
Chicory Extracts and Sesquiterpene Lactones Show Potent Activity against Bacterial and Fungal Pathogens

 , , , , , and
, , , , , and 
Abstract
:
1. Introduction
2. Results
2.1. Composition of the Extracts
2.2. Antibacterial Activity
2.3. Antifungal Activity
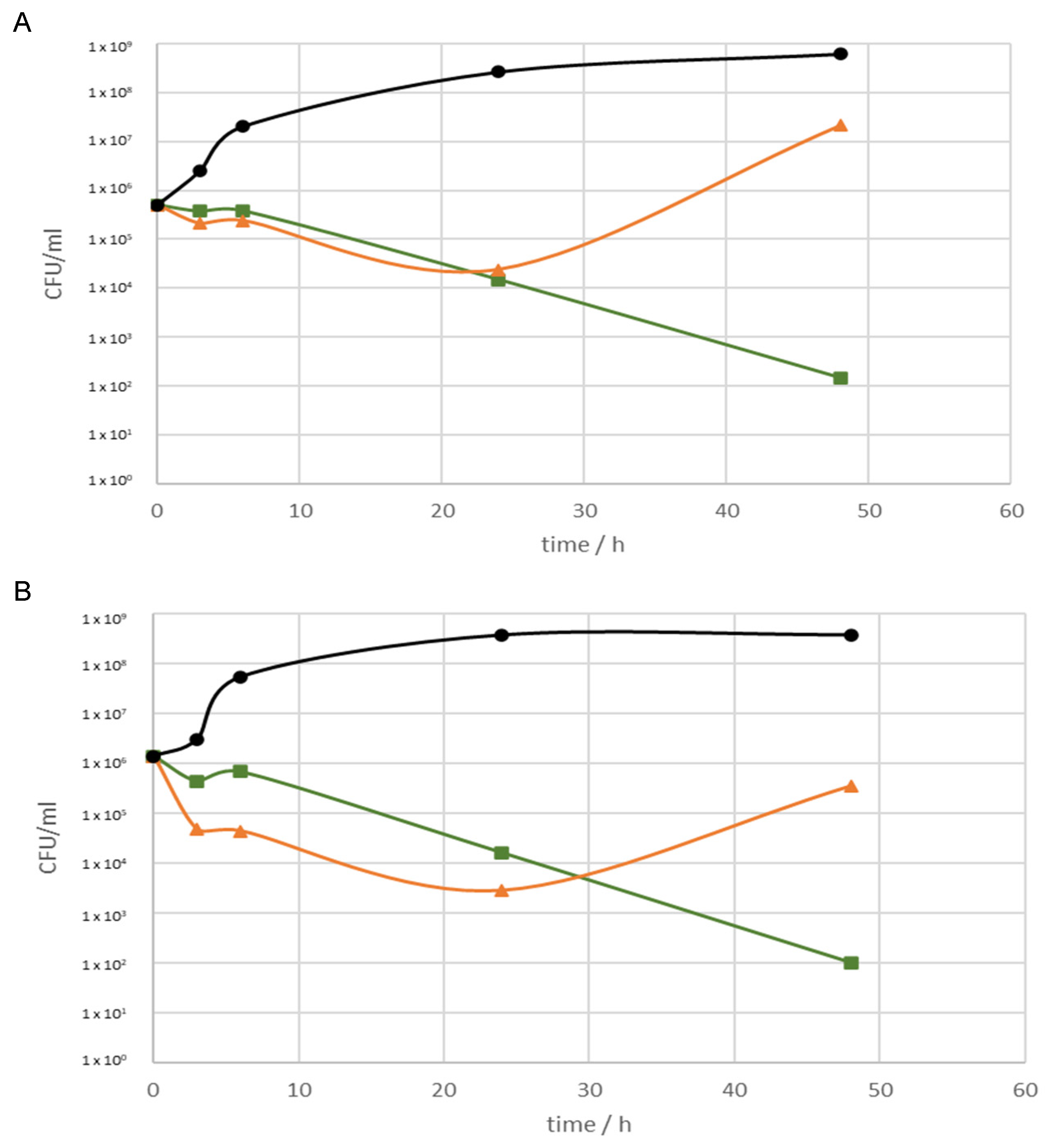
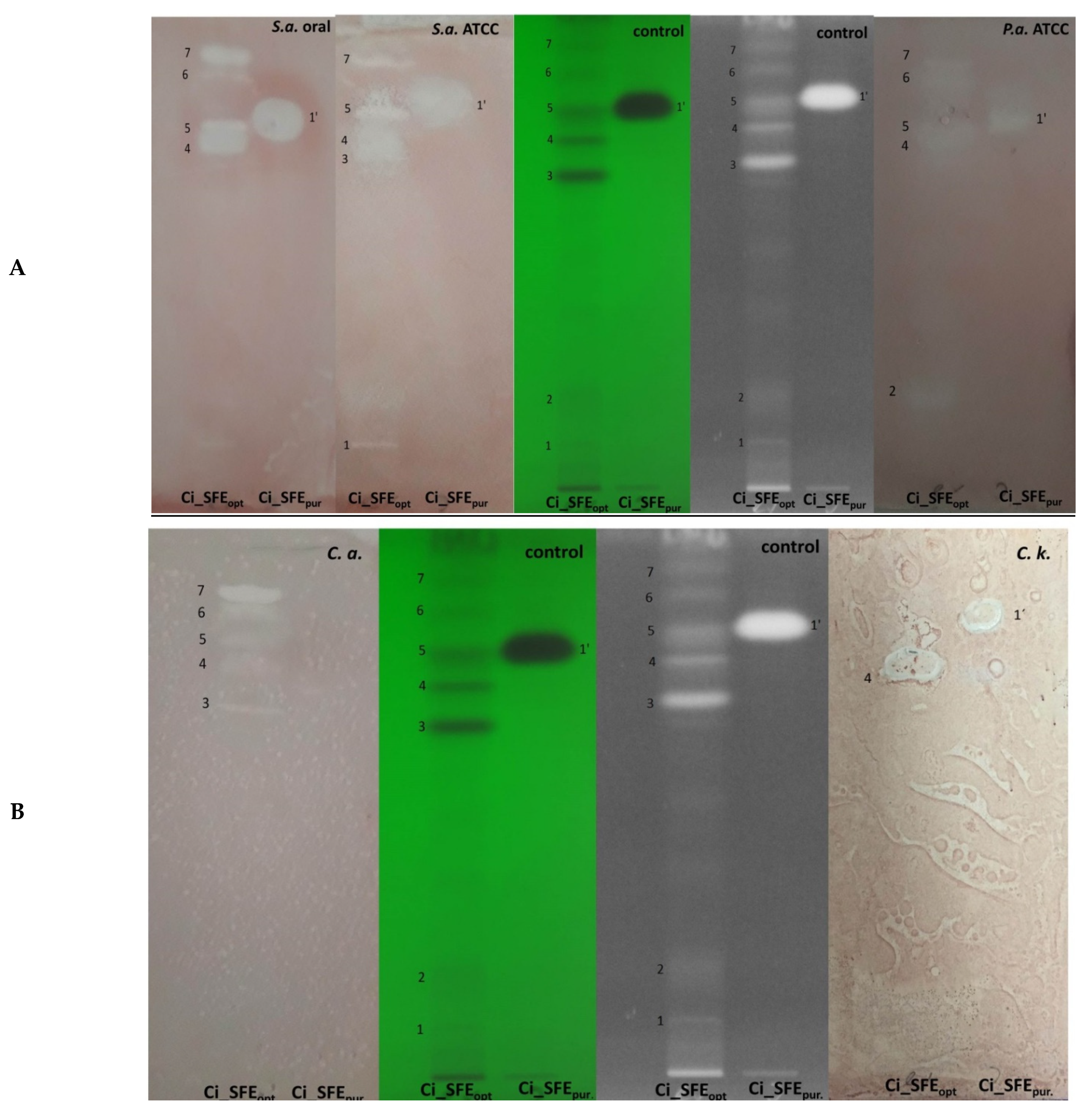
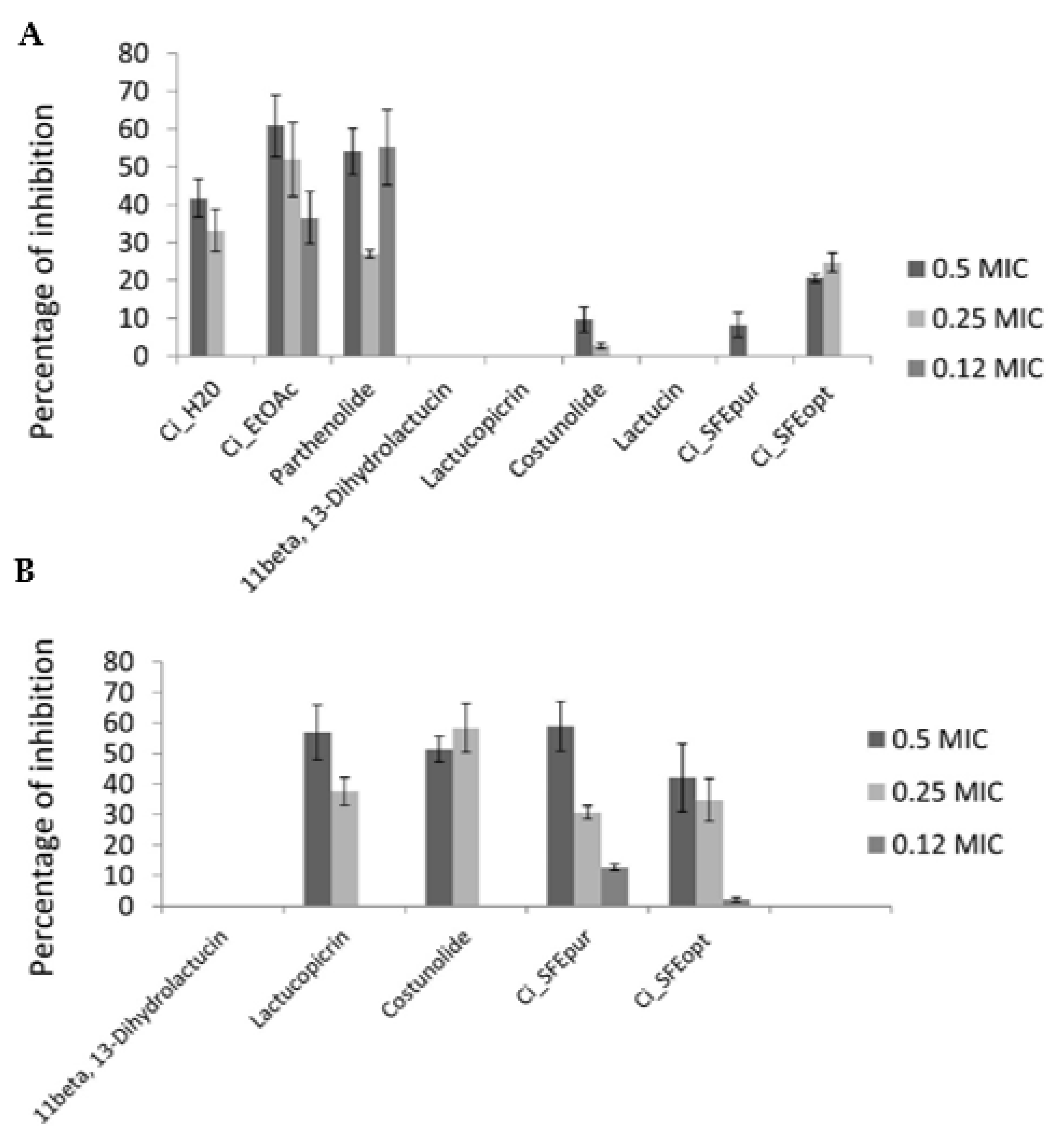
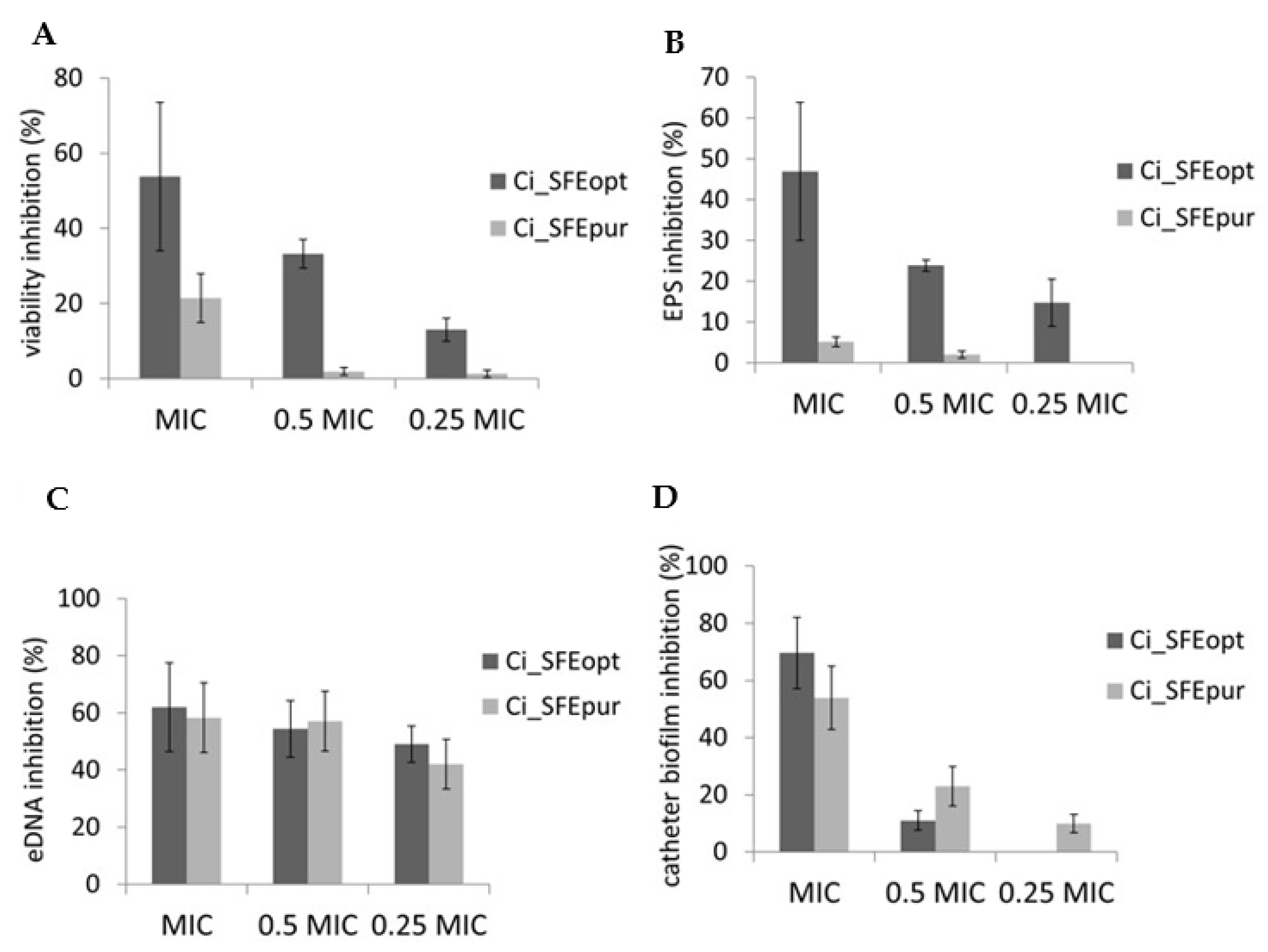
2.4. Antibiofilm Activity
2.5. Inhibition of Pyocyanin Production by Pseudomonas aeruginosa IBRS P001
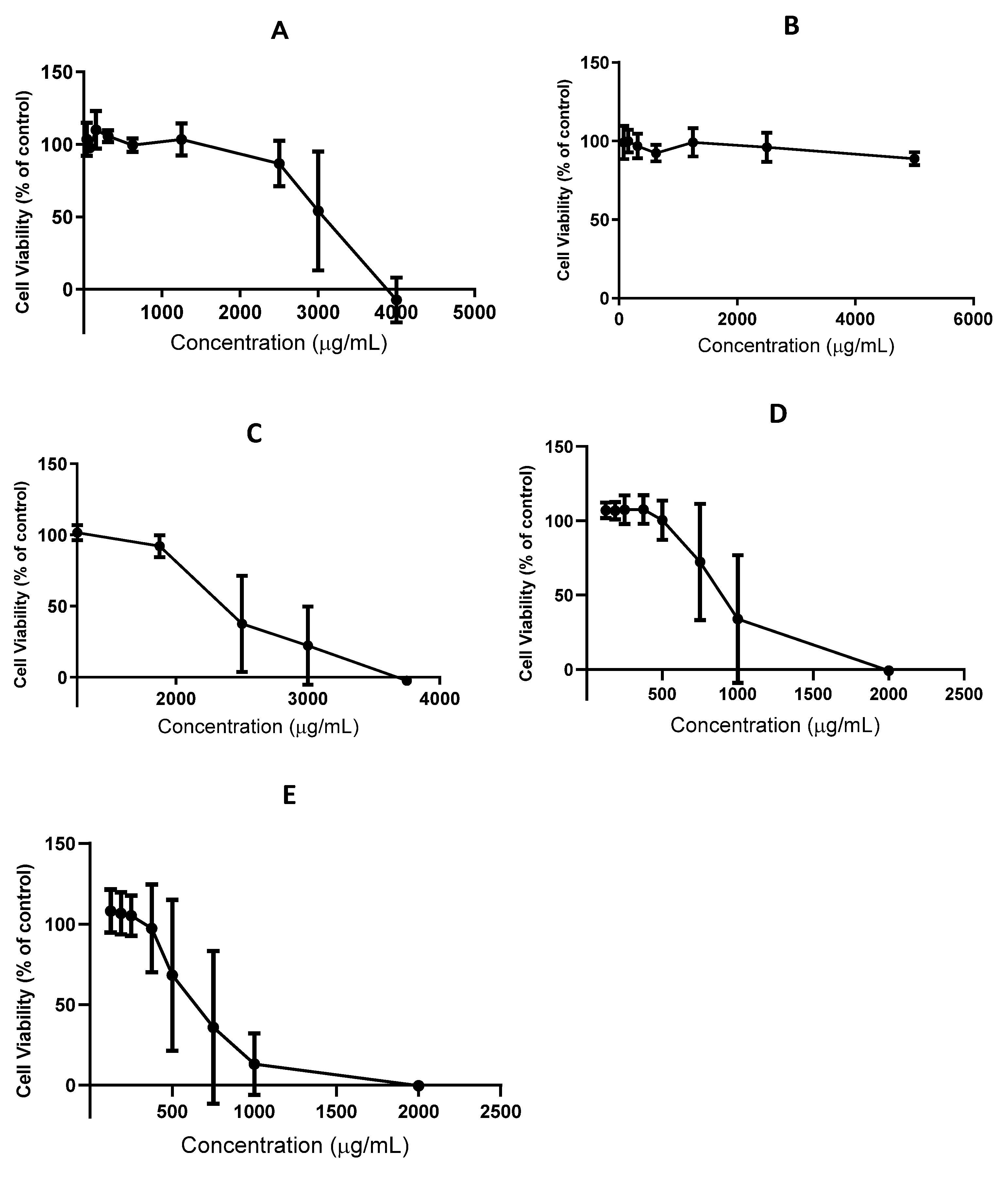
2.6. Toxicity
3. Discussion
4. Materials and Methods
4.1. Sample Extraction
4.2. Identification of STLs
4.3. Antimicrobial Activity
4.4. Antifungal Activity
4.5. Antibiofilm Activity
4.5.1. Calculation of Percentage Inhibition
4.5.2. Biofilm Cell Viability MTT Assay
4.5.3. Congo Red EPS-Binding Assay
4.5.4. Quantification of eDNA
4.5.5. Catheter Model of Biofilm Formation
4.6. Inhibition of Pseudomonas aeruginosa IBRS P001 Pyocyanin Production
4.7. Toxicity
4.7.1. Photobacterial Acute Toxicity Assay
4.7.2. HaCaT Cell Antiproliferative Activity Assay
4.7.3. Caco-2 Cell Cytotoxicity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Arkel, J.; Sévenier, R.; Hakkert, J.C.; Bouwmeester, H.J.; Koops, A.J.; van der Meer, I.M. Tailor-Made Fructan Synthesis in Plants: A Review. Carbohydr. Polym. 2013, 93, 48–56. [Google Scholar] [CrossRef]
- Van Arkel, J. Fructan Biosynthesis in Crop Plants, The Molecular Regulation of Fructan Biosynthesis in Chicory (Cichorium Intybus L.). Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2013. [Google Scholar]
- Cankar, K.; Bundock, P.; Sevenier, R.; Häkkinen, S.T.; Hakkert, J.C.; Beekwilder, J.; Meer, I.M.; Both, M.; Bosch, D. Inactivation of the Germacrene A Synthase Genes by CRISPR/Cas9 Eliminates the Biosynthesis of Sesquiterpene Lactones in Cichorium Intybus L. Plant Biotechnol. J. 2021, 13670. [Google Scholar] [CrossRef]
- Street, R.A.; Sidana, J.; Prinsloo, G. Cichorium Intybus: Traditional Uses, Phytochemistry, Pharmacology, and Toxicology. Evid. Based Complement. Altern. Med. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Li-Weber, M.; Giaisi, M.; Treiber, M.; Krammer, P. The Anti-Inflammatory Sesquiterpene Lactone Parthenolide Suppresses IL-4 Gene Expression in Peripheral Blood T Cells. Eur. J. Immunol. 2002, 32, 3587–3597. [Google Scholar] [CrossRef]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Saliba, N.A.; Darwiche, N. What Made Sesquiterpene Lactones Reach Cancer Clinical Trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef]
- Spratt, D.A.; Daglia, M.; Papetti, A.; Stauder, M.; O’Donnell, D.; Ciric, L.; Tymon, A.; Repetto, B.; Signoretto, C.; Houri-Haddad, Y.; et al. Evaluation of Plant and Fungal Extracts for Their Potential Antigingivitis and Anticaries Activity. J. Biomed. Biotechnol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sessa, R.A.; Bennett, M.H.; Lewis, M.J.; Mansfield, J.W.; Beale, M.H. Metabolite Profiling of Sesquiterpene Lactones from Lactuca Species. J. Biol. Chem. 2000, 275, 26877–26884. [Google Scholar] [CrossRef]
- Bogdanović, M.; Cankar, K.; Todorović, S.; Dragicević, M.; Simonović, A.; van Houwelingen, A.; Schijlen, E.; Schipper, B.; Gagneul, D.; Hendriks, T.; et al. Tissue Specific Expression and Genomic Organization of Bitter Sesquiterpene Lactone Biosynthesis in Cichorium Intybus L. (Asteraceae). Ind. Crops Prod. 2019, 129, 253–260. [Google Scholar] [CrossRef]
- Pfaffenrath, V.; Diener, H.; Fischer, M.; Friede, M.; Henneicke-von Zepelin, H. The Efficacy and Safety of Tanacetum Parthenium (Feverfew) in Migraine Prophylaxis—A Double-Blind, Multicentre, Randomized Placebo-Controlled Dose-Response Study. Cephalalgia 2002, 22, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Baixinho, J.P.; Anastácio, J.D.; Ivasiv, V.; Cankar, K.; Bosch, D.; Menezes, R.; de Roode, M.; dos Santos, C.N.; Matias, A.A.; Fernández, N. Supercritical CO2 Extraction as a Tool to Isolate Anti-Inflammatory Sesquiterpene Lactones from Cichorium Intybus L. Roots. Molecules 2021, 26, 2583. [Google Scholar] [CrossRef] [PubMed]
- Grand View Research. Market Report. Available online: www.reportlinker.com/p05744655/Antibiotics-Market-Size-Share-Trends-Analysis-Report-By-Action-Mechanism-By-Drug-Class-And-Segment-Forecasts.html (accessed on 20 September 2021).
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- OECD. Antimicrobial Resistance in G7 Countries and Beyond: Economic Issues, Policies and Options for Action. Available online: www.oecd:els/health-systems/Antimicrobial-Resistance-in-G7-Countries-and-Beyond.pdf?_ga=2.113836423.1405534895.1623152607-1871292710.1623152607 (accessed on 20 September 2021).
- OECD. Briefing Note for EU/EEA Countries: Antimicrobial Resistance: Tackling the Burden in the European Union. Available online: www.oecd:els/health-systems/antimicrobial-resistance.htm (accessed on 20 September 2021).
- Cong, Y.; Yang, S.; Rao, X. Vancomycin Resistant Staphylococcus Aureus Infections: A Review of Case Updating and Clinical Features. J. Adv. Res. 2020, 21, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wingard, J.R.; Kubilis, P.; Lee, L.; Yee, G.; White, M.; Louise, W.; Bowden, R.; Anaissie, E.; Hiemenz, J.; Lister, J. Clinical Significance of Nephrotoxicity in Patients Treated with Amphotericin B for Suspected or Proven Aspergillosis. Clin. Infect. Dis. 1999, 29, 1402–1407. [Google Scholar] [CrossRef] [PubMed]
- Aleksić, M.; Stanisavljević, D.; Smiljković, M.; Vasiljević, P.; Stevanović, M.; Soković, M.; Stojković, D. Pyrimethanil: Between Efficient Fungicide against Aspergillus Rot on Cherry Tomato and Cytotoxic Agent on Human Cell Lines. Ann. Appl. Biol. 2019, 175, 228–235. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M.; Parker, J. Brock Biology of Micro-Organisms; Carlson, G., Ed.; Pearson Education Ltd.: London, UK, 2006. [Google Scholar]
- Petrović, J.; Stojković, D.; Soković, M. Terpene Core in Selected Aromatic and Edible Plants: Natural Health Improving Agents. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2019; pp. 423–451. [Google Scholar] [CrossRef]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Molecular Mechanisms in Candida Albicans and Beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef]
- Abbas, M.; Adil, M.; Ehtisham-ul-Haque, S.; Munir, B.; Yameen, M.; Ghaffar, A.; Shar, G.A.; Asif Tahir, M.; Iqbal, M. Vibrio Fischeri Bioluminescence Inhibition Assay for Ecotoxicity Assessment: A Review. Sci. Total Environ. 2018, 626, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from January 1981 to September 2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D.S. Plant Natural Products Targeting Bacterial Virulence Factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Lobato Gómez, M.; Huang, X.; Alvarez, D.; He, W.; Baysal, C. Contributions of the International Plant Science Community to the Fight against Human Infectious Diseases—Part 1: Epidemic and Pandemic Diseases. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef]
- Porras, G.; Chassagne, F.; Lyles, J.T.; Marquez, L.; Dettweiler, M.; Salam, A.M.; Samarakoon, T.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. Ethnobotany and the Role of Plant Natural Products in Antibiotic Drug Discovery. Chem. Rev. 2020, 121, 3495–3560. [Google Scholar] [CrossRef]
- Luepke, K.H.; Suda, K.J.; Boucher, H.; Russo, R.L.; Bonney, M.W.; Hunt, T.D.; Mohr, J.F. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacotherapy 2017, 37, 71–84. [Google Scholar] [CrossRef] [PubMed]
- The Pew Charitable Trusts. Antibiotics Currently in Clinical Development. Available online: http://www.pewtrusts:en/multimedia/data-visualizations/2014/antibiotics-currently-in-clinical-development (accessed on 26 February 2018).
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How Antibiotics Kill Bacteria: From Targets to Networks. In Nature Reviews Microbiology; NIH Public: Bethesda, MD, USA, 2010; pp. 423–435. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D. Opportunities for Natural Products in 21st Century Antibiotic Discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.; Del Fiol, F.S.; Balcão, V.M. Prospects for the Use of New Technologies to Combat Multidrug-Resistant Bacteria. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Revie, N.M.; Iyer, K.R.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Evolution, Mechanisms and Impact. Curr. Opin. Microbiol. 2018, 45, 70–76. [Google Scholar] [CrossRef]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida Auris Sp. Nov., a Novel Ascomycetous Yeast Isolated from the External Ear Canal of an Inpatient in a Japanese Hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Manuel, R.; Brown, C.S. Candida Auris: A Review of the Literature. Clin. Microbiol. Rev. 2017, 31. [Google Scholar] [CrossRef] [Green Version]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural Products as Inspiration for the Development of Bacterial Antibiofilm Agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- Puupponen-Pimia, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kahkonen, M.; Heinonen, M.; Maatta-Riihinen, K.; Oksman-Caldentey, K.-M. Berry Phenolics Selectively Inhibit the Growth of Intestinal Pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef]
- Tylewicz, U.; Oliveira, G.; Alminger, M.; Nohynek, L.; Dalla Rosa, M.; Romani, S. Antioxidant and Antimicrobial Properties of Organic Fruits Subjected to PEF-Assisted Osmotic Dehydration. Innov. Food Sci. Emerg. Technol. 2020, 62, 102341. [Google Scholar] [CrossRef]
- Laitila, A.; Alakomi, H.-L.; Raaska, L.; Mattila-Sandholm, T.; Haikara, A. Antifungal Activities of Two Lactobacillus Plantarum Strains against Fusarium Moulds in Vitro and in Malting of Barley. J. Appl. Microbiol. 2002, 93, 566–576. [Google Scholar] [CrossRef]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; Griensven, L.J.L.D. van. Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an In Vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, J.; Novakovic, M.; Tesevic, V.; Ciric, A.; Sokovic, M.; Zdunic, G.; Dajic-Stevanovic, Z.; Godjevac, D. HPTLC-Direct Bioautography-Guided Isolation of Isogeranic Acid as the Main Antibacterial Constituent of Artemisia Santonicum Essential Oil. J. Serbian Chem. Soc. 2019, 84, 1355–1365. [Google Scholar] [CrossRef]
- Rodríguez-Tudela, J.L.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Denning, D.; Donnelly, J.P.; Dupont, B.; Fegeler, W.; Moore, C.; et al. Method for the Determination of Minimum Inhibitory Concentration (MIC) by Broth Dilution of Fermentative Yeasts. Clin. Microbiol. Infect. 2003, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cady, N.C.; McKean, K.A.; Behnke, J.; Kubec, R.; Mosier, A.P.; Kasper, S.H.; Burz, D.S.; Musah, R.A. Inhibition of Biofilm Formation, Quorum Sensing and Infection in Pseudomonas Aeruginosa by Natural Products-Inspired Organosulfur Compounds. PLoS ONE 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartsev, V.; Lichitsky, B.; Geronikaki, A.; Petrou, A.; Smiljkovic, M.; Kostic, M.; Oliver, R.; Sokovic, M. Design, Synthesis and Antimicrobial Activity of Usnic Acid Derivatives. Med. Chem. Commun. 2018, 9, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Dey, P.; Parai, D.; Banerjee, M.; Hossain, S.T.; Mukherjee, S.K. Naringin Sensitizes the Antibiofilm Effect of Ciprofloxacin and Tetracycline against Pseudomonas Aeruginosa Biofilm. Int. J. Med. Microbiol. 2020, 310, 151410. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, A.; Valliammai, A.; Premika, M.; Priya, A.; Bhaskar, J.P.; Krishnan, V.; Pandian, S.K. Sapindus Mukorossi Gaertn. and Its Bioactive Metabolite Oleic Acid Impedes Methicillin-Resistant Staphylococcus Aureus Biofilm Formation by down Regulating Adhesion Genes Expression. Microbiol. Res. 2021, 242, 126601. [Google Scholar] [CrossRef]
- Matos, M.S.; Anastácio, J.D.; Allwood, J.W.; Carregosa, D.; Marques, D.; Sungurtas, J.; McDougall, G.J.; Menezes, R.; Matias, A.A.; Stewart, D.; et al. Assessing the Intestinal Permeability and Anti-Inflammatory Potential of Sesquiterpene Lactones from Chicory. Nutrients 2020, 12, 3547. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STL (µg/mg Extract) * | ||||||
|---|---|---|---|---|---|---|
| Code | Raw Material | Method | 11β,13-Dihydrolactucin | Lactucin | 11β,13-Dihydrolactucopicrin | Lactucopicrin |
| Wit_H2O | Freeze-dried witloof | Water extract (solid–liquid extraction, S-L) | 2.13 | 2.08 | 1.07 | 2.23 |
| Wit_EtOAc | Freeze-dried witloof | Ethyl acetate extract (S-L extraction) | 0.86 | 1.35 | 1.18 | 3.17 |
| Ci_H2O | Fresh chicory | Water extract (S-L extraction) | 0.48 | 1.13 | 0.39 | 1.76 |
| Ci_EtOAc | Fresh chicory | Ethyl acetate extract (S-L extraction) | 4.81 | 8.09 | 3.18 | 26.88 |
| Ci_SFE | Freeze-dried chicory | Supercritical fluid extraction (SFE) | 36.31 | 109.31 | 19.50 | 262.19 |
| Ci_SFEopt | Freeze-dried chicory | Optimized SFE | 170.19 | 257.59 | 55.62 | 271.04 |
| Ci_SFEpur | Freeze-dried chicory | SFE fraction purified by flash column chromatography ** | - | - | - | - |
| S. aureus VTT E-70045 | P. aeruginosa VTT E- 96728 | E. coli VTT E-94564T | |
|---|---|---|---|
| Standards | |||
| Lactucopicrin 4 mM (1.64 mg/mL) | ++ | + | + |
| Parthenolide 4 mM (1.00 mg/mL) | ++ | + | + |
| 11-β-Dihydrolactucopicrin 4 mM (1.65 mg/mL) | ++ | + | + |
| Lactucin 4 mM (1.11 mg/mL) | + | + | + |
| Extracts | |||
| Ci_EtOAc (0.05 mg/mL) | ++ | + | + |
| Ci_H2O (1.00 mg/mL) * | + | - | NA |
| Wit_EtOAc (1.00 mg/mL) | ++ | - | NA |
| Wit_H2O (1.00 mg/mL) | - | - | NA |
| Ci_SFE (1.00 mg/mL) | +++ | - | NA |
| S. aureus oral | S. aureus ATCC 11632 | P. aeruginosa ATCC 27853 | P. aeruginosa IBRS P001 | E. coli ATCC 25922 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Extracts | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC |
| Ci_EtOAc | 0.75 | 1.50 | 0.75 | 1.50 | 1.50 | 3.00 | - | - | 6.00 | 6.00 |
| Ci_H2O | - | - | - | - | - | - | - | - | - | - |
| Standards | ||||||||||
| Costunolide | - | - | - | - | - | - | 0.50 | 1.00 | - | - |
| Parthenolide | 0.08 | 0.16 | 0.16 | 0.31 | 0.31 | 0.63 | - | - | - | - |
| Lactucin | - | - | - | - | - | - | - | - | - | - |
| Lactucopicrin | - | - | 0.16 | 0.31 | 0.31 | 0.63 | 0.50 | 1.00 | - | - |
| 11β, 13-Dihydrolactucin | - | - | - | - | - | - | 0.50 | 1.00 | - | - |
| Controls | ||||||||||
| Ampicillin | 0.002 | 0.003 | 0.002 | 0.003 | 0.0002 | 0.0004 | - | - | 0.003 | 0.003 |
| Streptomycin | 0.006 | 0.012 | 0.050 | 0.100 | 0.0004 | 0.0008 | 0.050 | 0.100 | 0.030 | 0.030 |
| Strain | Ci_SFEpur | Ci_SFEopt | Ampicillin | |||
|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | |
| Proteus mirabilis ATCC 7002 | 0.25 | 0.50 | 2.10 | 4.40 | 0.01 | 0.01 |
| Listeria monocytogenes NCTC 7973 | 0.25 | 0.50 | 1.05 | 2.1 | 0.40 | 0.50 |
| Pseudomonas aeruginosa IBRS P001 | 0.06 | 0.12 | 0.50 | 1.00 | - | - |
| Enterobacter cloacae human isolate | 0.25 | 0.50 | 2.10 | 4.40 | 0.10 | 0.15 |
| Yersinia enterocolitica ATCC 23715 | 0.25 | 0.50 | 2.10 | 4.40 | 0.004 | 0.008 |
| Klebsiella pneumoniae ATCC 13883 | - | - | 0.50 | 1.00 | 0.20 | 0.40 |
| Campylobacter jejuni ATCC 33560 | 0.25 | 0.50 | 1.00 | 2.10 | 0.02 | 0.04 |
| (A) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. albicans 475/15 | C. albicans ATCC 10231 | C. krusei | C. auris | C. parapsilosis | ||||||||||
| MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | |||||
| Extracts | ||||||||||||||
| Ci_EtOAc | 1.00 | 2.00 | - | - | - | - | - | - | - | - | ||||
| Ci_H2O | 1.00 | 2.00 | - | - | - | - | - | - | - | - | ||||
| Ci_SFEpur | 0.06 | 0.12 | 0.06 | 0.12 | 0.06 | 0.12 | 0.03 | 0.06 | 0.25 | 0.50 | ||||
| Ci_SFEopt | 0.50 | 1.10 | 0.50 | 1.10 | 0.50 | 1.10 | 0.25 | 0.50 | 0.50 | 1.10 | ||||
| Standards | ||||||||||||||
| Costunolide | 0.50 | 1.00 | - | - | 0.13 | 0.25 | - | - | - | - | ||||
| Parthenolide | 0.25 | 0.50 | - | - | 0.03 | 0.06 | - | - | - | - | ||||
| Lactucin | 1.00 | >1.00 | - | - | 0.50 | 1.00 | - | - | - | - | ||||
| Lactucopicrin | 1.00 | >1.00 | - | - | 0.50 | 1.00 | - | - | - | - | ||||
| 11β, 13-Dihydrolactucin | 0.50 | 1.00 | - | - | 0.50 | 1.00 | 0.25 | 0.50 | 0.25 | 0.50 | ||||
| Control | ||||||||||||||
| Ketoconazole | 0.030 | 0.060 | 0.002 | 0.032 | 0.006 | 0.010 | - | - | 0.003 | 0.006 | ||||
| (B) | ||||||||||||||
| A. fumigatus | A. versicolor | A. ochraceus | A. niger | T. viride | P. ochrochloron | P. funiculosum | ||||||||
| MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | |
| Extracts | ||||||||||||||
| Ci_EtOAc | - | - | 3.00 | 4.00 | 4.00 | 8.00 | 6.00 | 8.00 | 1.00 | 2.00 | 0.50 | 1.00 | NA | NA |
| Ci_H2O | - | - | - | - | - | - | - | - | - | - | - | - | NA | NA |
| Ci_SFEpur | 0.13 | 0.26 | 0.03 | 0.06 | 0.25 | 0.50 | 0.50 | 1.00 | NA | NA | NA | NA | 0.25 | 0.50 |
| Ci_SFEopt | 1.10 | 2.20 | 0.50 | 1.10 | 1.10 | 2.20 | 2.20 | 4.40 | NA | NA | NA | NA | 1.10 | 2.20 |
| Standards | ||||||||||||||
| Costunolide | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Parthenolide | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lactucin | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lactucopicrin | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 11β, 13-Dihydrolactucin | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Control | ||||||||||||||
| Ketoconazole | 0.180 | 0.250 | 0.045 | 0.062 | 0.250 | 0.500 | 0.375 | 0.500 | 0.375 | 0.500 | 0.031 | 0.159 | 0.150 | 0.200 |
| Standards | Inhibition (%) | Concentration mg/mL |
|---|---|---|
| Costunolide | 55.25 | 0.0054 |
| Lactucin | 69.63 | 0.0036 |
| Lactucopicrin | 52.83 | 0.0057 |
| 11β-13-dihydrolactucin | 36.13 | 0.0076 |
| Control | 0 | 0.0120 |
| Sample Code | EC50, mg/L a | Toxicity Category (According to Directive 93/67/EEC) b | |
|---|---|---|---|
| 15 min | 30 min | ||
| Ci_EtOAc, in ethanol | 1133 | 739 | not harmful |
| Ci_EtOAc, in water | 1891 | 1415 | not harmful |
| Wit_EtOAc | 203 | 184 | not harmful |
| Wit_H2O | 5737 | 6385 | not harmful |
| Ci_SFE | 83 | 68 | harmful |
| Ci_SFEopt | 131 | 142 | not harmful |
| Ci_SFEpur | 3076 c | 1674 c | not harmful |
| IC50 (µg/mL) | |
|---|---|
| Ci_SFEopt | 303.4 ± 1.5 |
| Ci_SFEpur | 16.8 ± 0.1 |
| K2Cr2O7 | 0.9 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häkkinen, S.T.; Soković, M.; Nohynek, L.; Ćirić, A.; Ivanov, M.; Stojković, D.; Tsitko, I.; Matos, M.; Baixinho, J.P.; Ivasiv, V.; et al. Chicory Extracts and Sesquiterpene Lactones Show Potent Activity against Bacterial and Fungal Pathogens. Pharmaceuticals 2021, 14, 941. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14090941
Häkkinen ST, Soković M, Nohynek L, Ćirić A, Ivanov M, Stojković D, Tsitko I, Matos M, Baixinho JP, Ivasiv V, et al. Chicory Extracts and Sesquiterpene Lactones Show Potent Activity against Bacterial and Fungal Pathogens. Pharmaceuticals. 2021; 14(9):941. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14090941
Chicago/Turabian StyleHäkkinen, Suvi T., Marina Soković, Liisa Nohynek, Ana Ćirić, Marija Ivanov, Dejan Stojković, Irina Tsitko, Melanie Matos, João P. Baixinho, Viktoriya Ivasiv, and et al. 2021. "Chicory Extracts and Sesquiterpene Lactones Show Potent Activity against Bacterial and Fungal Pathogens" Pharmaceuticals 14, no. 9: 941. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14090941