
Effects of GPR18 Ligands on Body Weight and Metabolic Parameters in a Female Rat Model of Excessive Eating

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
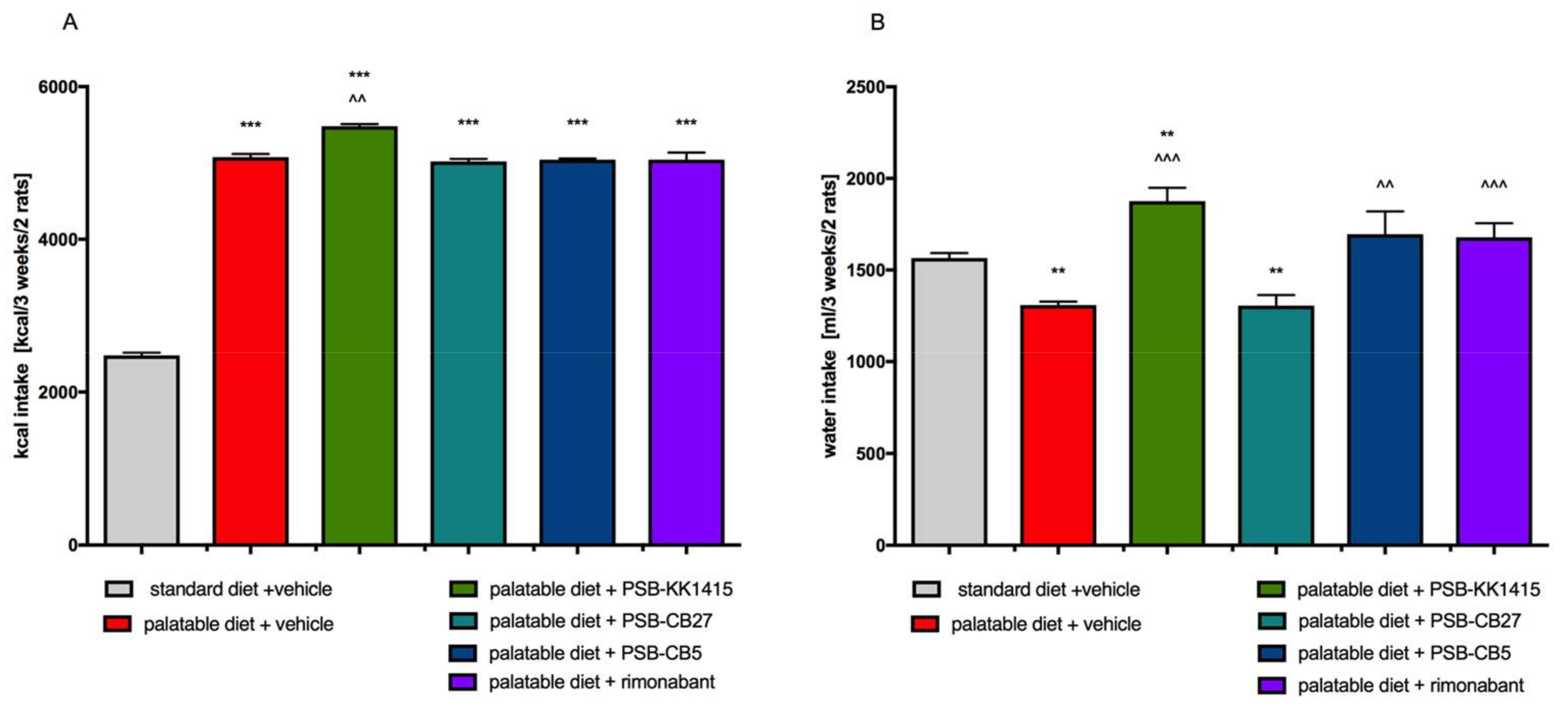
2.1. Effect of Diet or Tested Compounds on Body Weight, Amount of Intraperitoneal Adipose Tissue, Caloric, and Water Intakes
2.2. Effect of Diet or Tested Compounds on the Weight of Selected Organs
2.3. Effect of Diet or Tested Compounds on Caloric and Water Intakes
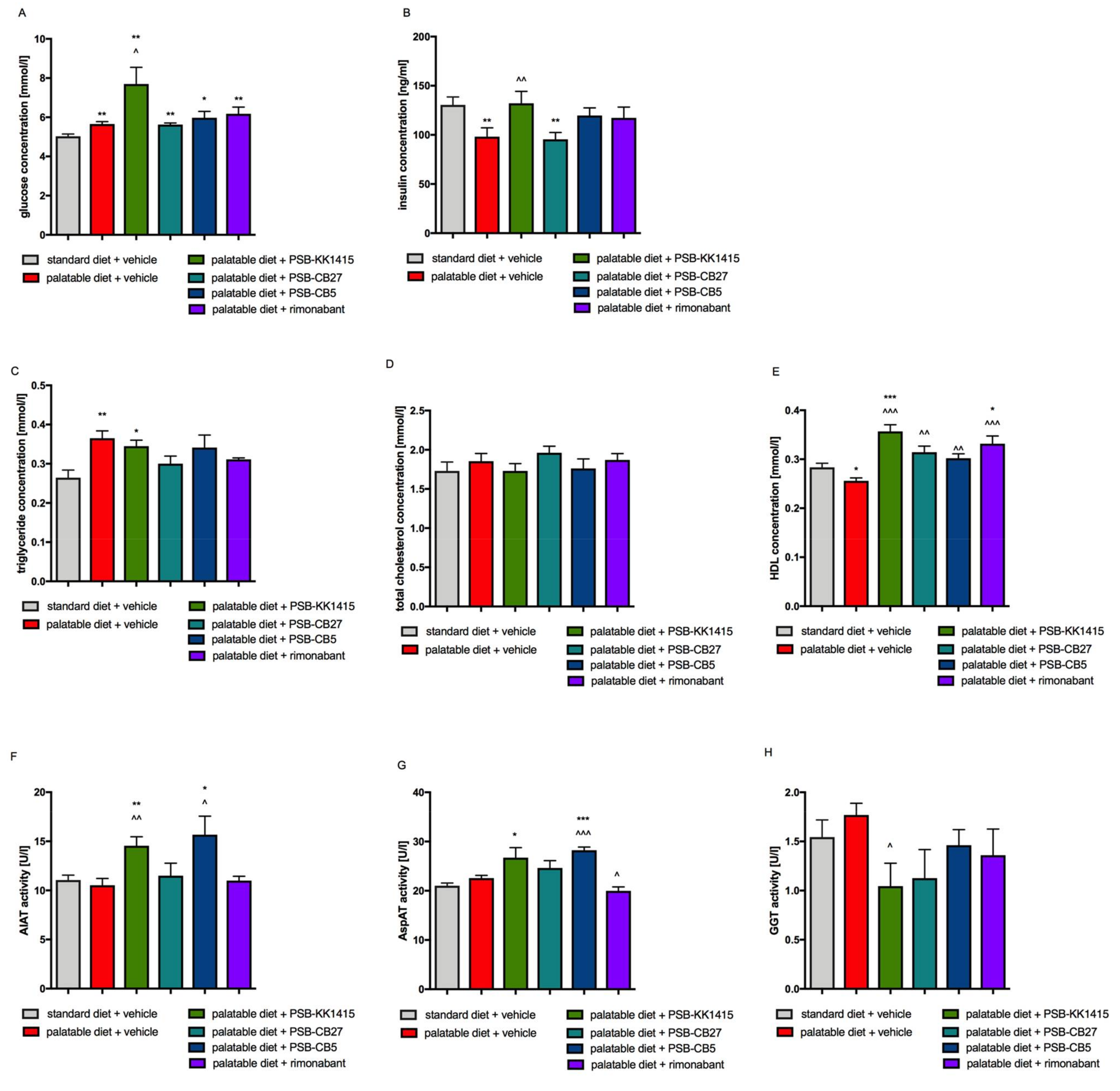
2.4. Influence of Diet or Tested Compounds on Plasma Glucose and Insulin Levels
2.5. Influence of Diet or Tested Compounds on Lipid Profile
2.6. Influence of Diet or Tested Compounds on Alanine Aminotransferase (AlAT), Aspartate Aminotransferase (AspAT), and γ-Glutamyl Transpeptidase (GGT) Activity in Rats Plasma
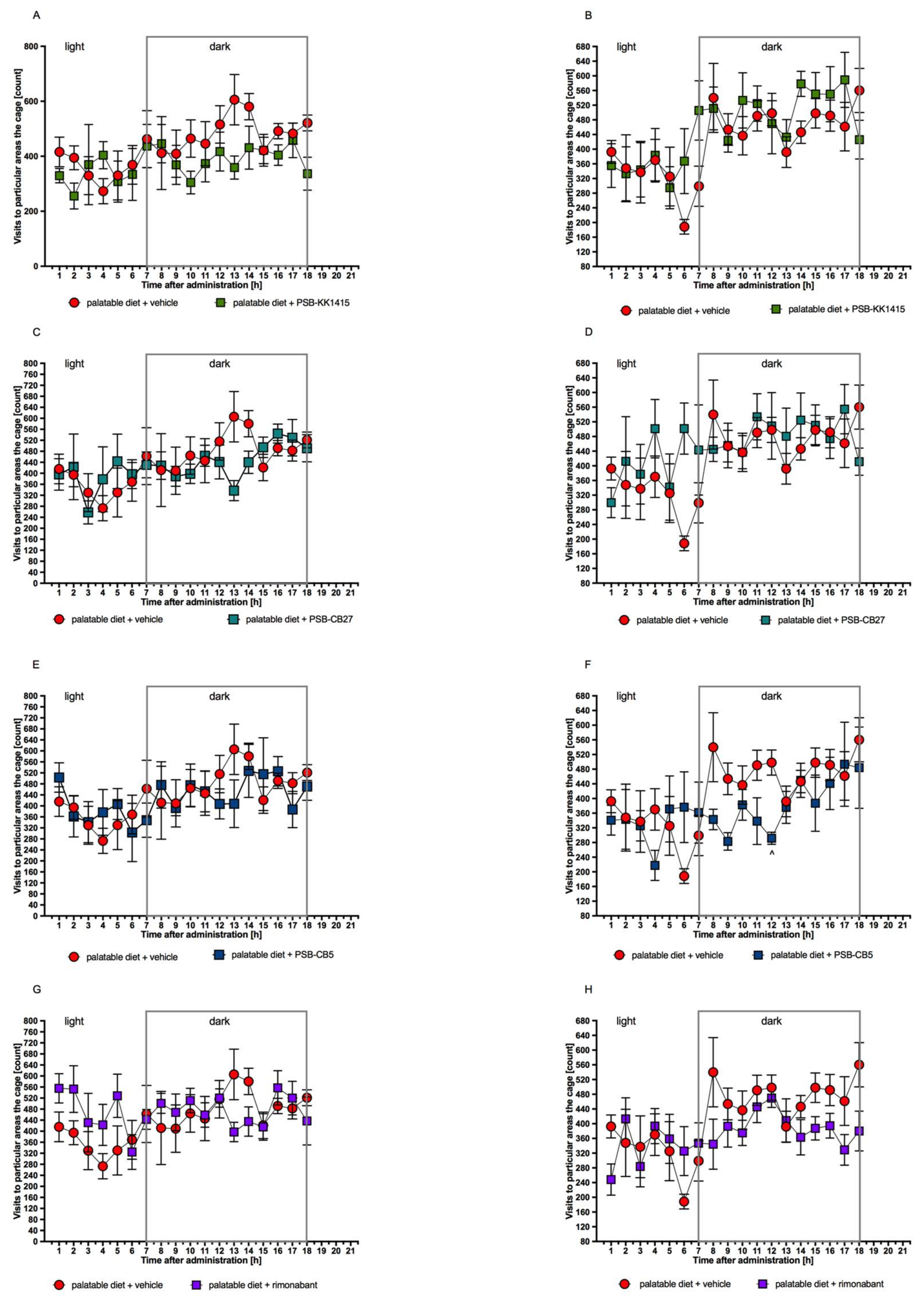
2.7. Influence of Single and Chronic Administration of the Tested Compounds on Locomotor Activity of Rats Fed Palatable Diet and Housed in Pairs in Home Cages
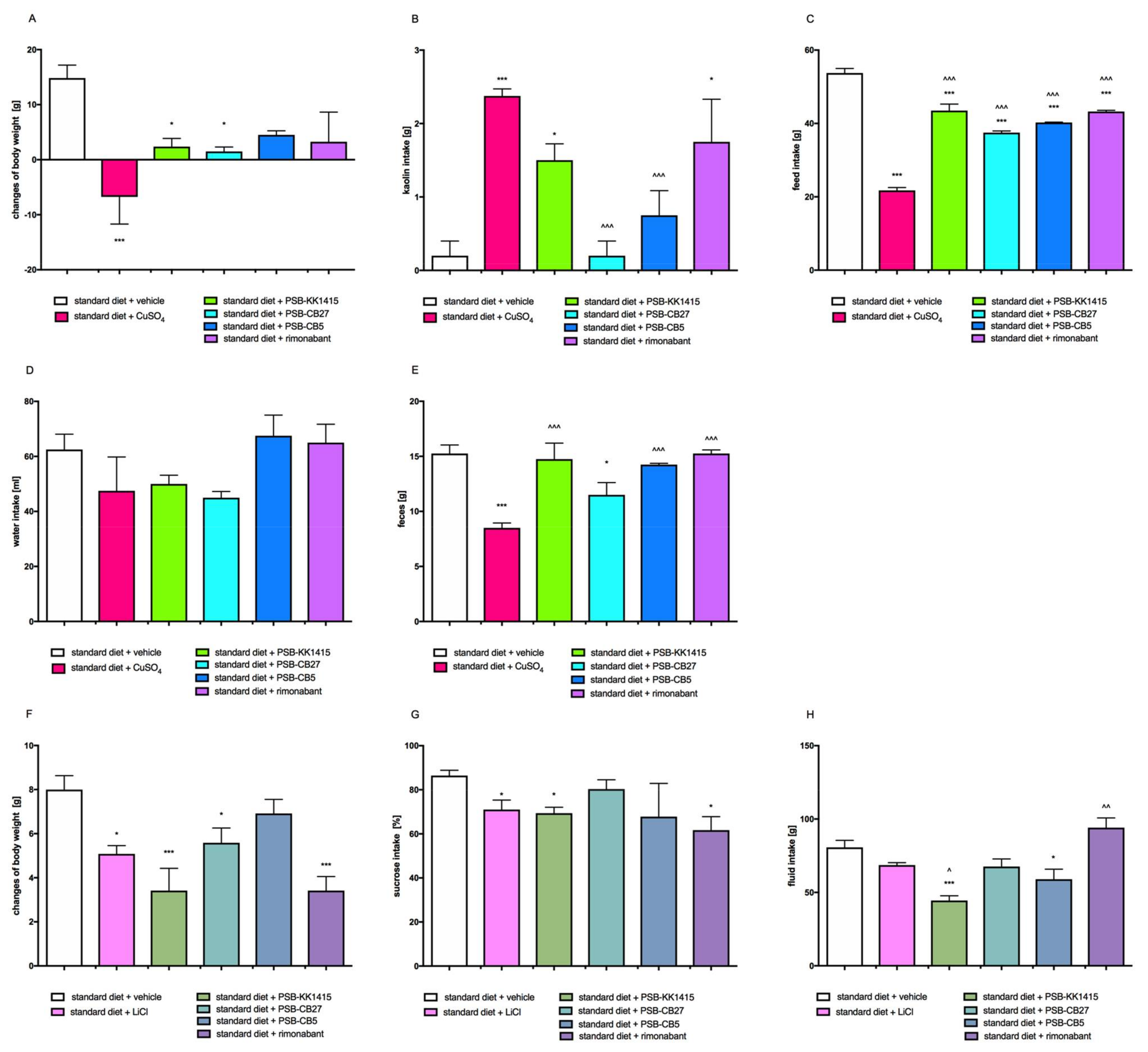
2.8. Effects on Visceral Illness via Measurement of Kaolin Intake (Pica Behavior Model)
2.9. Effects on Visceral Illness via Measurement of Saccharin Intake
3. Discussion
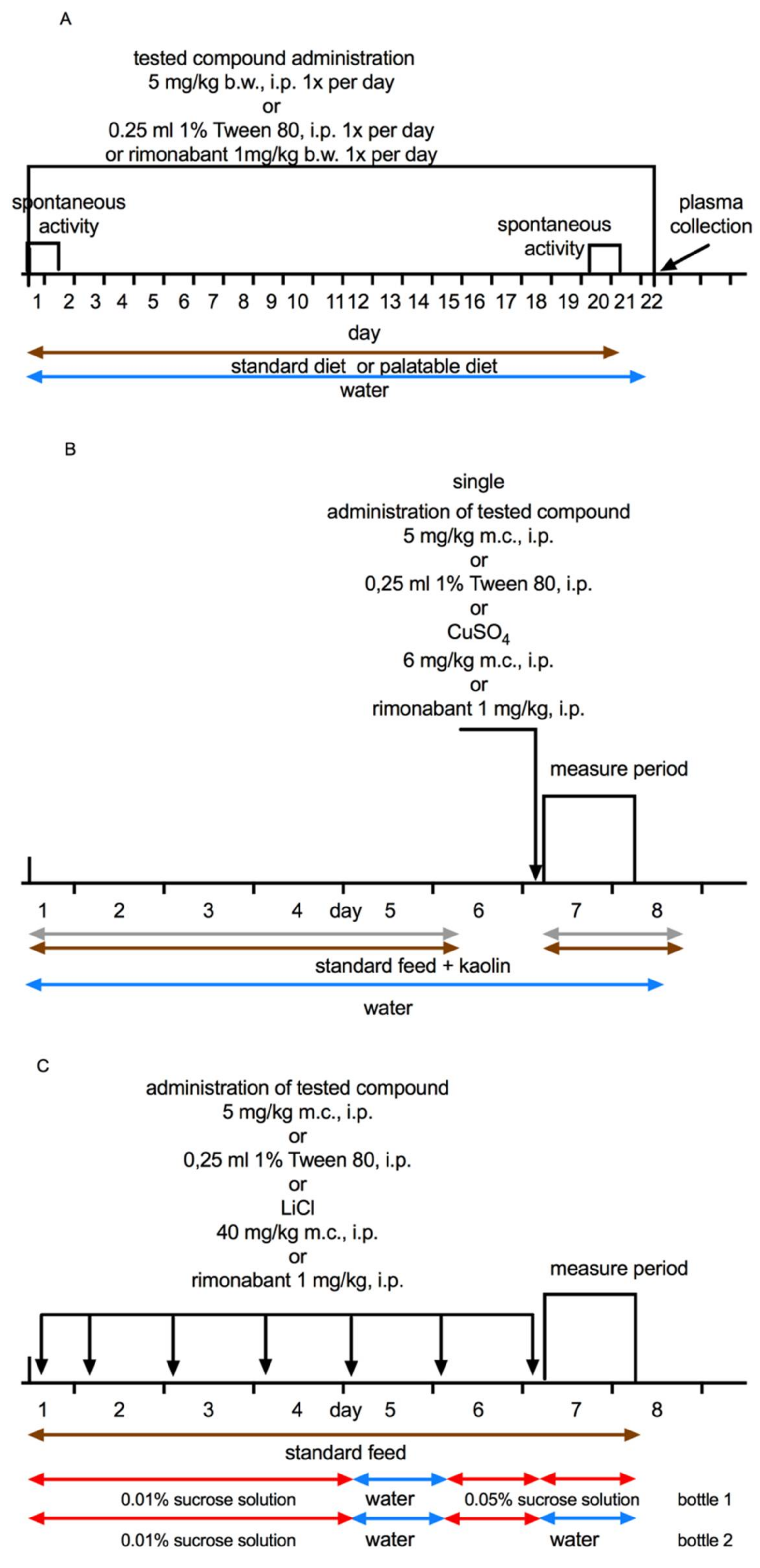
4. Materials and Methods
4.1. Animals and Tested Compounds
4.2. Drugs, Chemical Reagents, and Other Materials
4.3. Effect of Tested Ligands on Body Weight, Locomotor Activity, and Food and Water Intake
4.4. Influence of Tested Compounds on Lipid Profile, Glucose and Insulin Levels as Well as AlAT, AspAT, and GGT Activity in Plasma
4.5. Effects on Visceral Illness via Measurement of Kaolin Intake (Pica Behaviour)
4.6. Effects on Visceral Illness via Measurement of Sucrose Intake (Conditioned Tasted Aversion; CTA)
4.7. Data Analysis and Statistical Procedures
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poleszak, E.; Wośko, S.; Sławińska, K.; Szopa, A.; Wróbel, A.; Serefko, A. Cannabinoids in depressive disorders. Life Sci. 2018. [Google Scholar] [CrossRef]
- Adel, Y.; Alexander, S.P.H. Neuromolecular Mechanisms of Cannabis Action. Adv. Exp. Med. Biol. 2021, 1264, 15–28. [Google Scholar] [PubMed]
- Sun, Y.; Alexander, S.P.; Kendall, D.A.; Bennett, A.J. Cannabinoids and PPARalpha signalling. Biochem. Soc. Trans. 2006, 34, 1095–1097. [Google Scholar] [CrossRef] [PubMed]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Basavarajappa, B.S. Neuropharmacology of the endocannabinoid signaling system-molecular mechanisms, biological actions and synaptic plasticity. Curr. Neuropharmacol. 2007, 5, 81–97. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V.; Fontana, A.; Cadas, H.; Schinelli, S.; Cimino, G.; Schwartz, J.C.; Piomelli, D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 388, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Morales, R.; Reggio, P.H. Emerging roles of cannabinoids and synthetic cannabinoids in clinical experimental models. Adv. Exp. Med. Biol. 2021, 1264, 47–65. [Google Scholar]
- Kuder, J.K.; Karcz, T.; Kaleta, M.; Kieć-Kononowicz, K. Molecular modeling of an orphan GPR18 receptor. Lett. Drug Design Discov. 2018, 8, 1167–1174. [Google Scholar] [CrossRef]
- Rajaraman, G.; Simcocks, A.; Hryciw, D.H.; Hutchinson, D.S.; McAinch, A.J. G protein coupled receptor 18: A potential role for endocannaninoid signaling in metabolic dysfunction. Mol. Nutr. Food Res. 2016, 60, 92–102. [Google Scholar] [CrossRef]
- Chiang, N.; Dalli, J.; Colas, R.A.; Serhan, C.N. Identification of resolvin D2 receptor mediating resolution of infections and organ protection. J. Exp. Med. 2015, 212, 1203–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ9-THC and N-arachidonyl glycine are full agonists at GPR18 and cause migration in the human endometrial cell line, HEC-1B. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [Green Version]
- Lu, V.B.; Puhl, H.L., 3rd; Ikeda, S.R. N-Arachidonyl glycine does not activate G protein-coupled receptor 18 signaling via canonical pathways. Mol. Pharmacol. 2013, 83, 267–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, D.B.; Joseph, W.R.; Grimsey, N.L.; Glass, M. GPR18 undergoes a high degree of constitutive trafficking but is unresponsive to N-Arachidonoyl Glycine. PeerJ 2016, 4, e1835. [Google Scholar] [CrossRef] [Green Version]
- Schoeder, C.T.; Mahardhika, A.B.; Drabczyńska, A.; Kieć-Kononowicz, K.; Müller, C.E. Discovery of Tricyclic Xanthines as Agonists of the Cannabinoid-Activated Orphan G-Protein-Coupled Receptor GPR18. ACS Med. Chem. Lett. 2020, 11, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Kimura, S.; Toshida, T.; Ota, R.; Yamashita, A.; Sugiura, T. Lysophosphati- dylinositol induces rapid phosphorylation of p38 mitogen-activated protein kinase and activating tran- scription factor 2 in HEK293 cells expressing GPR55 and IM-9 lymphoblastoid cells. J. Biochem. 2010, 147, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Overton, H.A.; Babbs, A.J.; Doel, S.M.; Fyfe, M.C.; Gardner, L.S.; Griffin, G.; Jackson, H.C.; Procter, M.J.; Rasamison, C.M.; Tang-Christensen, M.; et al. Deorphanization of a G protein- coupled receptor for oleoylethanolamide and its use in the discovery of small-molecule hypophagic agents. Cell Metab. 2006, 3, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Silvestri, C.; Di Marzo, V. The endocannabinoid systém in energy homeostasis and the etiopathology of metabolic disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Silvestri, C.; Ligresti, A.; Di Marzo, V. Peripheral effects of endocannabinoid system in energy homeostasis: Adipose tissue, liver and skeletal muscle. Rev. Endocr. Metab. Disord. 2011, 12, 153–162. [Google Scholar] [CrossRef]
- Koch, M. Cannabinoid receptor signalling in cetral regulation of feeding behaviour: A mini-rewiew. Front. Neurosci. 2017, 11, 293. [Google Scholar] [CrossRef]
- Rempel, V.; Atzler, K.; Behrenswerth, A.; Karcz, T.; Schoeder, C.; Hinz, S.; Kaleta, M.; Thimm, D.; Kieć-Kononowicz, K.; Müller, C.E. Bicyclic imidazole-4-one derivatives: A new class of antagonists for the orphan G Protein-coupled receptors GPR18 and GPR55. Med. Chem. Comm. 2014, 5, 632–649. [Google Scholar] [CrossRef]
- Schoeder, C.T.; Kaleta, M.; Mahardhika, A.B.; Olejarz-Maciej, A.; Łażewska, D.; Kieć-Kononowicz, K.; Müller, C.E. Structure activity relationships of imidazothazinones and analogs as antagosnits of the cannabinoid-activated orphan G protein-coupled receptor GPR18. Eur. J. Med. Chem. 2018, 155, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.; Engel, V.; Mahardhika, A.B.; Schoeder, C.T.; Namasivayam, V.; Kieć-Kononowicz, K.; Müller, C.E. Computational Investigations on the Binding Mode of Ligands for the Cannabinoid-Activated G Protein-Coupled Receptor GPR18. Biomolecules 2020, 10, 686. [Google Scholar] [CrossRef] [PubMed]
- Simcocks, A.C.; Jenkin, K.A.; O’Keefe, L.; Samuel, C.S.; Mathai, M.L.; McAinch, A.J.; Hryciw, D.H. Atypical cannabinoid ligands O-1602 and O-1918 administered chronically in diet-induced obesity. Endocr. Connect. 2019, 8, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotańska, M.; Śniecikowska, J.; Jastrzębska-Więsek, M.; Kołaczkowski, M.; Pytka, K. Metabolic and Cardiovascular Benefits and Risks of EMD386088-A 5-HT6 Receptor Partial Agonist and Dopamine Transporter Inhibitor. Front. Neurosci. 2017, 11, 50. [Google Scholar] [CrossRef]
- Kotańska, M.; Lustyk, K.; Bucki, A.; Marcinkowska, M.; Śniecikowska, J.; Kołaczkowski, M. Idalopirdine, a selective 5-HT6 receptor antagonist, reduces food intake and body weight in a model of excessive eating. Metab. Brain Dis. 2018, 33, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepańska, K.; Pockes, S.; Podlewska, S.; Höring, C.; Mika, K.; Latacz, G.; Bednarski, M.; Siwek, A.; Karcz, T.; Nagl, M.; et al. Structural modifications in the distal, regulatory region of histamine H3 receptor antagonists leading to the identification of a potent anti-obesity agent. Eur. J. Med. Chem. 2020, 24, 113041. [Google Scholar] [CrossRef]
- Pagano, C.; Pilon, C.; Calcagno, A.; Urbanet, R.; Rossato, M.; Milan, G.; Bianchi, K.; Rizzuto, R.; Bernante, P.; Federspil, G.; et al. The endogenous cannabinoid system stimulates glucose up-take in human fat cells via phosphatidylinositol 3-kinase and calcium-dependent mechanisms. J. Clin. Endocrinol. Metab. 2007, 92, 4810–4819. [Google Scholar] [CrossRef] [Green Version]
- You, T.; Disanzo, B.L.; Wang, X.; Yang, R.; Gong, D. Adipose tissue endocannabinoid system gene expression: Depot differences and effects of diet and exercise. Lipids Health Dis. 2011, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Izzo, A.; Piscitelli, F.; Capasso, R.; Aviello, G. Peripheral endocannabinoid dysregulation in obesity: Relation to intestinal motility and energy processing induced by food deprivation and re-feeding. Br. J. Pharmacol. 2009, 58, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Kurczab, R.; Ali, W.; Łażewska, D.; Kotańska, M.; Jastrzębska-Więsek, M.; Satała, G.; Lubelska, A.; Latacz, G.; Partyka, A.; Starek, M.; et al. Computer-aided studies for novel arylhydantoin 1,3,5-triazine derivatives as 5-HT6 serotonin receptor ligands with antidepressive-like, anxiolytic and antiobesity action in vivo. Molecules 2018, 23, 2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Hyiaman, D.; Harvey-White, J.; Batkai, S.; Kunos, G. The role of the endocannabinoid system in the control of energy homeostasis. Int. J. Obes. 2006, 30 (Suppl. l1), 110–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkham, T.C.; Williams, C.M.; Fezza, F.; Di Marzo, V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satation: Stimulation of eating by 2-arachidonoyl glycerol. Br. J. Pharmacol. 2002, 136, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Matias, I. Endocannabinoid control of food intake and energy balance. Nat. Neurosci. 2005, 8, 585–589. [Google Scholar] [CrossRef]
- Engeli, S.; Böhnke, J.; Feldpausch, M.; Gorzelniak, K.; Janke, J.; Bátkai, S.; Pacher, P.; Harvey-White, J.; Luft, F.C.; Sharma, A.M.; et al. Activation of the peripheral endocannabinoid system in human obesity. Diabetes 2005, 54, 2838–2843. [Google Scholar] [CrossRef] [Green Version]
- Monteleone, P.; Matias, I.; Martiadis, V.; De Petrocellis, L.; Maj, M.; Di Marzo, V. Blood levels of the endocannabinoid anandamide are increased in anorexia nervosa and in binge-eating disorder, but not in bulimia nervosa. Neuropsychopharmacology 2005, 30, 1216–1221. [Google Scholar] [CrossRef] [Green Version]
- Dudek, M.; Marcinkowska, M.; Bucki, A.; Olczyk, A.; Kołaczkowski, M. Idalopirdine—A small molecule antagonist of 5-HT6 with therapeutic potential against obesity. Metab. Brain Dis. 2015, 30, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Vickers, S.P.; Jackson, H.C.; Cheetham, S.C. The utiliry of animal models to evaluate novel anti-obesity agents. Br. J. Pharmacol. 2011, 164, 1248–1262. [Google Scholar] [CrossRef] [Green Version]
- Takeda, N.; Hasegawa, S.; Morita, M.; Matsunaga, T. Pica in rats is analogous to emesis: An animal model in emesis research. Pharmacol. Biochem. Behav. 1993, 45, 817–821. [Google Scholar] [CrossRef]
- Malmlöf, K.; Zaragoza, F.; Golozoubova, V.; Refsgaard, H.H.F.; Cremers, T.; Raun, K.; Wulff, B.S.; Johansen, P.B.; Westerink, B.; Rimvall, K. Influence of a selective histamine H3 receptor antagonist on hypothalamic neural activity, food intake and body weight. Int. J. Obes. 2005, 29, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Bluher, M.; Michael, M.D.; Peroni, O.D.; Ueki, K.; Carter, N.; Kahn, B.B.; Kahn, C.R. Adipose tissue selective insulin receptor knockout protects against obesity and obesity-releted glucose intolerance. Dev. Cell 2002, 3, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Arteaga, A.; Vaźquez, M.; Vazquez-Martinez, P.M.R.; Suarez, J.; Velásquez, D.A.; López, M.; Ross, R.A.; Rodriguez de Fonseca, F.; Bermudez-Silva, F.J.; Malagón, M.M.; et al. The atypical cannabinoid O-1602 stimulates food intake and adiposity in rats. Diabetes Obes. Metab. 2012, 14, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Iguchi, H.; Nakata, M.; Ioka, R.X.; Tanaka, T.; Iwasaki, S.; Magoori, K.; Takayasu, S.; Yamamoto, T.T.; Kodama, T.; et al. Identification of N-arachidonylglycine, U18666A, and 4-androstene- 3,17-dione as novel insulin Secretagogues. Biochem. Biophys. Res. Commun. 2005, 333, 778–786. [Google Scholar] [CrossRef]
- Mohan, H.; Brandt, S.L.; Kim, J.H.; Wong, F.; Lai, M.; Prentice, K.J.; Rijjal, D.A.; Magomedova, L.; Batchuluun, B.; Burdett, E.; et al. 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF)prevents high fat diet-induced insulin resistance via maintenance of hepatic lipid homeostasis. Diabetes Obes. Metab. 2019, 21, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Mofrad, P.; Contos, M.J.; Haque, M.; Sargeant, C.; Fisher, R.A.; Luketic, V.A.; Sterling, R.K.; Shiffman, M.L.; Stravitz, R.T.; Sanyal, A.J. Clinical and histologic spectrumof nonalcoholic fatty liver disease associated with normal ALT values. Hepatology 2003, 37, 1286–1292. [Google Scholar] [CrossRef]
- Dudek, M.; Kuder, K.; Kołaczkowski, M.; Olczyk, A.; Żmudzka, E.; Rak, A.; Bednarski, M.; Pytka, K.; Sapa, J.; Kieć-Kononowicz, K. H3 histamine receptor antagonist pitolisant reverses some subchronic disturbances induced by olanzapine in mice. Metab. Brain Dis. 2016, 31, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Matsunaga, S.; Matsui, M.; Takeda, N.; Yamatodani, A. Pica in mice as a new model for the study of emesis. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 135–138. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotańska, M.; Mika, K.; Szafarz, M.; Kubacka, M.; Müller, C.E.; Sapa, J.; Kieć-Kononowicz, K. Effects of GPR18 Ligands on Body Weight and Metabolic Parameters in a Female Rat Model of Excessive Eating. Pharmaceuticals 2021, 14, 270. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030270
Kotańska M, Mika K, Szafarz M, Kubacka M, Müller CE, Sapa J, Kieć-Kononowicz K. Effects of GPR18 Ligands on Body Weight and Metabolic Parameters in a Female Rat Model of Excessive Eating. Pharmaceuticals. 2021; 14(3):270. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030270
Chicago/Turabian StyleKotańska, Magdalena, Kamil Mika, Małgorzata Szafarz, Monika Kubacka, Christa E. Müller, Jacek Sapa, and Katarzyna Kieć-Kononowicz. 2021. "Effects of GPR18 Ligands on Body Weight and Metabolic Parameters in a Female Rat Model of Excessive Eating" Pharmaceuticals 14, no. 3: 270. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030270