
Identification of a Thyroid Hormone Derivative as a Pleiotropic Agent for the Treatment of Alzheimer’s Disease

 ,
,  , ,
, ,  ,
,  , and
, and 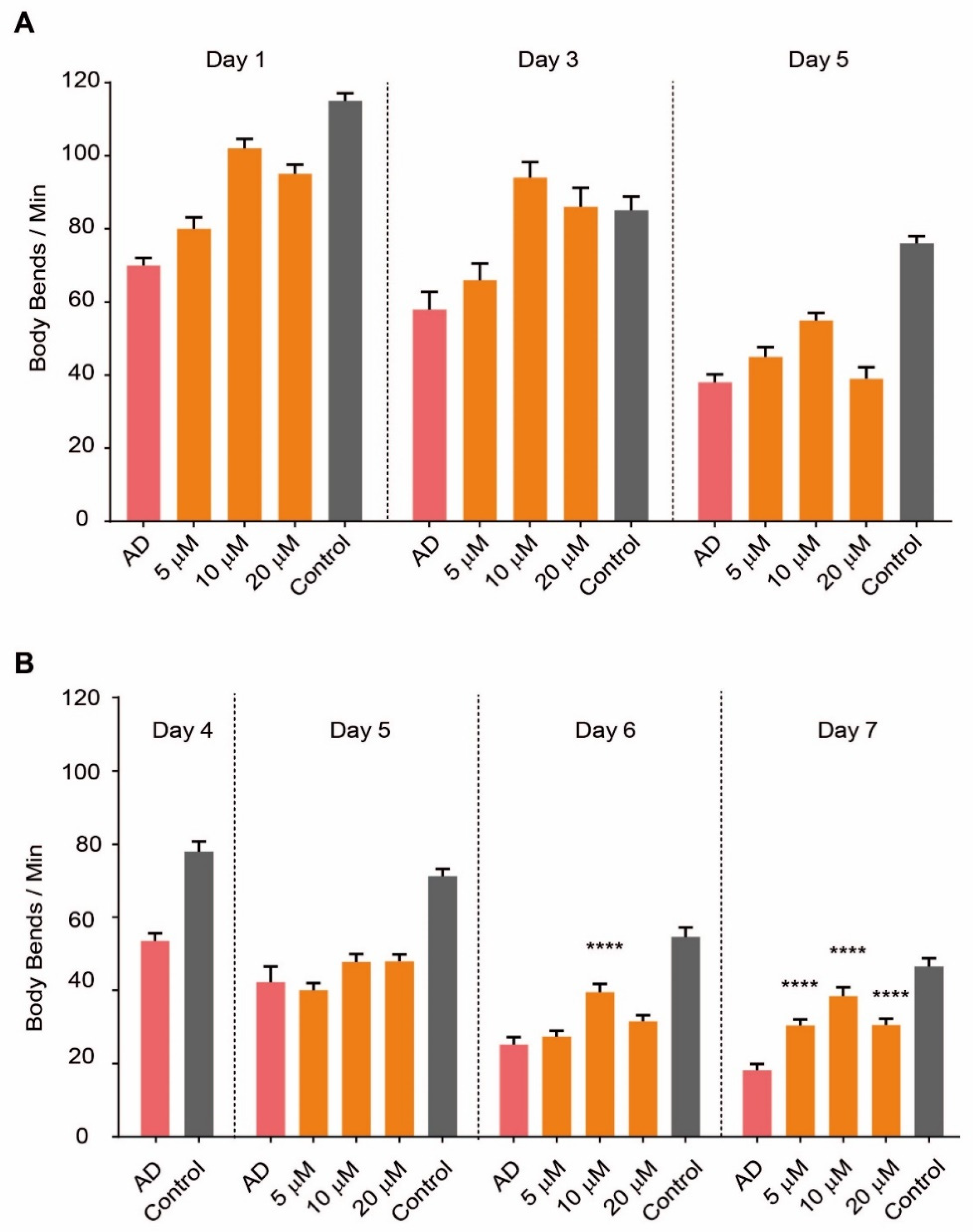{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SG2 Restores Motility in a C. elegans Model of AD
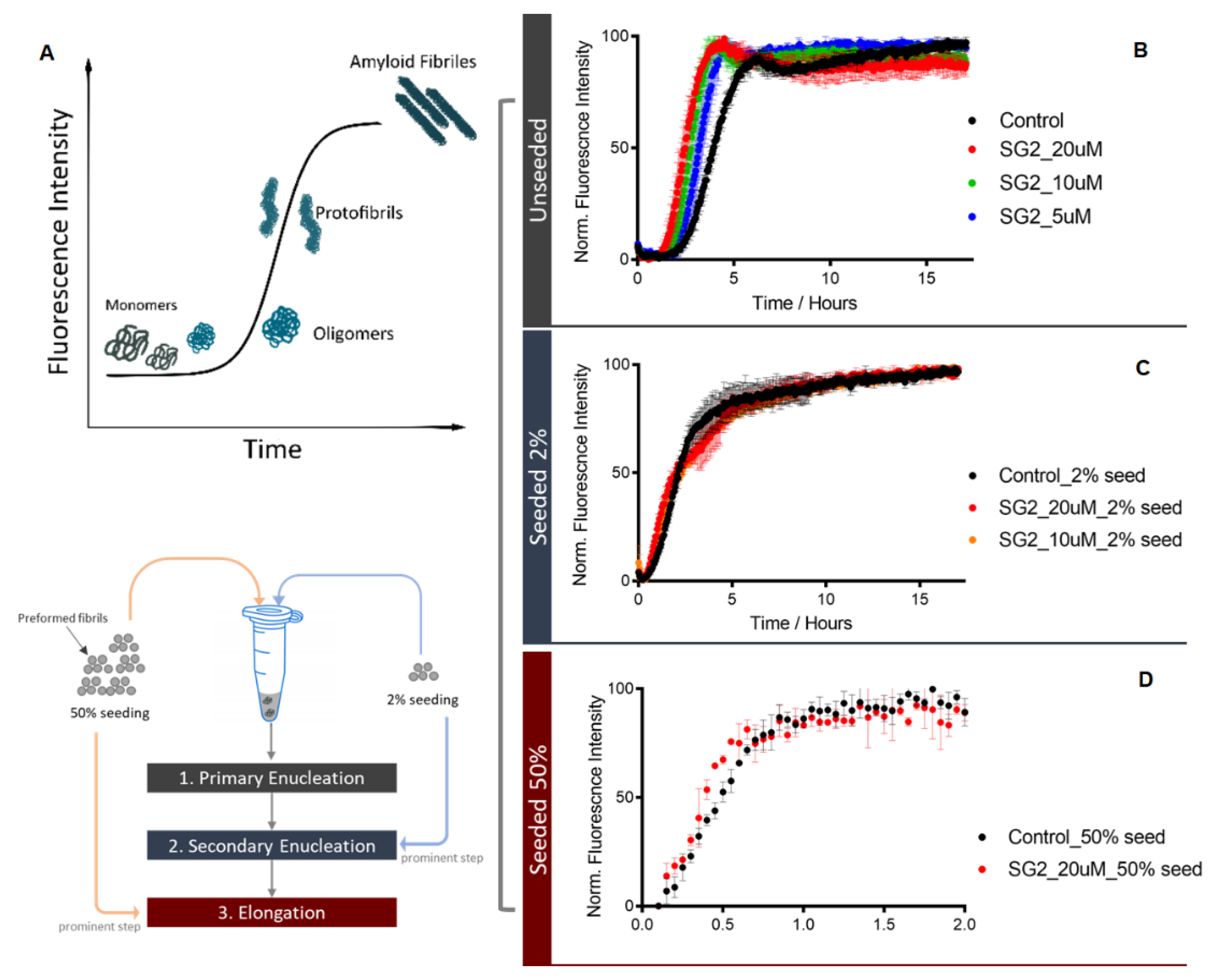
2.2. Studies on Aβ Aggregation Process In Vitro
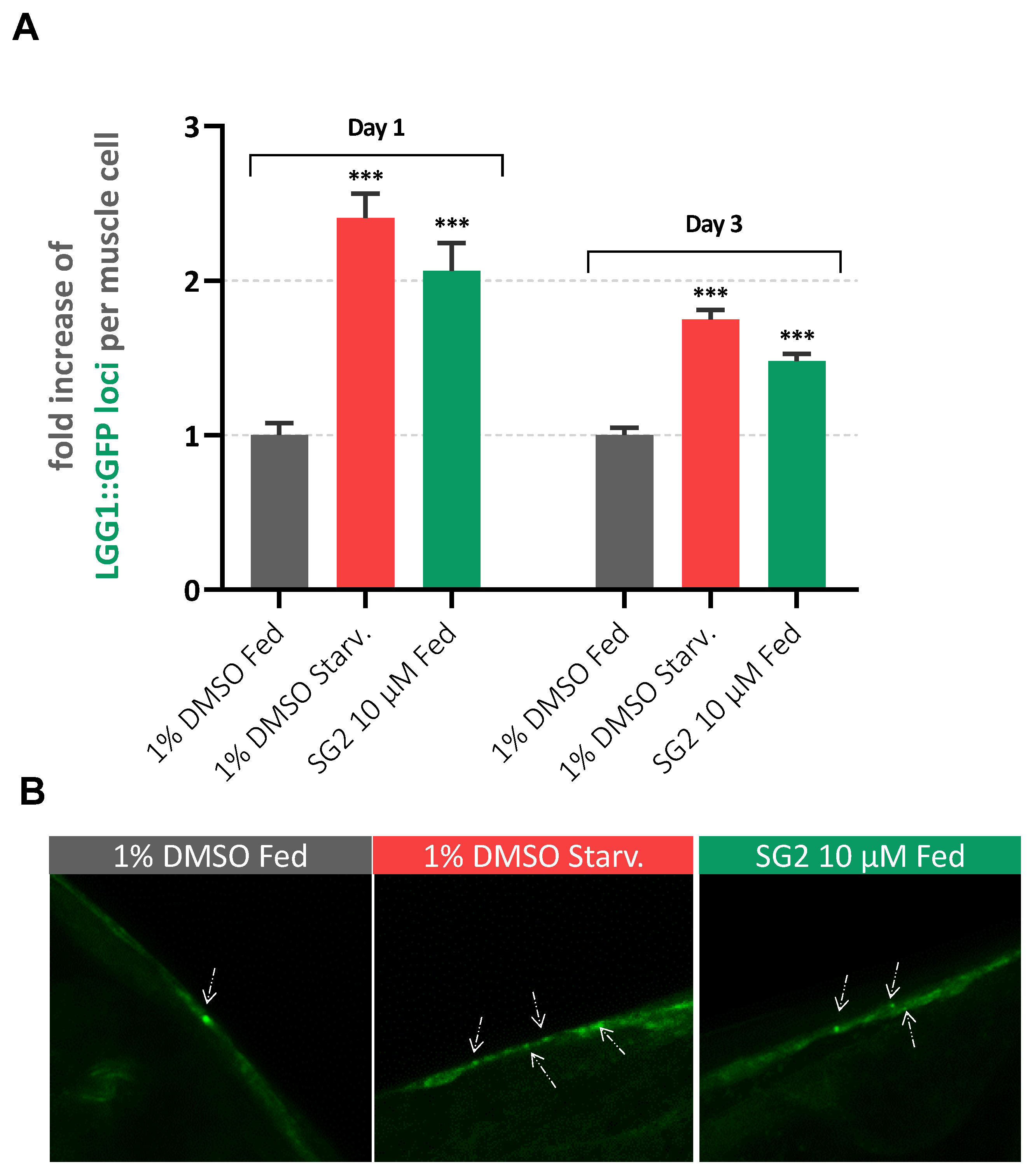
2.3. SG2 Activates Autophagy in a C. elegans Model of AD
2.4. Effects of SG2 in a Cell Model of Aβ-Induced Neurotoxicity
2.5. In Vitro ADME-Tox Profiling of SG2
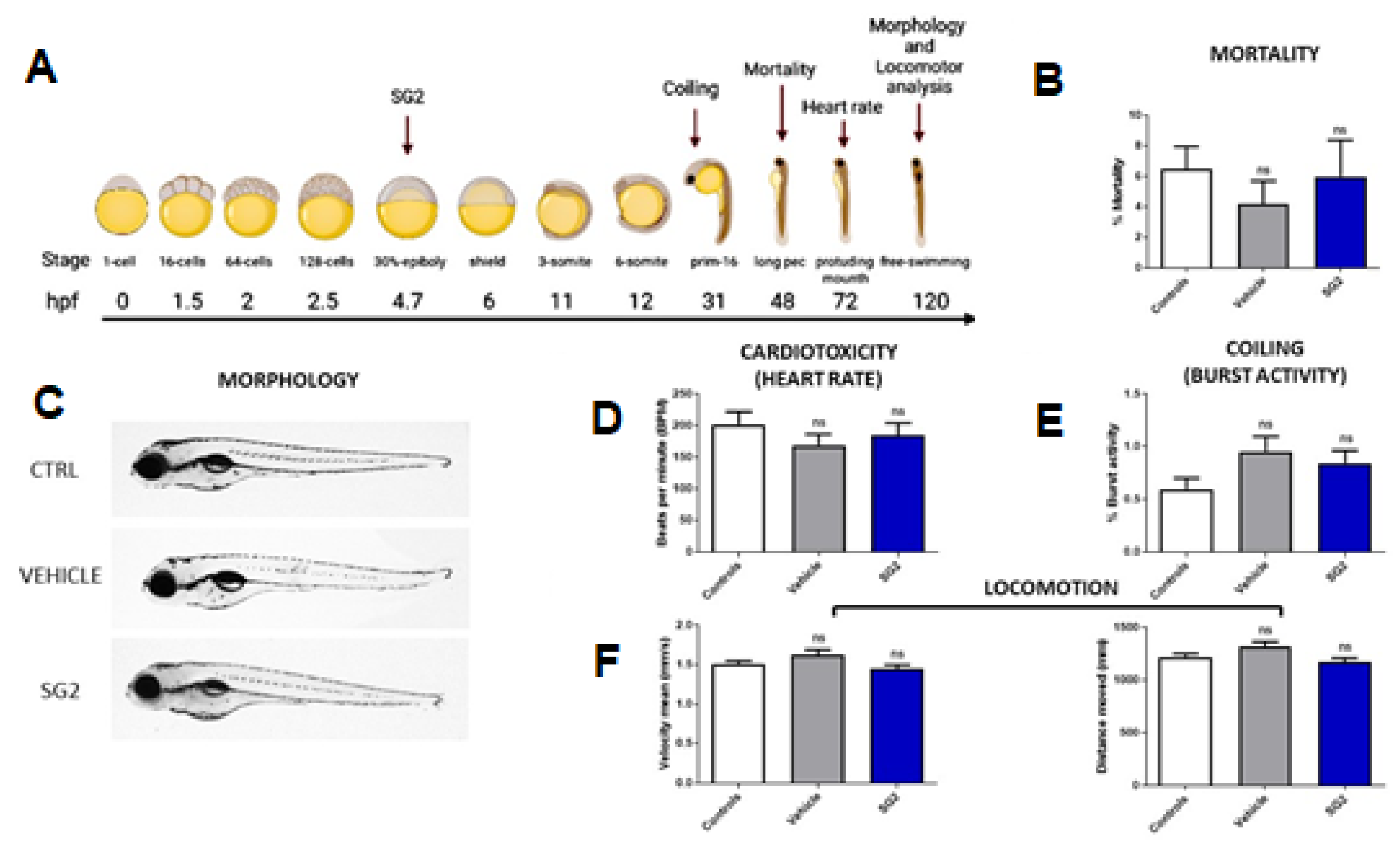
2.6. In Vivo Toxicological Analysis of SG2 in Zebrafish (Danio Rerio)
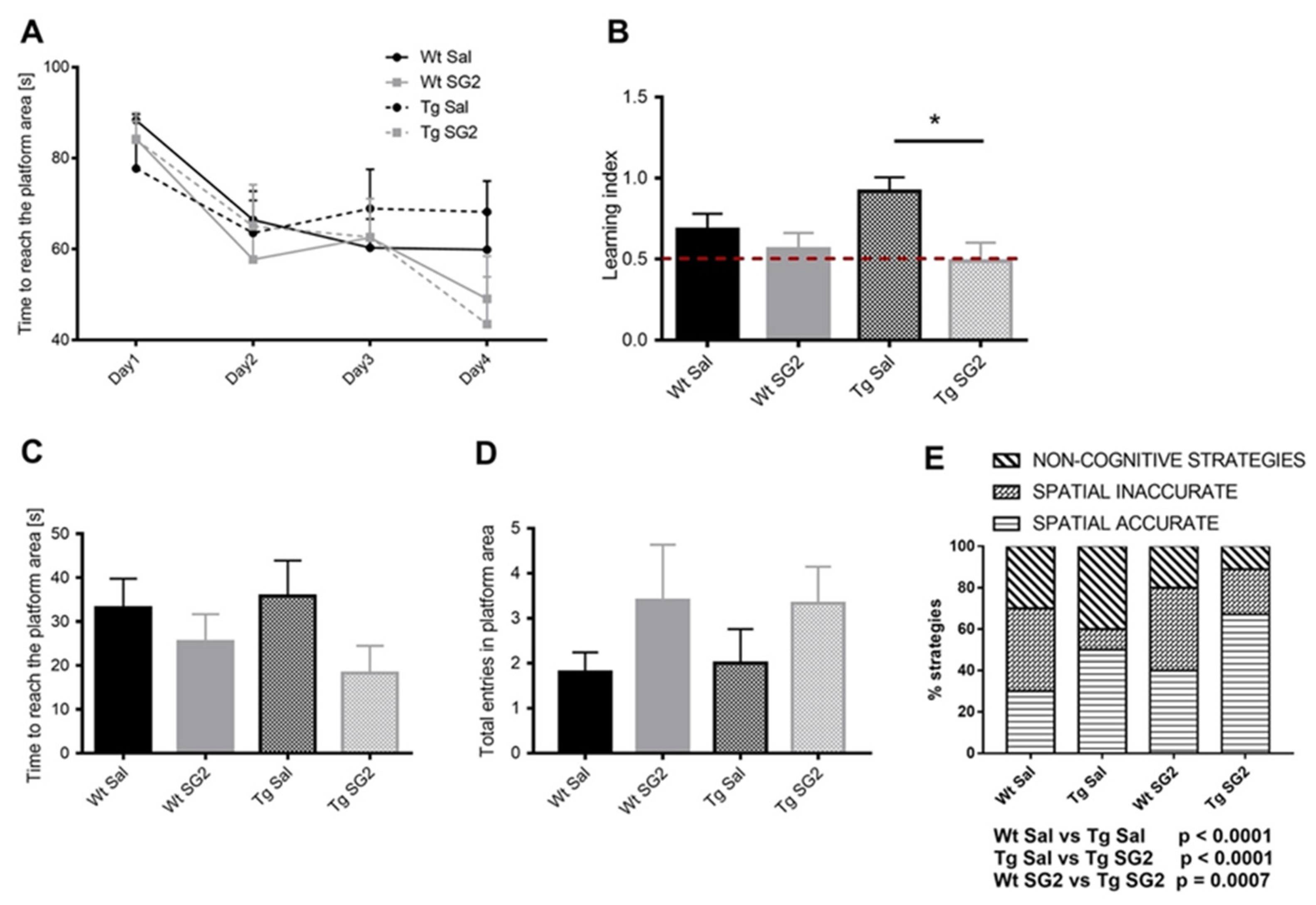
2.7. SG2 Treatment Improves Learning and Memory in 5XFAD Mice
3. Discussion
4. Materials and Methods
4.1. Aβ Aggregation
4.2. C. elegans Experiments
4.2.1. Media
4.2.2. Strains
4.2.3. Compound Administration
4.2.4. Automated C. elegans Motility Assays
4.2.5. Confocal Microscopy
4.3. Zebrafish Experiments
4.3.1. Zebrafish Husbandry
4.3.2. Toxicological Analyses on Zebrafish Embryos and Larvae
4.4. Mice Experiments
4.4.1. Mouse Model
4.4.2. Drugs and Procedures
4.4.3. Behavioural Analysis
4.4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C. The amyloid-β pathway in alzheimer’s disease. Mol. Psychiatry 2021, 1–23. [Google Scholar] [CrossRef]
- Mrak, R.E.; Griffin, W.S.T.; Graham, D.I. Aging-associated changes in human brain. J. Neuropathol. Exp. Neurol. 1997, 56, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Aufschnaiter, A.; Kohler, V.; Diessl, J.; Peselj, C.; Carmona-Gutierrez, D.; Keller, W.; Büttner, S. Mitochondrial lipids in neurodegeneration. Cell Tissue Res. 2017, 367, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and alzheimer’s disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar] [CrossRef]
- Soria Lopez, J.A.; González, H.M.; Léger, G.C. Alzheimer’s disease. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 167, pp. 231–255. [Google Scholar]
- Guix, F.X. The interplay between aging-associated loss of protein homeostasis and extracellular vesicles in neurodegeneration. J. Neurosci. Res. 2020, 98, 262–283. [Google Scholar] [CrossRef] [PubMed]
- Imbimbo, B.P.; Ippati, S.; Watling, M. Should drug discovery scientists still embrace the amyloid hypothesis for alzheimer’s disease or should they be looking elsewhere? Expert Opin. Drug Discov. 2020, 15, 1241–1251. [Google Scholar] [CrossRef]
- Bostancıklıoğlu, M. An update on the interactions between Alzheimer’s disease, autophagy and inflammation. Gene 2019, 705, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Oliver, D.M. Amyloid beta and phosphorylated tau-induced defective autophagy and mitophagy in Alzheimer’s disease. Cells 2019, 8, 488. [Google Scholar] [CrossRef] [Green Version]
- Mullane, K.; Williams, M. Alzheimer’s disease beyond amyloid: Can the repetitive failures of amyloid-targeted therapeutics inform future approaches to dementia drug discovery? Biochem. Pharmacol. 2020, 177, 113945. [Google Scholar] [CrossRef]
- Cummings, J.; Lee, G.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2021. Alzheimer’s Dement. 2021, 7, e12179. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J. Why aducanumab is important. Nat. Med. 2021, 27, 1498. [Google Scholar] [CrossRef]
- Frautschy, S.A.; Cole, G.M. Why pleiotropic interventions are needed for alzheimer’s disease. Mol. Neurobiol. 2010, 41, 392–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulterijs, S.; Braeckman, B.P. Phenotypic screening in C. elegans as a tool for the discovery of new geroprotective drugs. Pharmaceuticals 2020, 13, 164. [Google Scholar] [CrossRef]
- Swinney, D.C. Phenotypic vs. Target-based drug discovery for first-in-class medicines. Clin. Pharmacol. Ther. 2013, 93, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Chiellini, G.; Nesi, G.; Digiacomo, M.; Malvasi, R.; Espinoza, S.; Sabatini, M.; Frascarelli, S.; Laurino, A.; Cichero, E.; Macchia, M.; et al. Design, synthesis, and evaluation of thyronamine analogues as novel potent mouse trace amine associated receptor 1 (m taar1) agonists. J. Med. Chem. 2015, 58, 5096–5107. [Google Scholar] [CrossRef]
- Choi, B.W.; Kim, S.; Kang, S.; Won, K.S.; Yi, H.A.; Kim, H.W. Relationship between thyroid hormone levels and the pathology of alzheimer’s disease in euthyroid subjects. Thyroid Off. J. Am. Thyroid Assoc. 2020, 30, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Byun, M.S.; Yi, D.; Sohn, B.K.; Lee, J.H.; Lee, J.Y.; Kim, Y.K.; Lee, D.Y. Associations of thyroid hormone serum levels with in-vivo alzheimer’s disease pathologies. Alzheimer’s Res. Ther. 2017, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, P.; Horvath, A.; Eckerström, C.; Wallin, A.; Svensson, J. Altered thyroid hormone profile in patients with alzheimer’s disease. Psychoneuroendocrinology 2020, 121, 104844. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.S.; Vasan, R.S. Thyroid function and alzheimer’s disease. J. Alzheimer’s Dis. JAD 2009, 16, 503–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellusci, L.; Laurino, A.; Sabatini, M.; Sestito, S.; Lenzi, P.; Raimondi, L.; Rapposelli, S.; Biagioni, F.; Fornai, F.; Salvetti, A.; et al. New insights into the potential roles of 3-iodothyronamine (t1am) and newly developed thyronamine-like taar1 agonists in neuroprotection. Front. Pharmacol. 2017, 8, 905. [Google Scholar] [CrossRef] [Green Version]
- Rogowski, M.; Bellusci, L.; Sabatini, M.; Rapposelli, S. Lipolytic effects of 3-iodothyronamine (t1am) and a novel thyronamine-like analog sg-2 through the ampk pathway. Int. J. Mol. Sci. 2019, 20, 4054. [Google Scholar] [CrossRef] [Green Version]
- Bellusci, L.; Runfola, M.; Carnicelli, V.; Sestito, S.; Fulceri, F.; Santucci, F.; Lenzi, P.; Fornai, F. Endogenous 3-iodothyronamine (t1am) and synthetic thyronamine-like analog sg-2 act as novel pleiotropic neuroprotective agents through the modulation of sirt6. Molecules 2020, 25, 1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McColl, G.; Roberts, B.R.; Pukala, T.L.; Kenche, V.B.; Roberts, C.M.; Link, C.D.; Ryan, T.M.; Masters, C.L.; Barnham, K.J.; Bush, A.I.; et al. Utility of an improved model of amyloid-beta (aβ1-42) toxicity in caenorhabditis elegans for drug screening for alzheimer’s disease. Mol. Neurodegener. 2012, 7, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habchi, J.; Arosio, P.; Perni, M.; Costa, A.R.; Yagi-Utsumi, M.; Joshi, P. An anticancer drug suppresses the primary nucleation reaction that initiates the production of the toxic aβ42 aggregates linked with alzheimer’s disease. Sci. Adv. 2016, 2, e1501244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limbocker, R.; Chia, S.; Ruggeri, F.S.; Perni, M. Trodusquemine enhances aβ(42) aggregation but suppresses its toxicity by displacing oligomers from cell membranes. Nat. Commun. 2019, 10, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habchi, J.; Chia, S.; Limbocker, R.; Mannini, B.; Ahn, M.; Perni, M.; Hansson, O.; Arosio, P.; Kumita, J.R.; Challa, P.K.; et al. Systematic development of small molecules to inhibit specific microscopic steps of aβ42 aggregation in alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2017, 114, E200–E208. [Google Scholar] [CrossRef] [Green Version]
- Perni, M.; Challa, P.K.; Kirkegaard, J.B.; Limbocker, R.; Koopman, M.; Hardenberg, M.C.; Sormanni, P.; Muller, T.; Saar, K.L.; Roode, L.W.Y.; et al. Massively parallel C. elegans tracking provides multi-dimensional fingerprints for phenotypic discovery. J. Neurosci. Methods 2018, 306, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Koopman, M.; Peter, Q. Assessing motor-related phenotypes of caenorhabditis elegans with the wide field-of-view nematode tracking platform. Nat. Protoc. 2020, 15, 2071–2106. [Google Scholar] [CrossRef]
- Cohen, S.I.A.; Arosio, P.; Presto, J.; Kurudenkandy, F.R.; Biverstal, H.; Dolfe, L.; Dunning, C.; Yang, X.; Frohm, B.; Vendruscolo, M.; et al. A molecular chaperone breaks the catalytic cycle that generates toxic aβ oligomers. Nat. Struct. Mol. Biol. 2015, 22, 207–213. [Google Scholar] [CrossRef]
- Cohen, S.I.; Linse, S.; Luheshi, L.M.; Hellstrand, E.; White, D.A.; Rajah, L.; Otzen, D.E.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P. Proliferation of amyloid-β42 aggregates occurs through a secondary nucleation mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 9758–9763. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Abeliovich, H.; Agostinis, P.; Agrawal, D.K.; Aliev, G.; Askew, D.S.; Baba, M.; Baehrecke, E.H.; Bahr, B.A.; Ballabio, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008, 4, 151–175. [Google Scholar] [CrossRef]
- Palmisano, N.J.; Meléndez, A. Detection of autophagy in caenorhabditis elegans using gfp::Lgg-1 as an autophagy marker. Cold Spring Harbor Protoc. 2016, 2016, pdb-prot086496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feleciano, D.R.; Juenemann, K.; Iburg, M.; Brás, I.C.; Holmberg, C.I.; Kirstein, J. Crosstalk between chaperone-mediated protein disaggregation and proteolytic pathways in aging and disease. Front. Aging Neurosci. 2019, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, E.; Lantz, S.M.; Tobón-Velasco, J.C.; Newport, G.D.; Wu, Q.; Virmani, A.; Ali, S.F.; Santamaría, A. On the in vivo early toxic properties of aβ25–35 peptide in the rat hippocampus: Involvement of the receptor-for-advanced glycation-end-products and changes in gene expression. Neurotoxicol. Teratol. 2011, 33, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, Y.G.; Marlatt, M.W.; Smith, M.A.; Kosenko, E.A. Subcellular and metabolic examination of amyloid-β peptides in alzheimer disease pathogenesis: Evidence for aβ25–35. Exp. Neurol. 2010, 221, 26–37. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. Mtor signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, W. Sirtuins: From metabolic regulation to brain aging. Front. Aging Neurosci. 2013, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Cummings, J.; Decourt, B.; Leverenz, J.B.; Sabbagh, M.N. Clinical drug development for dementia with lewy bodies: Past and present. Expert Opin. Investig. Drugs 2019, 28, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Macri, C.; Wang, F.; Tasset, I.; Schall, N.; Page, N.; Briand, J.-P.; Cuervo, A.M.; Muller, S. Modulation of deregulated chaperone-mediated autophagy by a phosphopeptide. Autophagy 2015, 11, 472–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runfola, M.; Sestito, S.; Bellusci, L.; La Pietra, V.; D’Amore, V.M.; Kowalik, M.A.; Chiellini, G.; Gul, S.; Perra, A.; Columbano, A.; et al. Design, synthesis and biological evaluation of novel trβ selective agonists sustained by adme-toxicity analysis. Eur. J. Med. Chem. 2020, 188, 112006. [Google Scholar] [CrossRef] [PubMed]
- Niles, A.L.; Moravec, R.A.; Riss, T.L. In vitro viability and cytotoxicity testing and same-well multi-parametric combinations for high throughput screening. Curr. Chem. Genom. 2009, 3, 33–41. [Google Scholar] [CrossRef]
- Runfola, M.; Sestito, S.; Gul, S.; Chiellini, G.; Rapposelli, S. Collecting data through high throughput in vitro early toxicity and off-target liability assays to rapidly identify limitations of novel thyromimetics. Data Brief 2020, 29, 105206. [Google Scholar] [CrossRef] [PubMed]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef] [PubMed]
- Landel, V.; Baranger, K.; Virard, I.; Loriod, B.; Khrestchatisky, M.; Rivera, S.; Benech, P.; Féron, F. Temporal gene profiling of the 5xfad transgenic mouse model highlights the importance of microglial activation in alzheimer’s disease. Mol. Neurodegener. 2014, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- De Strooper, B.; Karran, E. The cellular phase of alzheimer’s disease. Cell 2016, 164, 603–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R.A.; Singh, B.K.; Zhou, J.; Wu, Y.; Farah, B.L.; Ohba, K.; Lesmana, R.; Gooding, J.; Bay, B.-H.; Yen, P.M. Thyroid hormone induction of mitochondrial activity is coupled to mitophagy via ros-ampk-ulk1 signaling. Autophagy 2015, 11, 1341–1357. [Google Scholar] [CrossRef] [Green Version]
- Lesmana, R.; Sinha, R.A.; Singh, B.K.; Zhou, J.; Ohba, K.; Wu, Y.; Yau, W.W.; Bay, B.-H.; Yen, P.M. Thyroid hormone stimulation of autophagy is essential for mitochondrial biogenesis and activity in skeletal muscle. Endocrinology 2016, 157, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, S. The genetics of caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Zhang, H.; Chang, J.T.; Guo, B.; Hansen, M.; Jia, K.; Kovács, A.L.; Kumsta, C.; Lapierre, L.R.; Legouis, R.; Lin, L.; et al. Guidelines for monitoring autophagy in caenorhabditis elegans. Autophagy 2015, 11, 9–27. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- MacArthur Clark, J. The 3rs in research: A contemporary approach to replacement, reduction and refinement. Br. J. Nutr. 2018, 120, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Brandeis, R.; Brandys, Y.; Yehuda, S. The use of the morris water maze in the study of memory and learning. Int. J. Neurosci. 1989, 48, 29–69. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, D.; Bitto, A.; Galantucci, M.; Zaffe, D.; Ottani, A.; Irrera, N.; Neri, L.; Cavallini, G.M.; Altavilla, D.; Botticelli, A.R.; et al. Melanocortins protect against progression of alzheimer’s disease in triple-transgenic mice by targeting multiple pathophysiological pathways. Neurobiol. Aging 2014, 35, 537–547. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daini, E.; Secco, V.; Liao, W.; Zoli, M.; Vilella, A. A regional and cellular analysis of the early intracellular and extracellular accumulation of aβ in the brain of 5xfad mice. Neurosci. Lett. 2021, 754, 135869. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Runfola, M.; Perni, M.; Yang, X.; Marchese, M.; Bacci, A.; Mero, S.; Santorelli, F.M.; Polini, B.; Chiellini, G.; Giuliani, D.; et al. Identification of a Thyroid Hormone Derivative as a Pleiotropic Agent for the Treatment of Alzheimer’s Disease. Pharmaceuticals 2021, 14, 1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121330
Runfola M, Perni M, Yang X, Marchese M, Bacci A, Mero S, Santorelli FM, Polini B, Chiellini G, Giuliani D, et al. Identification of a Thyroid Hormone Derivative as a Pleiotropic Agent for the Treatment of Alzheimer’s Disease. Pharmaceuticals. 2021; 14(12):1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121330
Chicago/Turabian StyleRunfola, Massimiliano, Michele Perni, Xiaoting Yang, Maria Marchese, Andrea Bacci, Serena Mero, Filippo M. Santorelli, Beatrice Polini, Grazia Chiellini, Daniela Giuliani, and et al. 2021. "Identification of a Thyroid Hormone Derivative as a Pleiotropic Agent for the Treatment of Alzheimer’s Disease" Pharmaceuticals 14, no. 12: 1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14121330