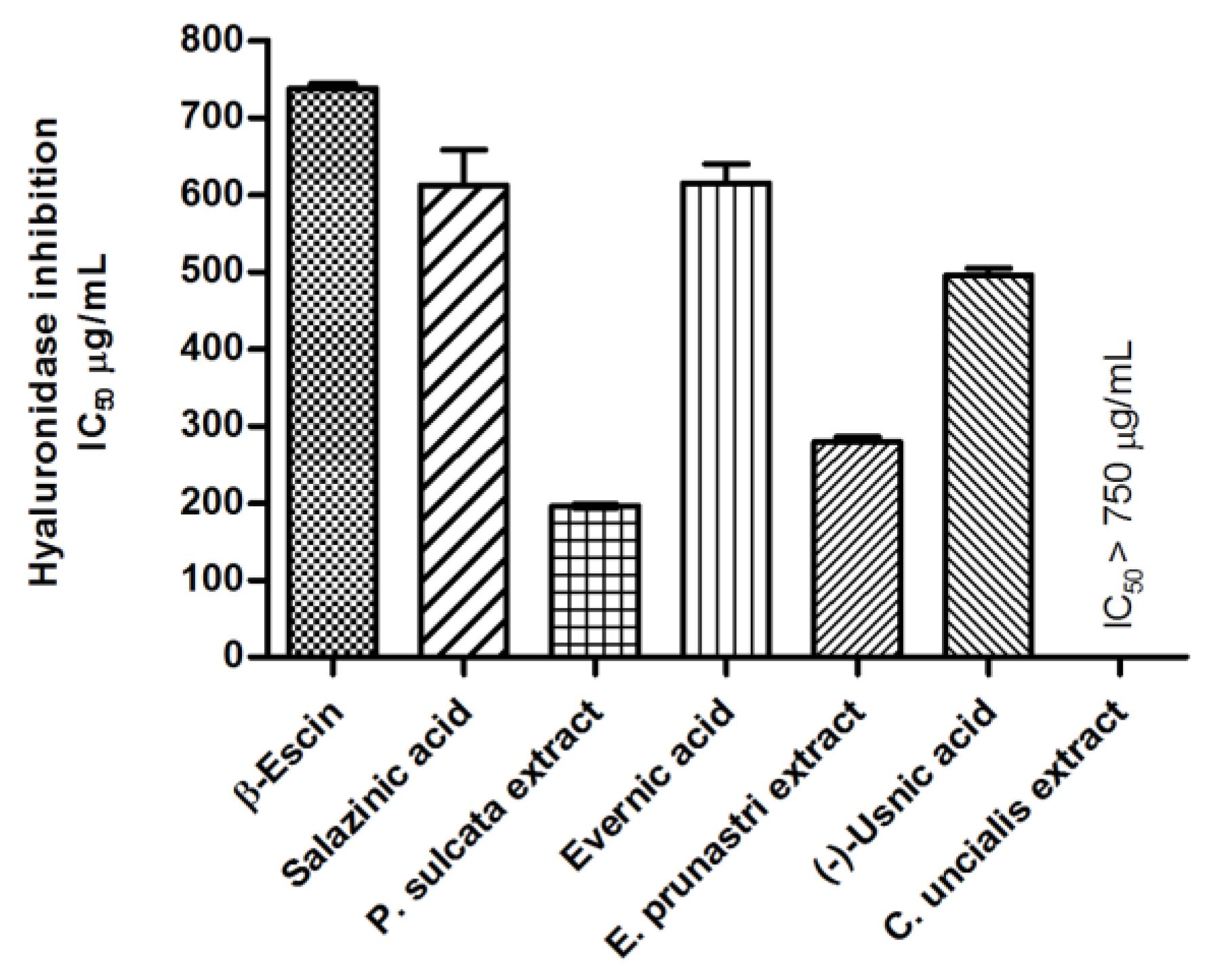
Anti-Hyaluronidase Activity
2.2.3. Impact on Reactive Oxygen Species (ROS) Homeostasis
Effect on Antioxidant Enzyme Activity
2.2.4. Anticholinergic Activity
2.3. Permeability through the Blood-Brain-Barrier (PAMPA-BBB)
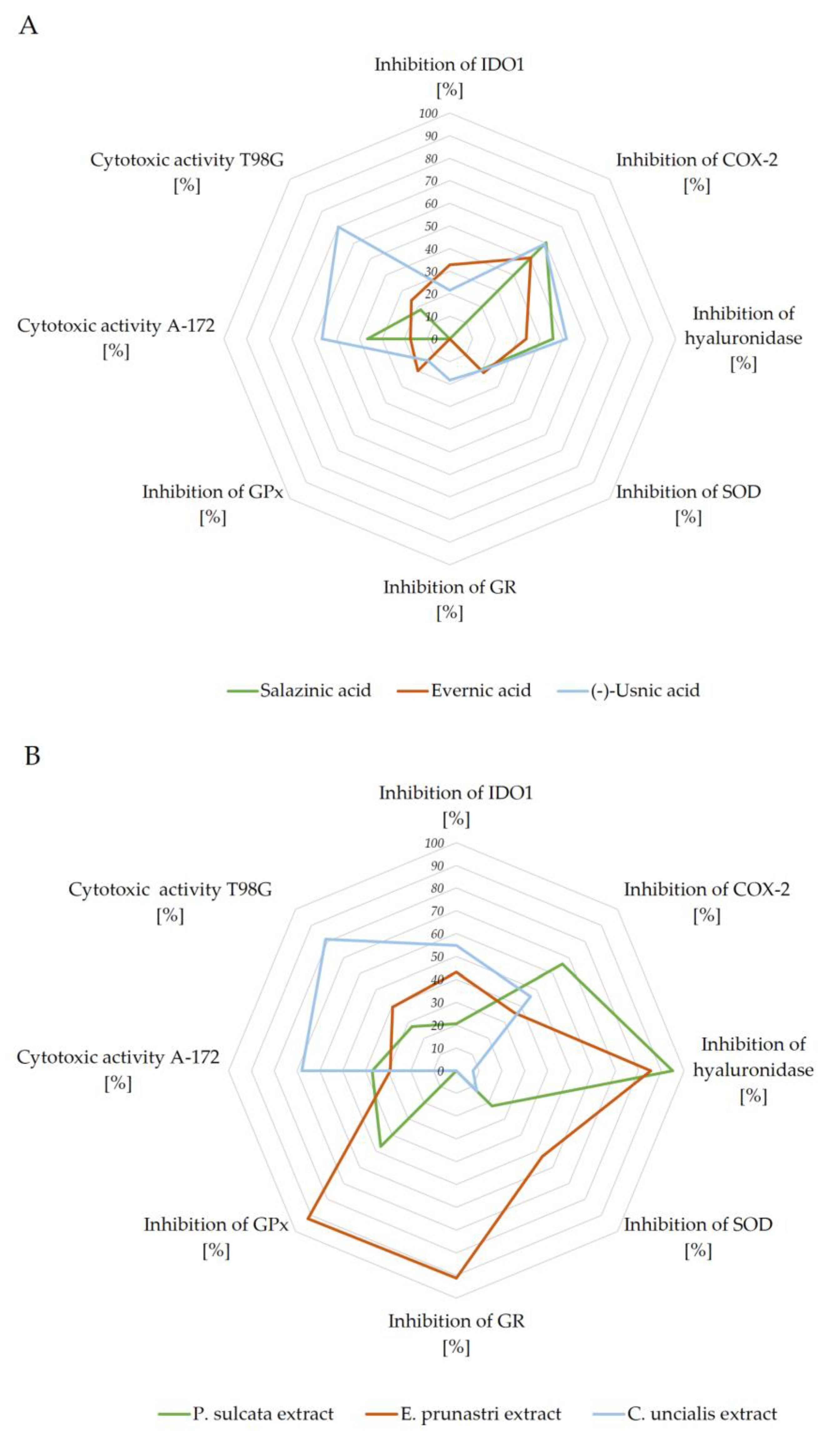
2.4. Summary of Biological Potential of Lichen-Derived Compounds and Extracts
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Solvents and Chemicals
4.3. Preparation of Extract
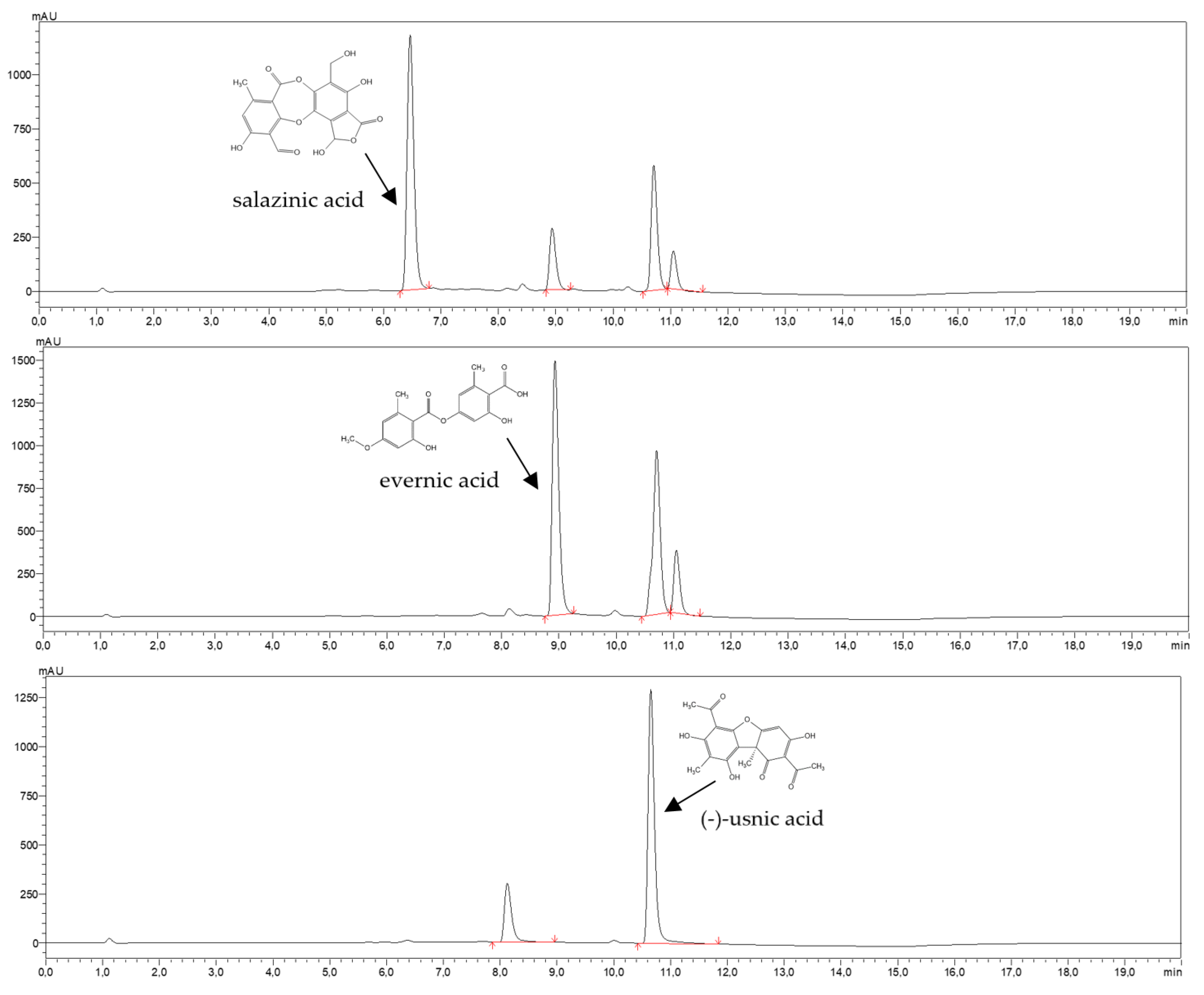
4.4. HPLC Analysis
4.5. Total Phenolic Content (TPC)
4.6. Determination of Cytotoxicity of Lichen-Derived Substances
4.6.1. Compounds/Extracts
4.6.2. Cell Culture and Assessment of Cell Viability
4.7. Inhibition of Indoleamine 2,3-Dioxygenase (IDO1)
4.8. Effect on Cyclooxygenase-2 (COX-2) Activity
4.9. Anti-Hyaluronidase Activity
4.10. Antioxidant Activity
4.10.1. DPPH and CUPRAC analysis
4.10.2. Effect on Antioxidant Enzymes Activity
Effect on Superoxide Dismutase Activity (SOD)
Effect on Glutathione Reductase (GR) Activity
Effect on Glutathione Peroxidase (GPx) Activity
4.11. Anti-Cholinesterase Activity
4.12. Permeability through the Blood-Brain-Barrier (PAMPA-BBB)
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Majchrzak-Celińska, A.; Kleszcz, R.; Stasiłowicz-Krzemień, A.; Cielecka-Piontek, J. Sodium Butyrate Enhances Curcuminoids Permeability through the Blood-Brain Barrier, Restores Wnt/β-Catenin Pathway Antagonists Gene Expression and Reduces the Viability of Glioblastoma Cells. Int. J. Mol. Sci. 2021, 22, 11285. [Google Scholar] [CrossRef] [PubMed]
- Solárová, Z.; Liskova, A.; Samec, M.; Kubatka, P.; Büsselberg, D.; Solár, P. Anticancer potential of lichens’ secondary metabolites. Biomolecules 2020, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, M.; Xu, B. A comprehensive review on secondary metabolites and health-promoting effects of edible lichen. J. Funct. Foods 2021, 80, 104283. [Google Scholar] [CrossRef]
- Purvis, O.W.; Coppins, B.J.; Hawksworth, D.L.; James, P.W. The Lichen Flora of Great Britain and Irleande; Natural History Museum: London, UK, 2019. [Google Scholar]
- Stojanovic, G.; Stojanovic, I.; Smelcerovic, A. Lichen depsidones as potential novel pharmacologically active compounds. Mini. Rev. Org. Chem. 2012, 9, 178–184. [Google Scholar] [CrossRef]
- Maslać, A.; Maslać, M.; Tkalec, M. The impact of cadmium on photosynthetic performance and secondary metabolites in the lichens Parmelia sulcata, Flavoparmelia caperata and Evernia prunastri. Acta Bot. Croat. 2016, 75, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Alexandrino, C.A.F.; Honda, N.K.; Matos, M.d.F.C.; Portugal, L.C.; Souza, P.R.B.d.; Perdomo, R.T.; Guimarães, R.d.C.A.; Kadri, M.C.T.; Silva, M.C.B.L.; Bogo, D. Antitumor effect of depsidones from lichens on tumor cell lines and experimental murine melanoma. Rev. Bras. Farmacogn. 2019, 29, 449–456. [Google Scholar] [CrossRef]
- Ari, F.; Ulukaya, E.; Oran, S.; Celikler, S.; Ozturk, S.; Ozel, M.Z. Promising anticancer activity of a lichen, Parmelia sulcata Taylor, against breast cancer cell lines and genotoxic effect on human lymphocytes. Cytotechnology 2015, 67, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Kosanić, M.; Manojlović, N.; Janković, S.; Stanojković, T.; Ranković, B. Evernia prunastri and Pseudoevernia furfuraceae lichens and their major metabolites as antioxidant, antimicrobial and anticancer agents. Food Chem. Toxicol. 2013, 53, 112–118. [Google Scholar] [CrossRef]
- Manojlović, N.; Ranković, B.; Kosanić, M.; Vasiljević, P.; Stanojković, T. Chemical composition of three Parmelia lichens and antioxidant, antimicrobial and cytotoxic activities of some their major metabolites. Phytomedicine 2012, 19, 1166–1172. [Google Scholar] [CrossRef]
- Verma, N.; Behera, B.C.; Joshi, A. Studies on nutritional requirement for the culture of lichen Ramalina nervulosa and Ramalina pacifica to enhance the production of antioxidant metabolites. Folia Microbiol. (Praha). 2012, 57, 107–114. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Vukojević, J. Antioxidant properties of some lichen species. J. Food Sci. Technol. 2011, 48, 584–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Suh, Y.J.; Yang, S.; Hong, D.G.; Ishigami, A.; Kim, H.; Hur, J.-S.; Chang, S.-C.; Lee, J. Neuroprotective and Anti-Inflammatory Effects of Evernic Acid in an MPTP-Induced Parkinson’s Disease Model. Int. J. Mol. Sci. 2021, 22, 2098. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moriano, C.; Divakar, P.K.; Crespo, A.; Gómez-Serranillos, M.P. Protective effects of lichen metabolites evernic and usnic acids against redox impairment-mediated cytotoxicity in central nervous system-like cells. Food Chem. Toxicol. 2017, 105, 262–277. [Google Scholar] [CrossRef]
- Galanty, A.; Paśko, P.; Podolak, I. Enantioselective activity of usnic acid: A comprehensive review and future perspectives. Phytochem. Rev. 2019, 18, 527–548. [Google Scholar] [CrossRef] [Green Version]
- Rabelo, T.K.; Zeidán-Chuliá, F.; Vasques, L.M.; dos Santos, J.P.A.; da Rocha, R.F.; de Bittencourt Pasquali, M.A.; Rybarczyk-Filho, J.L.; Araújo, A.A.S.; Moreira, J.C.F.; Gelain, D.P. Redox characterization of usnic acid and its cytotoxic effect on human neuron-like cells (SH-SY5Y). Toxicol. Vitr. 2012, 26, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Emsen, B.; Aslan, A.; Türkez, H.; Joughi, A.T.; Kaya, A. The anti-cancer efficacies of diffractaic, lobaric, and usnic acid: In vitro inhibition of glioma. J. Cancer Res Ther. 2018, 14, 941–951. [Google Scholar] [CrossRef]
- Koparal, A.T. Anti-angiogenic and antiproliferative properties of the lichen substances (−)-usnic acid and vulpinic acid. Zeitschrift für Naturforsch. C 2015, 70, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jo, S.; Lee, H.; Kim, T.U.; Kim, I.-C.; Yim, J.H.; Chung, H. Lobarstin enhances chemosensitivity in human glioblastoma T98G cells. Anticancer Res. 2013, 33, 5445–5451. [Google Scholar] [PubMed]
- Guzow-Krzemińska, B.; Guzow, K.; Herman-Antosiewicz, A. Usnic Acid Derivatives as Cytotoxic Agents Against Cancer Cells and the Mechanisms of Their Activity. Curr. Pharmacol. Reports 2019, 5, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Ingelfinger, R.; Henke, M.; Roser, L.; Ulshöfer, T.; Calchera, A.; Singh, G.; Parnham, M.J.; Geisslinger, G.; Fürst, R.; Schmitt, I.; et al. Unraveling the Pharmacological Potential of Lichen Extracts in the Context of Cancer and Inflammation with a Broad Screening Approach. Front. Pharmacol. 2020, 11, 1322. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Hołderna-Kędzia, E.; Galanty, A.; Bylka, W.; Kacprzak, K.; Ćwiklińska, K. In vitro antimicrobial activity of extracts and compounds isolated from Cladonia uncialis. Nat. Prod. Res. 2015, 29, 2302–2307. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Żarowski, M.; Plech, T.; Cielecka-Piontek, J. Permeability of Hypogymnia physodes Extract Component—Physodic Acid through the Blood–Brain Barrier as an Important Argument for Its Anticancer and Neuroprotective Activity within the Central Nervous System. Cancers 2021, 13, 1717. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, P.P.; Sordillo, L.A.; Helson, L. The Kynurenine Pathway: A Primary Resistance Mechanism in Patients with Glioblastoma. Anticancer. Res. 2017, 37, 2159–2171. [Google Scholar] [CrossRef] [Green Version]
- Teismann, P. COX-2 in the neurodegenerative process of Parkinson’s disease. Biofactors 2012, 38, 395–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majchrzak-Celińska, A.; Misiorek, J.O.; Kruhlenia, N.; Przybyl, L.; Kleszcz, R.; Rolle, K.; Krajka-Kuźniak, V. COXIBs and 2,5-dimethylcelecoxib counteract the hyperactivated Wnt/β-catenin pathway and COX-2/PGE2/EP4 signaling in glioblastoma cells. BMC Cancer 2021, 21, 493. [Google Scholar] [CrossRef] [PubMed]
- Strauss, K.I. Antiinflammatory and neuroprotective actions of COX2 inhibitors in the injured brain. Brain. Behav. Immun. 2008, 22, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Bralley, E.; Greenspan, P.; Hargrove, J.L.; Hartle, D.K. Inhibition of hyaluronidase activity by Vitis rotundifolia.(Muscadine) berry seeds and skins. Pharm. Biol. 2007, 45, 667–673. [Google Scholar] [CrossRef]
- Diao, S.; Xiao, M.; Chen, C. The Role of Hyaluronan in Myelination and Remyelination after White Matter Injury. Brain Res. 2021, 147522. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Kanzaki, H.; Wada, S.; Narimiya, T.; Yamaguchi, Y.; Katsumata, Y.; Itohiya, K.; Fukaya, S.; Miyamoto, Y.; Nakamura, Y. Pathways that Regulate ROS Scavenging Enzymes, and Their Role in Defense Against Tissue Destruction in Periodontitis. Front. Physiol. 2017, 8, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Ayllón, M.S.; Sáez-Valero, J.; Muñoz-Delgado, E.; Vidal, C.J. Identification of hybrid cholinesterase forms consisting of acetyl- and butyrylcholinesterase subunits in human glioma. Neuroscience 2001, 107, 199–208. [Google Scholar] [CrossRef]
- Pezzementi, L.; Nachon, F.; Chatonnet, A. Evolution of acetylcholinesterase and butyrylcholinesterase in the vertebrates: An atypical butyrylcholinesterase from the Medaka Oryzias latipes. PLoS ONE 2011, 6, e17396. [Google Scholar]
- Chen, X.; Murawski, A.; Patel, K.; Crespi, C.L.; Balimane, P. V A novel design of artificial membrane for improving the PAMPA model. Pharm. Res. 2008, 25, 1511–1520. [Google Scholar] [CrossRef]
- Latacz, G.; Lubelska, A.; Jastrzębska-Więsek, M.; Partyka, A.; Marć, M.A.; Satała, G.; Wilczyńska, D.; Kotańska, M.; Więcek, M.; Kamińska, K. The 1, 3, 5-triazine derivatives as innovative chemical family of 5-HT6 serotonin receptor agents with therapeutic perspectives for cognitive impairment. Int. J. Mol. Sci. 2019, 20, 3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studzinska-Sroka, E.; Dubino, A. Lichens as a source of chemical compounds with anti-inflammatory activity. Herba Pol. 2018, 64. [Google Scholar] [CrossRef] [Green Version]
- Aoussar, N.; Rhallabi, N.; Mhand, R.A.; Manzali, R.; Bouksaim, M.; Douira, A.; Mellouki, F. Seasonal variation of antioxidant activity and phenolic content of Pseudevernia furfuracea, Evernia prunastri and Ramalina farinacea from Morocco. J. Saudi Soc. Agric. Sci. 2020, 19, 1–6. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Tomczak, H.; Malińska, N.; Wrońska, M.; Kleszcz, R.; Galanty, A.; Cielecka-Piontek, J.; Latek, D.; Paluszczak, J. Cladonia uncialis as a valuable raw material of biosynthetic compounds against clinical strains of bacteria and fungi. Acta Biochim. Pol. 2019, 66, 597–603. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Brancato, V.; Nuzzo, S.; Tramontano, L.; Condorelli, G.; Salvatore, M.; Cavaliere, C. Predicting Survival in Glioblastoma Patients Using Diffusion MR Imaging Metrics—A Systematic Review. Cancers 2020, 12, 2858. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, A.; Strömstedt, A.A.; Göransson, U.; Gnezdilov, O.; Turanov, A.; Boldbaatar, D.; Kochkin, D.; Ulrich-Merzenich, G.; Koptina, A. Antimicrobial and antioxidant activity of Evernia prunastri extracts and their isolates. World J. Microbiol. Biotechnol. 2021, 37, 129. [Google Scholar] [CrossRef]
- Shcherbakova, A.; Nyugen, L.; Koptina, A.; Backlund, A.; Shurgin, A.; Romanov, E.; Ulrich-Merzenich, G. Screening of compounds of Evernia prunastri (L.) for their antiproliferative activity in glioblastoma cells. Planta Med. 2016, 82, P465. [Google Scholar] [CrossRef]
- Ari, F.; Aztopal, N.; Oran, S.; Bozdemir, S.; Celikler, S.; Ozturk, S.; Ulukaya, E. Parmelia sulcata Taylor and Usnea filipendula Stirt induce apoptosis-like cell death and DNA damage in cancer cells. Cell Prolif. 2014, 47, 457–464. [Google Scholar] [CrossRef] [PubMed]
- White, P.A.S.; Oliveira, R.C.M.; Oliveira, A.P.; Serafini, M.R.; Araújo, A.A.S.; Gelain, D.P.; Moreira, J.C.F.; Almeida, J.R.G.S.; Quintans, J.S.S.; Quintans-Junior, L.J.; et al. Antioxidant activity and mechanisms of action of natural compounds isolated from lichens: A systematic review. Molecules 2014, 19, 14496–14527. [Google Scholar] [CrossRef]
- Lee, S.; Lee, Y.; Ha, S.; Chung, H.Y.; Kim, H.; Hur, J.-S.; Lee, J. Anti-inflammatory effects of usnic acid in an MPTP-induced mouse model of Parkinson’s disease. Brain Res. 2020, 1730, 146642. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Yue, L.; Shi, J.; Shao, M.; Wu, T. Role of IDO and TDO in Cancers and Related Diseases and the Therapeutic Implications. J. Cancer 2019, 10, 2771–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Xing, Z.; Tao, B.; Li, T.; Yang, D.; Li, W.; Zheng, Y.; Kuang, C.; Yang, Q. Both IDO1 and TDO contribute to the malignancy of gliomas via the Kyn-AhR-AQP4 signaling pathway. Signal Transduct. Target. Ther. 2020, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, A.; Tankiewicz-Kwedlo, A.; Pawlak, D. Kynurenines as a Novel Target for the Treatment of Malignancies. Pharmaceuticals 2021, 14, 606. [Google Scholar] [CrossRef]
- Qiu, J.; Shi, Z.; Jiang, J. Cyclooxygenase-2 in glioblastoma multiforme. Drug Discov. Today 2017, 22, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Prayson, R.A.; Castilla, E.A.; Vogelbaum, M.A.; Barnett, G.H. Cyclooxygenase-2 (COX-2) expression by immunohistochemistry in glioblastoma multiforme. Ann. Diagn. Pathol. 2002, 6, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Joki, T.; Heese, O.; Nikas, D.C.; Bello, L.; Zhang, J.; Kraeft, S.-K.; Seyfried, N.T.; Abe, T.; Chen, L.B.; Carroll, R.S. Expression of cyclooxygenase 2 (COX-2) in human glioma and in vitro inhibition by a specific COX-2 inhibitor, NS-398. Cancer Res. 2000, 60, 4926–4931. [Google Scholar]
- Bauer, J.; Waltenberger, B.; Noha, S.M.; Schuster, D.; Rollinger, J.M.; Boustie, J.; Chollet, M.; Stuppner, H.; Werz, O. Discovery of depsides and depsidones from lichen as potent inhibitors of microsomal prostaglandin E2 synthase-1 using pharmacophore models. ChemMedChem 2012, 7, 2077. [Google Scholar] [CrossRef]
- Bugni, T.S.; Andjelic, C.D.; Pole, A.R.; Rai, P.; Ireland, C.M.; Barrows, L.R. Biologically active components of a Papua New Guinea analgesic and anti-inflammatory lichen preparation. Fitoterapia 2009, 80, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, C.S.; Viswanathan, S.; Reddy, M.K.; Parvathavarthini, S.; Kundu, A.B.; Sukumar, E. Anti-inflammatory activity of (+)-usnic acid. Fitoterapia 2000, 71, 564–566. [Google Scholar] [CrossRef]
- Chen, J.-W.E.; Pedron, S.; Shyu, P.; Hu, Y.; Sarkaria, J.N.; Harley, B.A.C. Influence of Hyaluronic Acid Transitions in Tumor Microenvironment on Glioblastoma Malignancy and Invasive Behavior. Front. Mater. 2018, 5, 39. [Google Scholar] [CrossRef]
- Monslow, J.; Govindaraju, P.; Puré, E. Hyaluronan–a functional and structural sweet spot in the tissue microenvironment. Front. Immunol. 2015, 6, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofuku, K.; Yokouchi, M.; Murayama, T.; Minami, S.; Komiya, S. HAS3-related hyaluronan enhances biological activities necessary for metastasis of osteosarcoma cells. Int. J. Oncol. 2006, 29, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowska, K.; Wróbel, D.; Żmudzki, P.; Podolak, I. Anti-inflammatory activity of saponins from roots of Impatiens parviflora DC. Nat. Prod. Res. 2020, 34, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Galanty, A.; Zagrodzki, P.; Gdula-Argasińska, J.; Grabowska, K.; Koczurkiewicz-Adamczyk, P.; Wróbel-Biedrawa, D.; Podolak, I.; Pękala, E.; Paśko, P. A Comparative Survey of Anti-Melanoma and Anti-Inflammatory Potential of Usnic Acid Enantiomers—A Comprehensive In Vitro Approach. Pharmaceuticals 2021, 14, 945. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Caffo, M.; Minutoli, L.; Marini, H.; Abbritti, R.V.; Squadrito, F.; Trichilo, V.; Valenti, A.; Barresi, V.; Altavilla, D.; et al. ROS and Brain Gliomas: An Overview of Potential and Innovative Therapeutic Strategies. Int. J. Mol. Sci. 2016, 17, 984. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Zheng, X.; Wu, M.-L.; Tian, X.-T.; Song, X.; Liu, Y.-N.; Li, P.-N.; Liu, J. Increased Reactive Oxygen Species and Distinct Oxidative Damage in Resveratrol-suppressed Glioblastoma Cells. J. Cancer 2021, 12, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, K.C.; Gülçin, İ. Anticholinergic and antioxidant activities of usnic acid-An activity-structure insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Hoa, N.T.; Van Bay, M.; Mechler, A.; Vo, Q. V Is Usnic Acid a Promising Radical Scavenger? ACS Omega 2020, 5, 17715–17720. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Wadhwani, A. Antioxidant enzymes and human health. Antioxid. Enzym. 2012, 3, 1–17. [Google Scholar]
- Zhu, Z.; Du, S.; Du, Y.; Ren, J.; Ying, G.; Yan, Z. Glutathione reductase mediates drug resistance in glioblastoma cells by regulating redox homeostasis. J. Neurochem. 2018, 144, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Expósito, M.J.; Martínez-Martos, J.M. The Delicate Equilibrium between Oxidants and Antioxidants in Brain Glioma. Curr. Neuropharmacol. 2019, 17, 342–351. [Google Scholar] [CrossRef]
- Barbosa, M.; Rios, O.; Velásquez, M.; Villalobos, J.; Ehrmanns, J. Acetylcholinesterase and butyrylcholinesterase histochemical activities and tumor cell growth in several brain tumors. Surg. Neurol. 2001, 55, 106–112. [Google Scholar] [CrossRef]
- Reddy, R.G.; Veeraval, L.; Maitra, S.; Chollet-Krugler, M.; Tomasi, S.; Lohezic-Le Devehat, F.; Boustie, J.; Chakravarty, S. Lichen-derived compounds show potential for central nervous system therapeutics. Phytomedicine 2016, 23, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Braganhol, E.; Jäger, E.; Figueiró, F.; Edelweiss, M.I.; Pohlmann, A.R.; Guterres, S.S.; Battastini, A.M.O. Indomethacin-loaded nanocapsules treatment reduces in vivo glioblastoma growth in a rat glioma model. Cancer Lett. 2009, 281, 53–63. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Dudek-Makuch, M.; Chanaj-Kaczmarek, J.; Czepulis, N.; Korybalska, K.; Rutkowski, R.; Łuczak, J.; Grabowska, K.; Bylka, W.; Witowski, J. Anti-inflammatory Activity and Phytochemical Profile of Galinsoga Parviflora Cav. Molecules 2018, 23, 2133. [Google Scholar] [CrossRef] [Green Version]
- Majchrzak-Celińska, A.; Zielińska-Przyjemska, M.; Wierzchowski, M.; Kleszcz, R.; Studzińska-Sroka, E.; Kaczmarek, M.; Paluszczak, J.; Cielecka-Piontek, J.; Krajka-Kuźniak, V. Methoxy-stilbenes downregulate the transcription of Wnt/β-catenin-dependent genes and lead to cell cycle arrest and apoptosis in human T98G glioblastoma cells. Adv. Med. Sci. 2021, 66, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Kikowska, M.A.; Chmielewska, M.; Włodarczyk, A.; Studzińska-Sroka, E.; Żuchowski, J.; Stochmal, A.; Kotwicka, M.; Thiem, B. Effect of pentacyclic triterpenoids-rich callus extract of Chaenomeles japonica (Thunb.) Lindl. ex Spach on viability, morphology, and proliferation of normal human skin fibroblasts. Molecules 2018, 23, 3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parschat, K.; Canne, C.; Hüttermann, J.; Kappl, R.; Fetzner, S. Xanthine dehydrogenase from Pseudomonas putida 86: Specificity, oxidation–reduction potentials of its redox-active centers, and first EPR characterization. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2001, 1544, 151–165. [Google Scholar] [CrossRef]
- Moreira, P.R.; Maioli, M.A.; Medeiros, H.C.D.; Guelfi, M.; Pereira, F.T.V.; Mingatto, F.E. Protective effect of bixin on carbon tetrachloride-induced hepatotoxicity in rats. Biol. Res. 2014, 47, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Padmavathi, B.; Rao, A.R. Modulatory influence of Adhatoda vesica (Justicia adhatoda) leaf extract on the enzymes of xenobiotic metabolism, antioxidant status and lipid peroxidation in mice. Mol. Cell. Biochem. 2000, 213, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres Jr, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Szwajgier, D.; Baranowska-Wójcik, E. Terpenes and phenylpropanoids as acetyl-and butyrylcholinesterase inhibitors: A comparative study. Curr. Alzheimer Res. 2019, 16, 963–973. [Google Scholar] [CrossRef]
- Rhee, I.K.; van Rijn, R.M.; Verpoorte, R. Qualitative determination of false-positive effects in the acetylcholinesterase assay using thin layer chromatography. Phytochem. Anal. An Int. J. Plant Chem. Biochem. Tech. 2003, 14, 127–131. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lichen-Derived Compound/ Extract | A-172 | T98G | ||
|---|---|---|---|---|
| IC50 (µg/mL) | IC50 (µM) | IC50 (µg/mL) | IC50 (µM) | |
| Salazinic acid | >38.8 | >100.0 | >38.8 | >100.0 |
| P. sulcata extract | 73.6 ± 7.3 | - | 89.8 ± 5.1 | - |
| Evernic acid | >33.2 | >100.0 | >33.2 | >100.0 |
| E. prunastri extract | 73.8 ± 5.5 | - | 61.0 ± 1.1 | - |
| (−)-Usnic acid | 31.5 ± 0.8 | 91.4 ± 2.0 | 13.0 ± 1.3 | 37.8 ± 3.8 |
| C. uncialis extract | 11.0 ± 4.6 | - | 3.9 ± 2.2 | - |
| Lichen-Derived Compound/Extract | Inhibition (%) | ||
|---|---|---|---|
| IDO1 | IDO2 | TDO | |
| Salazinic acid | - | - | - |
| P. sulcata extract | 20.5 ± 1.2 | - | - |
| Evernic acid | 32.8 ± 1.9 | - | - |
| E. prunastri extract | 43.1 ± 2.0 | - | - |
| (−)-Usnic acid | 21.6 ± 0.9 | - | - |
| C. uncialis extract | 54.8 ± 3.5 | - | - |
| epacadostat | 95.6 ± 2.8 | - | - |
| Lichen-Derived Compound/Extract | Equivalent Concentration of Acetylsalicylic Acid (mg/mL) | COX-2 Inhibition (%) |
|---|---|---|
| Salazinic acid | 12.9 ± 0.1 | 60.3 ± 3.0 |
| P. sulcata extract | 13.0 ± 0.8 | 65.9 ± 4.1 |
| Evernic acid | 10.9 ± 1.8 | 50.7 ± 2.1 |
| E. prunastri extract | 10.0 ± 0.2 | 35.9 ± 2.8 |
| (−)-Usnic acid | 12.9 ± 1.5 | 59.3 ± 3.5 |
| C. uncialis extract | 10.4 ± 0.1 | 45.9 ± 1.9 |
| Lichen-Derived Compound/Extract | DPPH IC50 (µg/mL) | CUPRAC IC0.5 (µg/mL) |
|---|---|---|
| Salazinic acid | >750.0 | >250.0 |
| P. sulcata extract | 669.3 ± 11.8 | 175.4 ± 1.0 |
| Evernic acid | >750.0 | >250.0 |
| E. prunastri extract | 1926.3 ± 33.2 | 103.4 ± 1.4 |
| (−)-Usnic acid | >750.0 | >250.0 |
| C. uncialis extract | >2500.0 | >312.5 |
| resveratrol | 25.1 ± 0.1 | 29.7 ± 0.1 |
| Lichen-Derived Compound/Extract | SOD Inhibition (%) |
|---|---|
| Salazinic acid | 19.4 ± 0.3 |
| P. sulcata extract | 22.0 ± 1.3 |
| Evernic acid | 21.2 ± 0.0 |
| E. prunastri extract | 53.4 ± 2.4 |
| (−)-Usnic acid | 19.6 ± 0.6 |
| C. uncialis extract | 12.8 ± 1.0 |
| Lichen-Derived Compound/Extract | GR Inhibition under Reaction Conditions (%) | GR Inhibitory Activity (nMol Depleted NADPH/min Incubation) | GPx Inhibition under Reaction Conditions (%) | GPx Inhibitory Activity (nMol Depleted NADPH/min Incubation) |
|---|---|---|---|---|
| Salazinic acid | - | - | - | - |
| P. sulcata extract | - | - | 47.1 ± 0.7 | 93.9 ± 5.2 |
| Evernic acid | - | - | 20.0 ± 2.1 | 39.9 ± 1.0 |
| E. prunastri extract | 91.1 ± 7.2 | 3551.2 ± 264.4 | 92.4 ± 4.3 | 184.2 ± 30.2 |
| (−)-Usnic acid | 18.2 ± 2.2 | 710.2 ± 42.3 | 13.7 ± 2.3 | 27.3 ± 1.3 |
| C. uncialis extract | - | - | - | - |
| Lichen-Derived Compound/Extract | Equivalent Reference Concentration (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Neostygmine | Magniflorine | Donepezil | Eserine | Rivastigmine | ||||||
| AChE | BChE | AChE | BChE | AChE | BChE | AChE | BChE | AChE | BChE | |
| Salazinic acid | - | 13.6 ± 0.1 | - | 43.2 ± 0.2 | - | 7.9 ± 0.3 | - | 9.0 ± 0.1 | - | 70.9 ± 0.3 |
| P. sulacata extract | - | 8.1 ±0.0 | - | 26.2 ± 0.6 | - | 4.8 ± 0.1 | - | 5.4 ± 0.6 | - | 42.9 ± 0.1 |
| Evernic acid | - | 16.5±0.2 | - | 52.3 ± 0.1 | - | 9.5 ± 0.0 | - | 10.9 ± 0.1 | - | 85.9 ± 0.1 |
| E. prunastri extract | - | - | - | - | - | - | - | - | - | - |
| (−)-Usnic acid | - | - | - | - | - | - | - | - | - | - |
| C. uncialis extract | 0.7 ± 0.1 | 16.5 ± 0.1 | 7.9 ± 0.0 | 52.3 ± 0.1 | 0.6 ± 0.1 | 9.5 ± 0.4 | 0.1 ± 0.0 | 10.9 ± 0.0 | 1.4 ± 0.0 | 85.9 ± 0.2 |
| Lichen-Derived Compound/Extract | Pe × 10−6 (cm/s) t = 1 h | Pe × 10−6 (cm/s) t = 4 h |
|---|---|---|
| Salazinic acid (PC) | np | np |
| Salazinic acid (from PSE) | np | np |
| Evernic acid (PC) | 5.2 ± 0.8 | 8.6 ± 0.4 |
| Evernic acid (from EPE) | 5.0 ± 0.7 | 7.2 ± 0.4 |
| (−)-Usnic acid (PC) | 92.8 ± 6.3 | nd |
| (−)-Usnic acid (from CUE) | 140.5 ± 7.3 | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Kaproń, B.; Plech, T.; Żarowski, M.; Cielecka-Piontek, J. Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties. Pharmaceuticals 2021, 14, 1293. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121293
Studzińska-Sroka E, Majchrzak-Celińska A, Zalewski P, Szwajgier D, Baranowska-Wójcik E, Kaproń B, Plech T, Żarowski M, Cielecka-Piontek J. Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties. Pharmaceuticals. 2021; 14(12):1293. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121293
Chicago/Turabian StyleStudzińska-Sroka, Elżbieta, Aleksandra Majchrzak-Celińska, Przemysław Zalewski, Dominik Szwajgier, Ewa Baranowska-Wójcik, Barbara Kaproń, Tomasz Plech, Marcin Żarowski, and Judyta Cielecka-Piontek. 2021. "Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties" Pharmaceuticals 14, no. 12: 1293. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121293