
Plant Terpenoids as Hit Compounds against Trypanosomiasis
by
 , and
, and
Raquel Durão
1,†, 
Cátia Ramalhete
1,2,† ,
,
Ana Margarida Madureira
1,
Eduarda Mendes
1 and
Noélia Duarte
1,* 1
Research Institute for Medicines (iMED.Ulisboa), Faculdade de Farmácia, Universidade de Lisboa, Av. Prof. Gama Pinto, 1649-003 Lisboa, Portugal
2
ATLANTICA—Instituto Universitário, Fábrica da Pólvora de Barcarena, 2730-036 Barcarena, Portugal
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pharmaceuticals 2022, 15(3), 340; https://0-doi-org.brum.beds.ac.uk/10.3390/ph15030340
Submission received: 27 January 2022
/
Revised: 22 February 2022
/
Accepted: 4 March 2022
/
Published: 10 March 2022
(This article belongs to the Special Issue Plant and Marine-Derived Natural Product Research in Drug Discovery: Strengths and Perspective)
Abstract
:Human African trypanosomiasis (sleeping sickness) and American trypanosomiasis (Chagas disease) are vector-borne neglected tropical diseases, caused by the protozoan parasites Trypanosoma brucei and Trypanosoma cruzi, respectively. These diseases were circumscribed to South American and African countries in the past. However, human migration, military interventions, and climate changes have had an important effect on their worldwide propagation, particularly Chagas disease. Currently, the treatment of trypanosomiasis is not ideal, becoming a challenge in poor populations with limited resources. Exploring natural products from higher plants remains a valuable approach to find new hits and enlarge the pipeline of new drugs against protozoal human infections. This review covers the recent studies (2016–2021) on plant terpenoids, and their semi-synthetic derivatives, which have shown promising in vitro and in vivo activities against Trypanosoma parasites.
1. Introduction
Human African trypanosomiasis (sleeping sickness) and American trypanosomiasis (Chagas disease) are among the twenty Neglected Tropical Diseases (NTDs) defined as such by the World Health Organization (WHO). NTDs are a heterogeneous group of diseases including, among others, several parasitic, viral, and bacterial infections, responsible for high morbidity and mortality, and affecting more than one billion people globally [1,2,3]. Sometimes, the impact of NTDs on health can be underestimated because many infections are asymptomatic and associated with long incubation periods. Nevertheless, NTDs are recognized as a public health problem, particularly for people living in rural and conflict areas in developing countries [1]. These diseases are considered neglected due to the general lack of attention in developed countries and almost non-existent financial investment in the research and development of new drugs and vaccines [4]. In addition to all these problems, the COVID-19 pandemic has been affecting the programs of mass drug administration and other NTD control measures [5,6]. Currently, the pharmacological therapy of NTDs is not ideal, as it has some limitations that include severe side effects, unfavorable toxicity profiles, prolonged treatment duration, difficult administration procedures, and development of drug resistance [7,8,9]. The investment in these therapeutic areas by large pharmaceutical companies is not financially attractive due to the poor prospect of financial returns. Thus, the research of drugs against these diseases is not motivated by commercial reasons. Additionally, many pharmaceutical companies take an opportunistic approach to drug repositioning, using drugs that were previously developed and registered for other therapeutic indications and applying them in the treatment of NTDs. This strategy has obvious advantages, namely reduced development costs. However, the major disadvantage is the non-introduction of new specific drugs used in the treatment of these diseases [7].
Therefore, the discovery and development of new drugs is essential and urgent and should embrace the development of new therapeutic classes, the reduction in toxicity in the host, better administration processes, and the development of combined therapies [10]. One of the main strategies includes the phytochemical study of plants and other natural sources (marine organisms, animals, microorganisms, and fungi). Natural products have played an important role in the drug discovery and development processes [11]. However, in recent decades, most pharmaceutical companies have reduced their drug discovery and development programs from natural sources, largely due to the development of combinatorial chemistry programs [7]. Nevertheless, despite the large number of drugs derived from total synthesis, natural products and/or synthetic derivatives using their novel structures, contribute to the global number of new chemical entities that continue to be introduced on the market [12].
Over the last decade, several reviews have reported the bioactivity of natural products against protozoan neglected diseases [7,13,14,15,16,17,18,19,20,21]. However, natural products described in these reviews were obtained from different sources, including microbial [19], endophytes [20] and other fungi [21], marine [22], or animal origins [23,24]. Some reviews also present a mix of natural compounds origins [7,17]. Regarding natural products from plants, since 2016, there have been some reviews focusing exclusively on compounds from higher plants [16,18,25,26,27,28,29,30]. Nevertheless, the information is scattered amongst the diverse antiprotozoal diseases, compound families, and sources. To the best of our knowledge, a comprehensive review gathering the data concerning the most recent studies on terpenic compounds with antitrypanosomal activities is still missing. Therefore, in this work, a compilation of terpenes obtained from plants and evaluated for their activity against T. brucei and T. cruzi, covering the period from 2016 to 2021, will be presented and discussed. When available, data regarding in vivo activity and considerations about possible mechanisms of action will be also addressed.
The literature search was performed from June to December 2021 using Web of Science, ScienceDirect, PubMed, and some official websites (WHO, DNDi, CDC). An appropriate combination of keywords and truncation was selected and adapted for each database (for example, combinations of terpenoids or terpenes with Trypanosoma, Human African trypanosomiasis, and Chagas disease). Only peer-reviewed research articles or reviews in a six-year timespan (2016–2021) and in English language were considered. No restriction geographical origin of authors was applied. In particular cases, important reviews older than six years were also included. The literature was individually screened, applying as exclusion criteria, poor quality, inaccurate data, not considered relevant to the aim of the review, and articles reporting antitrypanosomal activity of extracts. Mendeley Reference Manager Software (2020) was used to manage the references and eliminate duplicates.
2. Trypanosomiasis
2.1. Human African Trypanosomiasis (HAT)
HAT is endemic in 36 African countries, with approximately 60 million people at risk, and approximately 10.8 million people living in areas of moderate to high risk of infection. In 1995, about 25,000 cases were detected and about 300,000 cases remained undetected. However, in 2001, the WHO launched an initiative to strengthen control and surveillance, and HAT declined in the following years. In 2019, less than 1000 cases were reported. It is noteworthy that this reduction is not due to a lack of control efforts as active and passive screening have been maintained at similar levels (about 2.5 million people screened per year) [31]. HAT is essentially present in poorer and rural areas, affecting populations dedicated to agriculture, fishing, livestock, and hunting, who are more exposed to the vector of transmission. Furthermore, their access to adequate health services is limited, thus lacking medical surveillance, also associated with difficulties in diagnosis and treatment [32]. This parasitic disease is transmitted mostly by the bite of the tsetse fly (Glossina palpalis), but other routes of transmission are possible, such as congenital transmission, blood transfusion, and transplants, despite being poorly documented [33]. The life cycle of Trypanosoma brucei sp. is illustrated in Figure 1 and reviewed elsewhere [10,34].
There are two subspecies of Trypanosoma brucei (T. b.), T. b. gambiense, and T. b. rhodesiense, with different geographic distributions. T. b. gambiense is found in 24 countries in West and Central Africa, accounting for more than 98% of reported cases. T. b. rhodesiense is present in 13 countries in East Africa, representing less than 3% of reported cases [32]. The two subspecies have different rates of progression and clinical characteristics. Infections by T. b. gambiense are characterized by a low parasitemia, with slow progression leading to the development of the chronic form of the disease, while T. b. rhodesiense progresses rapidly with high parasitemia, being characterized by the acute form. Both infections, if not diagnosed and treated, lead to death [34]. The clinical evolution of HAT has two phases. In the first phase, also known as the hemolymphatic phase, the parasite is found in the host’s blood and lymphatic stream. This initial phase includes non-specific symptoms such as pyrexia, headache, muscle and joint pain, weight loss and even enlarged lymph nodes, usually in the neck area. The second or neurologic phase (meningoencephalitis) occurs when the parasite crosses the blood–brain barrier and reaches the central nervous system. The clinical manifestations are usually behavioral changes, such as anxiety and irritability, sensory, motor, and sleep cycle disorders [32,33].
2.1.1. Antitrypanosomal Chemotherapy Targets and Current Drugs against HAT
Trypanosome-specific metabolic and cellular pathways represent excellent molecular targets. The ability to synthesize polyamines, putrescine, and spermidine, is of vital importance for the proliferation of bloodstream forms in trypanosomes. In this process, ornithine decarboxylase has a crucial function. This enzyme is considered the best-validated drug target in T. brucei, which is the target of eflornithine, a drug that is used clinically for the treatment of HAT [10,35]. In addition, the enzyme N-myristoyltransferase (NMT) has been well validated as a molecular target for HAT since its inhibition may lead to the death of the parasites. NMT catalyzes the covalent attachment of myristate, a 14-carbon saturated fatty acid, via amide bond to the N-terminal glycine residue of several proteins. NMT is also present in humans, but T. brucei is extremely sensitive to NMT inhibition, probably because endocytosis occurs at a very high rate in T. brucei [36]. Recently, significant progress in targeting the ubiquitin-proteasome system was reported [37]. The ubiquitin-proteasome system (UPS) is a crucial protein degradation system in eukaryotes and is essential for the survival of eukaryotes including trypanosomatids. There are promising inhibitors of this, but the overall success of clinical trials is low and therefore more drug candidates are needed. The bloodstream forms of T. brucei produce energy exclusively through glycolysis. Thus, inhibition of glycolytic enzymes, such as glyceraldehyde 3-phosphate dehydrogenase, phosphoglycerate mutase, phosphofructokinase and pyruvate kinase, could be a potential therapeutic approach. However, there is little prospect of killing trypanosomes by suppressing glycolysis unless inhibition is irreversible or uncompetitive, owing to the enormous glycolytic flux through the system [10,35]. Regarding redox metabolism, a fundamental metabolic difference between host and parasite is the existence of trypanothione reductase in trypanosomes instead of glutathione reductase, which is essential for the parasite’s survival. The inhibition of trypanothione reductase compromises the parasite’s oxidative defenses, sometimes leading to its death. Unfortunately, until now, compounds suitable for clinical development have not been discovered [10].
Currently, there is no vaccination or chemoprophylaxis for HAT, and its combat is mainly conducted through prophylactic measures aimed at reducing the reservoir of the disease and controlling the vector. The latter is the main strategy in use, which aims to minimize human contact with the fly. The recommended measures in the most affected areas are the use of clothes with neutral colors, and the use of insecticide repellents [34]. Recently, rapid diagnostic tests have been also developed in order to detect the presence of the antigen, offering accurate and sensitive results [38]. The treatment of HAT depends essentially on the stage of the disease and the causative agent. Until recently, five drugs have been used to treat sleeping sickness, donated by manufacturers to WHO for free distribution [38]. For the first phase, suramin (Naganinum®, Naganol®) and pentamidine (Nebupent®, Pentam®) are the first line drugs. For the final stage of sleeping sickness, the treatment includes the use of melarsoprol (Arsobal®), eflornithine (Vaniqa®) and the nifurtimox (Lampit®) -eflornithine combination therapy (NECT) [39]. Recently, fexinidazole was approved by the European Medicine Agency (EMA) and the United States Food and Drug Administration (USFDA) as the first all-oral therapy for the treatment of phase-1 and phase-2 HAT [39]. Additionally, acoziborole a recently developed benzoxaborole, is currently in advanced clinical trials, for treatment of phase-1 and phase-2 caused by both T.b. gambiense and T.b. rhodesiense. Acoziborole is orally bioavailable, and importantly, curative with one dose [40].
2.2. Human American Trypanosomiasis (Chagas Disease, CD)
Human American trypanosomiasis, commonly known as Chagas disease (CD), is endemic in 21 countries in Latin America. However, human migrations have turned it into a global disease with a significant number of cases in non-endemic regions such as Canada or Europe, among others. CD is present in rural areas and affects populations living in poverty. The WHO estimates that about 6 to 7 million people worldwide are infected and there are approximately 70 million people at risk [41,42]. CD is caused by the protozoan parasite Trypanosoma cruzi (T. cruzi), and it is generally transmitted by vectors, such as the triatomine hematophagous insects of the Reduviidae family, usually known as “barbers”. The other transmission routes are blood transfusions, transplantation, congenital transmission, and oral transmission (breast milk), and also by ingestion of contaminated food. The life cycle of Trypanosoma cruzi is illustrated in Figure 2 and reviewed elsewhere [41,42,43].
CD has two successive clinical phases, an acute phase, and a chronic phase. The acute phase can be symptomatic or, most frequently asymptomatic. The initial acute phase occurs immediately after the infection, which can last for weeks or months. It is characterized by local manifestations such as the Romaña Sign, when the parasite penetrates the conjunctiva, or the skin, causing a skin lesion or a purplish swelling of the lids of one eye called Chagoma. After a period of 4 to 8 weeks, the parasitemia decreases and the clinical manifestations spontaneously disappear in 90% of the cases, when the disease enters the chronic phase [41,43]. During the chronic phase, the parasites are in the heart and gastrointestinal tract. Despite the long-lasting nature of the infection in which most individuals do not develop overt pathology, there are about 30% of people who can achieve the chronic phase, characterized by progressive heart and/or digestive disease. In most of these cases, it takes decades to become apparent. Cardiomyopathy is the most serious result of T. cruzi infection, and in many areas of South America, it is a major cause of heart disease. Digestive symptoms, including megaesophagus and megacolon, also have serious consequences and may require surgery [43,44,45,46,47]
2.2.1. Antitrypanosomal Chemotherapy Targets and Current Drugs against CD
Infective trypomastigotes and intracellular replicative amastigotes are the clinically relevant life-cycle stages of T. cruzi that are potential targets for drug intervention [4,48]. T. cruzi requires specific sterols for cell viability and proliferation at all stages of the life cycle. The main sterol component of the parasite is ergosterol, while in the mammalian hosts it is cholesterol. Inhibitors of sterol biosynthesis have been shown antitrypanosomal in vitro activity [48]. Other trypanosomal targets are related to cysteine proteases that are involved in many crucial processes, including host cell invasion, cell division, and differentiation. T. cruzi contains a cysteine protease, cruzipain, which is responsible for proteolytic activity at all stages of the parasite’s life. Although no inhibitors of this family of enzymes have progressed to clinical trials, the parasite cysteine proteases remain a promising area of research [47,48]. In addition, the trypanothione reductases and synthetases have also been considered key enzymes in the oxidative metabolism of the parasite. Although several potential inhibitors of the trypanothione reductase possess potent in vitro anti-T. cruzi activity, to date, none have achieved parasitological cure in animal models [47].
Only two old nitroheterocyclic drugs, benznidazole (Rochagan® or Rodanil®) and nifurtimox (Lampit®) have been available for the treatment of CD, as reviewed elsewhere [41,49]. They are effective for the acute phase of infection, but they have variable efficacy in the chronic phase of the disease, besides requiring prolonged treatment (60–90 days). In addition, significant problems of resistance have emerged with both drugs. In this context, there is an urgent need for more efficacious and safer drugs or drugs regimens, in particular for the treatment of the chronic stage of the infection. Presently, new benznidazole monotherapy regimens with reduced exposure to improve tolerability while maintaining efficacy, and combination regimens of benznidazole with fosravuconazole to improve efficacy are being developed [50].
3. Terpenic Compounds with Antitrypanosomal Activity
Herein, 150 terpenic compounds with antitrypanosomal activity, isolated from plants or obtained by derivatization, and reported in the literature from 2016 to 2021, are presented (Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13, Figure 14 and Figure 15 and Table 1 and Table 2). For clarity reasons, the terpenes are divided into four classes: monoterpenes and iridoids (C10, Figure 3), sesquiterpenes (C15, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9), diterpenes (C20, Figure 10, Figure 11 and Figure 12), and triterpenes (C30, Figure 13, Figure 14 and Figure 15). The selected compounds were tested for their in vitro activity against T. brucei (T. b. brucei, and T. b. rhodesiense), and T. cruzi. Additionally, the in vivo results of some compounds were also described.
The in vitro antitrypanosomal activities are expressed in micromolar concentrations (µM) and some were transformed into this unit to allow an accurate comparison. Furthermore, the in vitro cytotoxicity of these compounds on mammalian cells lines is also indicated, when evaluated simultaneously, allowing the assessment of the selectivity index (SI). SI is defined as the ratio between the half-maximal cytotoxic concentration against the mammalian cell line (CC50) and the half-maximal inhibitory concentration against the parasite (IC50) [51,52]. Although the SI values do not allow extrapolation to the in vivo condition, this parameter is valuable for the selection of compounds with selective activity against trypanosomes.
Presently, there are general and specific criteria proposed by DNDi aiming at identifying hit and lead compounds for further development of drugs against trypanosomiasis, and other infectious diseases. Although these criteria are not strictly applied, they are very important to guide the development of hit and lead series, taking into account their potency, selectivity, toxicity, and chemical profile, among other requirements [51]. For a hit definition, the criteria are divided into two main sets: the disease-specific criteria that focus on potency, efficacy, pathogenicity, and the compound-specific criteria that evaluate the chemical profile of the compounds, in silico pharmacokinetics and pharmacodynamics (DMPK), as well as the physical properties that are predictive of oral therapy [52]. Accordingly, a compound is considered active if it has an IC50 ≤ 10 µM in the in vitro assay against the bloodstream forms of T. b. brucei subspecies, and against the T. cruzi intracellular amastigote forms (TcVI (Tulahuen) or TcII/Y strain) [51,52]. The selectivity of the promising hit compound should be 10-fold higher for the parasite than for the mammalian cell line tested. On the other hand, a lead compound for a future drug against HAT or CD should display an IC50 value more than 10–20-fold higher than the IC50 value of the hit compound, and ideally, its selectivity should be ≥ 50 times higher for the parasite than for the mammalian cell line. Moreover, a significant reduction in parasitemia and/or increase in life-span should be observed in the acute mouse model of HAT at the end of the treatment with up to 4 doses at 50 mg/kg (i.p or p.o). Concerning CD, the lead selection criteria include a hit that causes an 80% parasitemia reduction in organs or tissues, or no parasites detected at the end of treatment and an increase in lifespan with up to 10 doses at 50 mg/kg (p.o) in a mouse model [51,52].
3.1. Monoterpenes and Iridoids
Compound 1 is a limonene benzaldehyde-thiosemicarbazone derivative that showed in vitro antitrypanosomal activity and high selectivity against T. cruzi amastigotes (IC50 1.3 µM, CC50 795 µM, mammalian LLCMK2; SI = 611.2). It is believed that this compound act by inhibiting the proliferation of T. cruzi and inducing morphological changes that lead to the cell death of the parasite. In addition, a reduction in cell volume, depolarization of the mitochondrial membrane and an increase in production of reactive oxygen species (ROS) were also observed. Due to promising in vitro results, an in vivo study was performed on a murine model of acute Chagas disease, and a significant reduction in parasitemia in animals treated with 1 alone (100 mg/kg/day) or combined with benznidazole (5 mg/kg/day each) was found, when compared to the untreated animals. Moreover, it was observed that the survival rate of the animals treated with both compounds during the period of infection was the same that the group treated just with benznidazole, however, with only 5% of the dose used [53].
The essential oil of Origanum onites L. and its major components carvacrol (2) and thymol (3) were evaluated for their antitrypanosomal activity against T. b. rhodesiense trypomastigote forms (mammalian stage), and T. cruzi amastigotes. Good results were only observed against T. b. rhodesiense, and both compounds showed IC50 values of 1.0 µM and 0.73 µM, respectively, and a high selectivity for the parasite (SI = 327.5 and 454.4, respectively, L6 mammalian cell line). Additionally, in the in vivo T. b. brucei mouse model, only compound 3 extended the mean survival of animals, while none cured the infected animals when compared to the reference drug pentamidine [54].
The essential oils of some Apiaceae plants (Echinophora spinosa L., Sison amomum L., Crithmum maritimum L., Helosciadium nodiflorum (L.) W.D.J.Koch) were studied against T. brucei bloodstream forms (TC221 BSFs strain), showing IC50 values in the range of 2.7–10.7 µg/mL. From those, only the essential oil of C. maritimum had a good selectivity (SI = 13, mouse Balb3T3 fibroblasts cell line). Additionally, using the same parasite, the trypanocidal activity of the major compounds (4–6) of C. maritimum was also tested. Terpinolene (4) was the most potent showing an IC50 value of 0.26 µM with a SI = 180. Two other compounds displayed promising activities on the same model namely α-pinene (5, IC50 = 7.4 µM, SI > 100) and β-ocimene (6, IC50 = 8 µM, SI > 91) [55].
Three tetracyclic iridoids (7–9) were isolated from Morinda lucida Benth., a plant traditionally used to treat parasitic diseases in West Africa. Iridoids were evaluated for their in vitro activity against the bloodstream forms of T. b. brucei. Compound 7 was the most active (IC50 0.43 μM) and less toxic than 8 (IC50 1.27 μM), displaying CC50 values of 14.24 μM (SI = 33.1) and 4.74 μM (SI = 3.7), respectively [56]. The activity of compound 9 is lower than 7 and 8 (IC50 3.75 μM), but it did not show cytotoxicity (CC50 ≤ 50 µM) [56]. The main structural differences between compounds 7–9 are the functional groups at C-4 on the side chain. Compound 9 has a carboxylic acid, while compound 7 and 8 have ethyl ester and methyl functional groups, respectively [56]. SI values of the three compounds showed that 7 and 9 are more specific against the parasite than compound 8. Compound 7 was tested in vivo, and a complete clearance of parasitemia, with 100% cure for 20 days post infection, was observed, when 5 consecutive daily shots of 30 mg/kg of compound 7 were taken. It was concluded that compounds 7 and 9 suppressed the expression of paraflagellum rod protein subunit 2, and caused cell cycle alteration, which can preceded apoptosis induction in the bloodstream form of the parasite [56].
3.2. Sesquiterpenes
Two sesquiterpene lactones (10 and 11) were isolated from the methanolic extract of Tithonia diversifolia (Hemsl) A. Grey, and their activities were evaluated against the blood forms of T. brucei [57]. Compound 10 was the most active, exhibiting a very low IC50 value (0.012 μM), but displaying a high cytotoxicity on the mammalian fibroblasts cells (CC50 0.036 μM, SI = 3). Likewise, compound 11 showed antitrypanosomal activity (IC50 0.97 μM) and high cytotoxicity (CC50 1.27 μM, SI = 1.3) [57]. Some considerations can be made regarding the mechanism of action of these sesquiterpene lactones in the parasite cells. Due to the characteristic α,β-unsaturated lactone function present in these structures, which can act as a Michael acceptor, these compounds react with nucleophiles, such as thiol groups in proteins, leading to macromolecular dysfunction, oxidative stress and genetic mutations [57]. The presence of an extra carbonyl group conjugated with two double bonds in 10 can explain the higher antitrypanosomal activity of 10 when compared with 11. In fact, the mechanism of action of these compounds against trypanosomes may be related with formation of thiol adducts with components found in the intracellular medium (namely trypanothione, glutathione and thiol groups in proteins). The parasite’s cells become more vulnerable to oxidative stress with reduction in trypanothione [57].
Several sesquiterpene lactones were isolated from the dichloromethane extract of Vernonia cinerascens Sch.Bip., and evaluated for their in vitro activity against the blood forms of T. b. rhodesiense and for cytotoxicity on the L6 mammalian cell line [58]. Lactone 12 was the most active and selective exhibiting an IC50 value of 0.16 µM and SI = 35. Compounds 13 and 14 also showed activity against T. b rhodesiense (IC50 values of 0.5 µM and 1.1 µM, SI = 13 and 4.2, respectively), being lactone 13 the most selective [58]. Compounds 15 and 16 exhibited similar activities; however, compound 15 showed a higher selectivity (IC50 values of 4.8 µM and 5.0 µM, SI = 27 and 4.3, respectively). Moreover, lactones 12–16 displayed the lowest cytotoxicity in the cells tested [CC50 5.6 µM (12), CC50 6.9 µM (13), CC50 4.7 µM (14), CC50 128 µM (15), CC50 22 µM (16)] [58,59].
Two sesquiterpene lactones (17 and 18) isolated from the dichloromethane extract of Tarchonanthus camphoratus L. aerial parts, and twenty sesquiterpene lactones, including compounds 19–27 obtained from Schkuhria pinnata (Lam.) Kuntze ex Thell. were studied for their in vitro antitrypanosomal activity and cytotoxicity on mammalian L6 cell lines [60]. Lactones 17 and 18 were active against T. b. rhodesiense with IC50 values of 0.39 μM and 2.8 μM, respectively. Furthermore, 17 (SI = 18.6, CC50 7.2 μM) proved to be more selective although it was more cytotoxic than 18 (SI = 6.2, CC50 17.3 μM) [60]. Regarding compounds isolated from S. pinnata, most of them displayed antitrypanosomal activity with IC50 values ranging from 0.10 to 7.30 μM, Compounds 19 and 20 stood out for their high activities against the trypomastigotes, with IC50 values of 0.10 and 0.13 μM, respectively. However, they exhibited cytotoxicity on the cells tested (CC50 values of 2.10 and 3.90 μM, respectively) despite exhibiting some selectivity (SI = 20.5 and 29.7, respectively). Compounds 21 (IC50 0.35 μM, SI = 11.5) and 22 (IC50 0.52 μM, SI = 13) were particularly active, but cytotoxic against the mammalian cell lines assayed (CC50 values of 4.10 and 6.80 μM, respectively). Moreover, compounds 23 (IC50 0.60 μM, SI = 19.2), 24 (IC50 0.82 μM, SI = 13.4), 25 (IC50 0.91 μM, SI = 15.8) and 26 (IC50 0.92 μM, SI = 15.8) also showed very good activities. Finally, sesquiterpene 27 display an IC50 value of 1.7 μM, and was the most selective and least cytotoxic in this group of compounds (SI = 31.1 and CC50 54.6 μM) [60].
Three sesquiterpene lactones (28–30) were isolated from the dichloromethane extract of Achillea fragrantissima (Forssk.) Sch.Bip. and tested against trypomastigote forms of T. b. brucei [61]. Lactone 28 was the most active (IC50 3.03 μM), while lactones 29 and 30 showed the same activity against the parasite with IC50 value of 10.97 μM. The authors did not assess the cytotoxicity of these compounds on mammalian cells [61].
Four sesquiterpene lactones were isolated by bioassay-guided fractionation from extracts of Mikania variifolia Hieron. and Mikania micrantha Kunth, and evaluated against the epimastigote, trypomastigote and amastigote forms of T. cruzi. Compounds 31 and 32 were found in both extracts (2.2% and 0.4% for M. variifolia, and 21.0% and 6.4% for M. micrantha, respectively, calculated based on the dry extract) [62]. Three of the isolated lactones (31, 32, and 33) showed trypanocidal activity, being active against the epimastigote form with IC50 values of 2.41 (SI = 31.9), 0.29 (SI = 992.5) and 8.55 (SI = 5.2) μM, respectively [62]. Compounds 31, 32 and 33 also displayed activity against the trypomastigote form of the parasite with IC50 values of 7.24 (SI = 10.6), 5.43 (SI = 54.0) and 1.03 (SI = 49.0) μM, respectively. Finally, the activities of 31, 32 and 33 against the amastigote forms were lower than those observed for the two previous forms of the parasite (IC50 15.5, 22.8 and 29.1 μM, and SI = 4.3, 12.5, and 1.5, respectively). From those compounds, 32 was the most selective for the human infective parasite, showing a SI of 54 when assayed on human monocyte leukemia THP1 cells. Due to its good selectivity, 32 was also tested in an in vivo model of T. cruzi infection, and was able to decrease the parasitemia and the weight loss associated with the acute phase of the parasite infection. Additionally, 70% of treated mice (1 mg/kg of body weight/day) survived, while all of the control mice died by day 22 after the infection. The authors also observed that this compound increased the production of TNF-α and IL-12 by macrophages [62].
The essential oils from different parts of Smyrnium olusatrum L. were evaluated against the bloodstream forms of T. b. brucei. All oils effectively inhibited the growth of the parasite [63]. From the main constituents of essential oils, sesquiterpene 34 exhibited a significant and selective inhibitory activity against the tested parasite (IC50 3.0 µM, SI = 30, mouse Balb/3T3 fibroblast)) [63].
From Anthemis nobilis L. dichlorometane extract, 19 sesquiterpene lactones, including 15 germacranolides, 2 seco-sesquiterpenes, 1 guaianolide sesquiterpene lactone, and 1 cadinane acid were obtained [64]. Among these compounds, thirteen were tested for their in vitro activity against the bloodstream forms of T. b rhodesiense, with compound 35 being the most potent and selective (IC50 0.08 μM, SI = 63.1). Compounds 36–38 exhibited also a significant anti-trypanosomal activity, but with lower selectivity (36, IC50 0.61 μM, SI = 8.3; 37, IC50 0.36 μM, SI = 14.1; 38, IC50 0.88 μM, SI = 8.3). Moreover, the compounds were assessed against T. cruzi intracellular amastigotes, and the best result was observed for compound 39 (IC50 2.8 μM), but a very low selectivity index was also observed (SI = 0.5). Compound 39 also exhibited a good activity against T. b rhodesiense (IC50 0.4 μM), but with a low selectivity due to its cytotoxicity on mammalian L6 cells (CC50 1.5 μM, SI = 3.8). Compound 40 was considered the one with higher selectivity for T. cruzi (IC50 4.2 μM, SI = 6.1) [64].
Calea pinnatifida (R. Br.) Less. is used in folk medicine as giardicidal, amoebicidal and to treat digestive disorders. Its phytochemical study led to the isolation of a furanoheliangolide sesquiterpene lactone (11,13-dihydroxy-calaxin, 41) which showed a promising trypanocidal activity, displaying an IC50 value of 8.30 μM against T. cruzi amastigotes, and inhibiting the parasite growth in 94.3%. However, compound 41 presented a low selectivity for the parasite cells (CC50 < 15.60 μM on THP-1 cells) [65].
Three sesquiterpene lactones (42–44) were isolated from Smallanthus sonchifolius (Poepp.) H. Rob. and evaluated against T. cruzi epimastigotes, using benzonidazole as positive control [66]. Compounds 42 and 43 showed identical activities with IC50 values of 0.78 and 0.79 μM, respectively. Lactone 44 was also active exhibiting an IC50 value of 1.38 μM. All compounds were more effective than benzonidazole (IC50 10.6 μM). The authors did not assess the cytotoxicity of these compounds on mammalian cells. Due to the high in vitro activity, compounds 42 and 43 were also tested on mice inoculated with T. cruzi trypomastigotes. A significant decrease in circulating parasites (50–71%) was observed, with no signs of toxicity in the dose administrated (1 mg/kg/day). Complementary studies showed marked ultrastructural alteration in trypanosome parasites when treated with these compounds [66].
Several sesquiterpenes isolated from the Cameroonian spice Scleria striatinux De Wild. were studied for their in vitro and in silico antiparasitic activity [67]. From those, sesquiterpene 45 exhibited the best activity against T. cruzi and T. b. rhodesiense bloodstreams forms, with IC50 values of 0.025 μM (SI = 0.74) and 0.002 μM (SI = 8.3), respectively, but the compounds were cytotoxic on HT-29 (human bladder carcinoma) cells. On the other hand, compound 46 showed better activity against T. b. rhodesiense than T. cruzi, with IC50 values of 0.025 μM (SI = 3.4) and 0.085 μM (SI = 1), respectively [67]. The in silico drug metabolism and pharmacokinetic parameters of these two sesquiterpene isomers were also studied, showing compound 46 a good solubility profile, moderate partition coefficient and acceptable in silico pharmacokinetic properties. Similar characteristics were observed for compound 45, but with less optimal parameters, namely for the partition coefficient [67]. Nevertheless, despite the good pharmacokinetic features and the low IC50 values observed, SI values were very small, and compounds also showed a considerable cytotoxicity against HT-29 cell line, which reduces its possible application as a hit compound.
Vernonia lasiopus (O.Hoffm.) H.Rob. extracts were obtained with solvents of different polarities and evaluated in vitro for antiprotozoal activity [59]. The dichloromethane extract was shown to be particularly active against T. b. rhodesiense, and its phytochemical study led to the isolation and identification of six sesquiterpene lactones. These compounds were tested for their in vitro antitrypanosomal activity and cytotoxicity on L6 mammalian cells [59]. Compound 47, the main component of the extract, was the most potent against T. b. rhodesiense trypomastigotes (IC50 0.185 µM); however, it displayed some cytotoxicity (CC50 2.68 µM and SI = 14.5). Moreover, compound 48 presented very similar values (IC50 0.26 µM, CC50 3.67 µM, SI = 14.4). Lactone 49 was the least selective (SI = 4.5) and displayed some cytotoxicity (CC50 2.26 µM), despite showing a considerable antitrypanosomal activity (IC50 0.51 µM). Compound 50 was the least cytotoxic compound in this group (CC50 34.6 µM, SI = 13.7) still showing a good activity against trypomastigotes (IC50 2.53 µM) [59].
On a recent work, the sesquiterpene lactones eupatoriopicrin (51), estafietin (52), eupahakonenin B (53) and minimolide (54) isolated from Argentinean Astearaceae species, which had previously showed activity against T. cruzi epimastigotes, were tested against other forms of the parasite [68]. On the bloodstream forms of T. cruzi the IC50 values obtained were 19.9 µM (51, SI = 12.9), 33.0 µM (53, SI = 10.4), and 21.0 µM (54, SI = 12.8). On the intracellular T. cruzi amastigotes the most active compound was 51 with an IC50 value of 6.3 µM (SI = 40.6). Moderate activities were observed for compound 54 (IC50 = 25.1 µM; SI = 10.7), and 53 (IC50 = 89.3 µM; SI = 3.8) against the same form of the parasite. The majority of compounds showed a significant selectivity for the parasite forms tested compared to Vero cells. The in vivo administration of eupatoriopicrin (51, 1 mg/kg/day) to mice infected with T. cruzi trypomastigotes, for five consecutive days, produced a significant reduction in the parasitemia levels in comparison with non-treated animals (area under parasitemia curves 4.48 vs. 30.47, respectively), being this reduction similar to that achieved with the reference drug, benznidazole. Authors also presented some information regarding the prevention of tissue damage during the chronic phase of the parasite infection, showing beneficial effects on skeletal and cardiac muscular tissues of infected mice treated with the sesquiterpenoid compound. Compound 52 was inactive [68].
The sesquiterpene lactone 55 (tagitinin C) isolated from leaves of Tithonia diversifolia (Hemsl.) A. Gray showed a high inhibition activity against the epimastigote forms of T. cruzi, with IC50 of 1.15 μM, being more active than benznidazole (35.81 μM). However, the cytotoxic concentration (6.54 μM) and the selectivity index (5.69) of compound 55 did not show to be favorable. In an in vivo combination assay, it was observed a complete suppression of parasitemia and parasitological cure in all infected mice (100%) compared to those receiving benznidazole alone (70%). Moreover, despite its lower in vitro selectivity index, compound 55 was well tolerated during the in vivo assays. Interestingly, it was also found that tagitinin C was able to reduce myocarditis, especially when combined with benznidazole [69].
The antitrypanosomal potential of three sesquiterpene lactones (56–58) isolated from Helianthus tuberosus L. (Asteraceae) was evaluated against T. b. rhodesiense trypamastigote bloodstream form (56, IC50 0.077 μM; 57, 0.26 μM; 58, 0.92 μM) and T. cruzi trypomastigotes (56, IC50 1.6 μM; 57, 3.1 μM; 58, 5.7 μM); however, the selectivity index was not promising (CC50 between 0.52 and 3.9 μM, on L6 rat skeletal myoblasts) [70].
Seventeen sesquiterpene lactones were isolated from five plant species of Vernonieae tribe and assessed against T. cruzi epimastigotes [71]. The best trypanocidal effect was observed by elephantopus-type sesquiterpene lactones 59 (IC50 1.5 µM) and 60 (IC50 2.1 µM), obtained from Vernonanthura nebularum (Cabrera) H. Rob., and hirsutinolide 61 (IC50 2.0 µM), isolated from Vernonanthura pinguis (Griseb.) H.Rob. Furthermore, these compounds showed a high selectivity for the parasite (SI > 14) when compared to their cytotoxic effect against the mammalian Hela cells. Compounds 62–65, also isolated from V. nebularum, showed a good antitrypanosomal activity on the same strain with IC50 values ranging from 3.7 to 9.7 µM, being compound 62 the most selective (IC50 = 3.7 µM and SI = 14.3). From V. pinguis, besides compound 61, compounds 66 (IC50 10.7 µM; SI = 9.0) and 67 (IC50 8.1 µM; SI = 13.9) were also isolated and displayed a significant activity; however, it was lower than the observed to hirsutinolide (61). From the remaining species, compound 68 (IC50 6.8 µM; SI = 1.6) isolated from Centratherum puctatum ssp. Punctatum Cass. and compound 69 (IC50 4.7 µM; SI = 11.5) isolated from Elephantopus mollis Kunth also showed antitrypanosomal activity [71].
The sesquiterpene lactones eucannabinolide (70) and santhemoidin C (71), isolated from the dewaxed dichloromethane extract of Urolepis hecatantha (DC.) R.King & H.Rob., were active on T. cruzi epimastigotes with IC50 values of 10 µM and 18 µM, respectively. Both compounds showed low SI values (CC50 > 15 µM for 70 and CC50 = 15 µM for 71) [72].
Goyazensolide (72) is a sesquiterpene lactone isolated from Lychnophora passerina (Mart ex DC) Gardn. that displayed promising results against the intracellular amastigote form of T. cruzi (IC50 = 0.181 µM/24 h, and IC50 = 0.020 µM/48 h), showing a higher selectivity index than the positive control benznidazol (SI = 52.82 and 915.0 for 72, at 24 h and 48 h, respectively, and SI = 4.85 and 41.0 for benznidazol, at 24 h and 48 h, respectively). Further in vivo assays were performed and 72 showed an important therapeutic activity in mice infected with T. cruzi, which was demonstrated by the high percentage of negative parasitological tests employed by the authors in the successive post-treatment evaluations [73].
From the leaves of Hedyosmum brasiliense Mart. Ex Miq., five sesquiterpene lactones together with a sesterpene were isolated and tested against the amastigote and trypomastigote forms of T. cruzi. Among the assessed compounds, compound 73, with a rare terpenoid structure, was the most active displaying an IC50 value of 21.6 μM and SI > 9 for the amastigote form, and an IC50 value of 28.1 μM and SI > 7 for the trypomastigote. The remaining compounds were inactive, excepting compound 74 that exhibited a very weak activity against both parasite forms tested, and a decrease in selectivity for the parasite when compared with selectivity of compound 73 [74].
The bio-guided fractionation of ethanolic extract of leaves of Inula viscosa (L.) Greuter (Asteraceae) led to the isolation of two sesquiterpenoids (75 and 76), which were tested against T. cruzi epimastigotes with IC50 values of 4.99 µM and 15.52 µM, respectively. Both compounds showed modest SI (3.67 and 3.38, respectively), when compared to murine macrophages cells [75]. A preliminary structure-activity study of these compounds demonstrated the importance of the lactone ring to the antiparasitic activity. Regarding the mechanism of action, authors suggested that compounds induced programmed cell death in the tested parasite [75].
Costic acid (77), a eudesmane sesquiterpenoid isolated from the bio-guided fractionation of the n-hexane extract of Nectandra barbellata Coe-Teix. Twigs (Lauraceae) induced a trypanocidal effect with high selectivity for the intracellular amastigote form of T. cruzi (IC50 7.9 µM). A modest activity against T. cruzi trypomastigotes was also observed (IC50 37.8 µM). No cytotoxicity was observed on L929 human cells, revealing its selectivity for both forms of the parasite (CC50 > 200 µM, SI > 25 on amastigote forms and SI > 5 on trypomastigote forms). The authors suggested that costic acid (77) has a key action on the mitochondria activity of the parasite [76].
Some germacranolide sesquiterpene lactones were isolated from the aerial parts and flowers of Tanacetum sonbolii Mozaff. Compounds 78 and 79 were the most active showing an IC50 of 5.1 and 10.2 µM, respectively, against T. b. rhodesiense bloodstream forms, and SI values of 3.9 (78) and 4.0 (79) when compared with rat myoblast (L6) cells [77].
The bicyclic drimane-type sesquiterpene polygodial (80), firstly isolated from Polygonum hydropiper L. (Polygonaceae), and some natural and synthetic compounds of the same family were evaluated for growth inhibition against the amastigote, trypomastigote, and epimastigote forms of T. cruzi. The parent drug 80 exhibited a moderate inhibitory activity (GI50 = 34.4 μM amastigotes; GI50 = 68.2 μM trypomastigotes; GI50 = 51.0 μM epimastigotes). The best inhibition growth activities were observed for its synthetic derivatives, namely compound 81 (GI50 = 9.9 μM amastigotes; GI50 = 8.4 μM trypomastigotes; GI50 = 13.0 μM epimastigotes), 82 (GI50 = 6.7 μM amastigotes; GI50 = 6.4 μM trypomastigotes; GI50 = 12.3 μM epimastigotes), and 83 (GI50 = 8.3 μM amastigotes; GI50 = 6.9 μM trypomastigotes; GI50 = 7.2 μM epimastigotes). Selectivity index values were not determined. The synthetic α,β-unsaturated phosphonate (83) was favorably compared with the clinically approved drugs benznidazole and nifurtimox during a competition assay, being even effective against trypomastigotes, contrarily to benznidazole that showed no activity against this trypanosomal form. The effect of polygodial derivative 81 on the growth of the parasite in infected human retinal pigment epithelial (ARPE) cells was studied using confocal microscopy. A significant reduction in the intracellular parasites was observed, with no alterations of replication or viability of the cells [78]. Compound 80 was also previously isolated from the Chilean species Drimys winteri, and was tested on the same parasite forms with weak comparable results. On this work, the authors associated the trypanosomal activity of this compound with intracellular effects occurring in the parasite, namely, mitochondrial dysfunctions, ROS production and autophagic phenotype [79].
Epi-polygodial (84), isolated from the Brazilian plant Drimys brasiliensis Miers (Winteraceae), exhibited a high parasite selectivity towards T. cruzi trypomastigotes (IC50 = 5.01 μM, SI > 40 to NCTC cells). Authors correlated the antitrypanosomal activity of this compound with its effects on cellular membranes by the interaction of 84 with DPPE-monolayers (the Langmuir monolayers of dipalmitoylphosphoethanolamine) at the air–water interface, which affects the physical chemical properties of the mixed film [80].
The sesquiterpene (-)-T-cadinol (85) isolated from Casearia sylvestris Sw. displayed a moderate activity against amastigotes and trypomastigotes forms of T. cruzi with IC50 values of 15.8 and 18.2 μM, respectively, and no toxic effect on the mammalian cells was observed (CC50 200 μM, SI > 15). The mechanism of action was studied using different techniques, and it was observed that 85 affected the parasite mitochondria. However, additional studies are necessary in order to confirm this organelle as a candidate target [81].
A sesquiterpene glycoside ester (86) isolated from the flowers of Calendula officinalis L. have shown a moderate activity against T. brucei (IC50 16.9 μM). A closely similar compound (87), only differing in the type of sugar residue, did not display antitrypanosomal activity, suggesting the importance of sugar moiety conformation to the activity [82].
3.3. Diterpenes
Andrographolide (88) is a labdane-type diterpene, isolated from Andrographis paniculate (Burm. F.) Wall. Ex Nees, with reported anticancer, anti-inflammatory, antioxidant, cardioprotective and hepatoprotective properties [83]. To determine its effect on the viability of T. brucei procyclic trypomastigotes (the form of the parasite that differentiate in the insect gut), the parasites were incubated at different concentrations (0–200 μM) for 72 h. Compound 88 inhibited the growth of the parasite, exhibiting an IC50 = 8.3 μM and SI = 8.5. At this concentration, no cytotoxic effect was observed (CC50 70.5 μM) [83]. Giemsa staining of parasites treated with 88 allowed the observation of morphological changes, in particular, loss of integrity, damage to the cell membrane, general rounding, and loss of cells’ flagella. Ultimately, the authors concluded that the trypanocidal activity of 88 is mediated by inducing the oxidative stress together with the depolarization of the mitochondrial membrane potential, generating an apoptosis-like programmed cell death [83].
The phytochemical study of aerial parts of Baccharis retusa DC., a medicinal plant used in Brazilian folk medicine to treat parasitic diseases, allowed the isolation and identification of the kaurane-type diterpene 89. This compound was active against T. cruzi trypomastigotes (IC50 3.8 μM) with a high selectivity (SI = 50.0) due to its reduced cytotoxicity (CC50 189.7 μM) on NCTC cells [84].
Ent-kaurenoic acid (90) and ent-pimaradienoic acid (91) were used as starting material to obtain several derivatives. From those, the ent-kaurane derivatives 92 (IC50 < 12.5 μM) and 93 (IC50 26.1 μM) showed the highest antitrypanosomal activity when compared to compound 90 (IC50 225.8 μM). Regarding the ent-pimaradienoic acid (91, IC50 68.7 μM) set, compound 94 (IC50 3.8 μM) was the most active against trypomastigotes forms of T. cruzi. However, due to the lack of cytotoxicity data it is not possible to determine the selectivity index of these compounds [85].
Three quinone methide-type diterpenes (95–97) were isolated from the roots of Salvia austriaca Jacq. and tested for their in vitro activity against T. b. rhodesiense and T. cruzi. Cytotoxicity was determined on L6 cells [86]. The diterpene 95 was the most active and selective against T. b. rhodesiense trypomastigotes (IC50 0.05 µM and SI = 38). However, despite exhibiting an IC50 value of 7.11 µM against T. cruzi amastigotes, its selectivity was very low (SI = 0.27). Compounds 96 and 97 also showed activity against both parasites, being more active against T. b. rhodesiense with IC50 values of 0.62 µM (SI = 5.0) and 1.67 µM (SI = 2.4), respectively. Regarding their activity against T. cruzi, an IC50 value of 7.76 µM (SI = 0.4), and 7.63 µM (SI = 0.5) was observed for compound 96 and 97, respectively [86], but the compounds were not selective to the parasite [86].
The phytochemical study of dichloromethane extract of Aldama discolors (Baker) E.E.Schill. & Panero leaves led to the isolation of four structurally and biosynthetically related diterpenes. These were evaluated for in vitro activity against T. b. rhodesiense trypomastigotes and T. cruzi amastigotes [87]. Among the isolated diterpenes, compound 98 showed a moderate in vitro activity with an IC50 value of 15.4 µM against T. cruzi, and an IC50 24.3 µM for T. b. rhodesiense. On the other hand, compound 99, structurally similar to 98, showed less activity against the amastigote forms of T. cruzi (IC50 19.4 µM), and no activity against the trypomastigote forms of T. b. rhodesiense. The selectivity of these compounds was very low, with SI values ranging from 2 to 4 [87].
The commercially available dehydroabietylamine (100), an abietane-type diterpenoid isolated in large amount from Plectranthus genus, was used as a starting material to produce a set of amides derivatives. Among these compounds, 100–103 were tested against T. cruzi amastigotes, showing compound 103 the highest antitrypanosomal activity and selectivity (IC50 0.6 μM; SI = 58). The remaining compounds, including 100, in spite of displaying antitrypanosomal activity (IC50 values between 3.7 and 7.4 μM) were not very selective to the parasite, showing CC50 values ranging from 6.5 to 33.5 μM when tested on the human cell line (SI values between 1 and 6, L6 rat myoblasts) [88].
Leriifolione (104), isolated from the lipophilic extract of the roots of Salvia leriifolia Benth., showed high activity against T. b. rhodesiense (IC50 1.0 μM) and T. cruzi (IC50 4.6 μM), but an undesirable cytotoxicity on L6 cells (SI = 2.6 and 0.6, respectively) [89].
From the n-hexane extract of the roots of Zhumeria majdae Rech. F., eight abietane-type diterpenes were isolated. The antitrypanosomal activity was investigated for compounds 105, 106, and 107 against T. b. rhodesiense and T. cruzi. All the compounds showed a high activity against T. b. rhodesiense, (IC50 = 3.6 µM, 1.8 µM, and 0.1 µM to 105, 106, and 107, respectively), being compound 106 the most selective for the parasite when compared with L6 cell line tested (105, SI = 1.7; 106, SI = 21.9; 107, SI = 15.4). None of the compounds were active against T. cruzi [90].
Seventeen diterpenes were isolated from the aerial parts of Perovskia abrotanoides Kar. These compounds were evaluated for antiprotozoal activity (T. cruzi amastigotes, and T. b rhodesiense trypomastigotes), and cytotoxicity was also assessed on rat skeletal myoblast L6 cell line. Most of the diterpenes were less active against T. cruzi than against T. b rhodesiense [91]. Compound 108 showed the best activity against T. b. rhodesiense (IC50 0.5 µM, SI = 10.5). However, it was inactive against T. cruzi, showing a lack of selectivity (IC50 58.7 µM; SI = 0.1, CC50 5.2 µM). With similar activity, but less cytotoxic (CC50 12.1 µM) than 108, compound 109 displayed an IC50 value of 0.8 µM (SI = 14.9) against T. b. rhodesiense and an IC50 of 34.7 µM (SI = 0.3) against T. cruzi. Compounds 110–113 were particularly active against T. b. rhodesiense, displaying IC50 values ranging from 3.8 to 12 µM but low SI values (SI = 1.5–12.5) [91].
Bokkosin (114), a new cassane diterpene isolated from the Nigerian species Calliandra portoricensis Hassk. used in traditional medicine to treat tuberculosis, and helmintic diseases, showed a strong trypanocidal activity against the bloodstream forms of T. b. brucei, sensitive (IC50 = 1.1. μM) and resistant to pentamidine (IC50 = 0.5 μM). A highly favorable selectivity for the parasite strains was also observed, when compared with its cytotoxic effect on two mammalian cell lines (CC50 = 269 μM; SI = 246, on U937; CC50 = 230 μM; SI = 215, on RAW 246.7) [92].
From the n-hexane and ethylacetate extracts of the roots of Acacia nilotica L. several diterpenes were isolated. Among them, the cassane-type diterpenoid 115 was tested against the T. b brucei bloodstream form, exhibiting a high activity a (IC50 1.4 μM). Additionally, 115 was tested for its cytotoxic effect on human HEK cells (CC50 = 29.5 μM; SI = 21.1) displaying a significant selectivity for the parasite [93].
3.4. Triterpenes
Ursolic acid (116) was tested for antitrypanosomal activity against T. brucei trypomastigotes displaying an IC50 value of 3.35 μM. Similar IC50 values were also obtained by Catteau et al. (IC50 2.4 μM, CC50 > 11.1 μM). However, it is important to note that the compound showed a lack of selectivity for the trypanosoma parasite due to its cytotoxic effect against the mammalian WI38 cells used [94]. In order two find out a possible mode of action, in silico molecular modelling studies were also performed using the parasitic enzymes of the trypanosome, namely trypanothione reductase, methionyl-tRNA synthetase, and inosine-adenosine-guanosine nucleoside hydrolase [95]. Ursolic acid showed a good binding affinity for trypanothione reductase and methionyl-tRNA synthetase, which was higher than that obtain for the reference drug difluoromethylornithine. On the other hand, no inhibition was observed for inosine–adenosine–guanosine. These data may suggest that the inhibition of the two former enzymes may be responsible for the antitrypanosomal activity of compound 116 [95].
A new ursane-type triterpenoid glycoside (117) isolated from the dried roots of Vangueria agrestis (Schweinf. ex Hiern) Lantz exhibited a considerable growth suppressing effect against T. brucei trypomastigotes (IC50 11.1 μM and IC90 12.3 μM). However, no cytotoxicity studies on mammalian cell lines were performed [96].
Betulin acid (118), a lupane-type pentacyclic triterpene, and some semysynthetic amide derivatives were tested against T. cruzi trypomastigotes. Compound 118 showed a moderate activity (IC50 19.5 µM and SI = 18.8), while an increasing activity was observed in derivatives 119 (IC50 1.8 µM, SI = 17.3), 120 (IC50 5.0 µM, SI = 10.7), and 121 (IC50 5.4 µM, SI = 5.3). The mechanism of action of compound 119 in trypomastigotes was studied and seemed to be associated with the death of the parasite by necrosis, characterized by the rupture of the membrane, flagellar retraction, and appearance of atypical cytoplasmic vacuoles and dilation of the Golgi cisterns. Furthermore, the amide derivatives of compound 118 act by reduction in the invasion process, as well as the development of the intracellular parasite in host cells [97]. Sousa et al. corroborated those mechanistic results, showing that betulinic acid was able to inhibit all the development forms of T. cruzi (namely epimastigotes, trypomastigotes and amastigotes) not only by using necrotic processes but also due to modifications on the parasite mitochondrial membrane potential and the increase in reactive oxygen species [98].
Six limonoids (122–127) obtained from the roots of Pseudocedrela kotschyi (Schweinf.) Harms were investigated for their trypanocidal activity using bloodstream forms of T. brucei, showing GI50 values ranging from 2.5 to 14.5 μM. The most active compound was 122 (GI50 2.5 μM) but showed some cytotoxicity on human HL-60 cells (GI50 31.5 μM). On the other hand, limonoid 125 exhibited a similar activity (GI50 3.18 μM) with no cytotoxic effect on the mammalian cell line (GI50 > 100 μM) [99].
From Tabernaemontana longipes Donn.Sm., baurenol acetate (128) was isolated and tested for its ability to inhibit the growth of T. brucei bloodstream forms, showing an IC50 value of 3.1 μM. Baurenol (129) displayed a higher activity against the same parasite (IC50 = 2.7 μM). Both compounds showed a low effect on cellular viability on Hep G2 cells (IC50 > 80 μM) [100].
Four new triterpenoids (130–133), with a rare scaffold, isolated from Salvia hydrangea DC. ex Benth. showed antitrypanosomal activity against T. cruzi amastigotes with IC50 values ranging from 3.5 to 19.8 μM, with no significant activity against T. b. rhodesiense trypomastigotes All the compounds showed a modest selectivity (SI ranging from 2.4 to 10.7; L6 cell line) [101].
Two lanostane-type triterpenoids, polycarpol (134) and dihydropolycarpol (135) were isolated from Greenwayodendron suaveolens (Engl. & Diels) Verdc., a plant traditional used in Congo to treat malaria. Both compounds were tested against T. b. brucei (IC50 = 8.1 μM; SI < 1.0, 134; IC50 = 8.1 μM; SI = 2.4, 135), and T. cruzi (IC50 = 1.4 μM; SI = 2.0, 134; IC50 = 2.4 μM; SI = 8.1, 135), showing good activities but very low selectivity [102].
From Buxus sempervirens L. leaf extract, several triterpenic-alkaloid derivatives were isolated, being the majority tested for their activity against T. b. rhodesiense. Cytotoxicity assays were performed on mammalian L6 cells. Compounds 136–142 displayed high activities with IC50 values < 3 μM. The highest activities and selectivities for the parasite strain tested were obtained for compounds 136 (IC50 = 1.5 μM; SI = 25), 138 (IC50 = 2.3 μM; SI = 42), 141 (IC50 = 2.4 μM; SI = 30), and 142 (IC50 = 1.3 μM; SI = 33). In spite of its promising activity against the parasite (IC50 = 1.1 μM), compound 146 showed a lower selectivity (SI = 12). The remaining compounds 143–150 displayed a significant activity with IC50 values ranging from 3.1 μM (compound 149) to 9.0 μM (compound 144), but a modest selectivity to the parasite (SI < 9) [103].
4. Discussion
Human African trypanosomiasis and American trypanosomiasis continue to be a major public health problem, affecting a significant proportion of the world’s population, especially in tropical countries. Currently, the drugs used to treat these diseases are scarce and far from being ideal. Therefore, the discovery and development of new drugs and treatments should be a continuous process, and all possible approaches should be explored, mainly focusing on multidisciplinary collaborations. It is also important to stress that the new drugs should be affordable and easy to administrate, improving the adherence to the therapeutic protocol, and decreasing the need for patient hospitalization.
Several approaches have been considered for the development of new drugs against these diseases. Due to the high costs and slow pace of new drug discovery, one of the main strategies is the repositioning or repurposing of drugs that were developed and used to treat other diseases. Although several advantages can be addressed, including the lower risk of failing, reduced time frame for drug development, less investment and rapid return [104], drug repositioning have also major drawbacks. These include, for example, the existence of undesirable side effects, problems concerning a different target population, poor stability in conditions of high temperature and humidity, lack of oral bioavailability, and various regulatory issues and intellectual property barriers. Therefore, the development of a new drug that ideally would be suitable for combination therapy, increasing the clinical efficacy, and decreasing side effects and the development of resistance, is a goal of utmost importance [10]. Besides combinatorial chemistry, one of the main strategies for drug discovery is through the phytochemical study of plants or other sources of natural origin. The importance of natural products for the development of drug leads or actual drugs along the last three decades has been reported in various reviews [12].
The evaluation of compounds with anti-parasitic activity is usually performed by two main approaches: the target-based and the phenotypic approaches [10,104]. The target-based methodologies (Section 2.1.1 and Section 2.2.1) focus on the specific biochemical pathway of the parasites, and consist of the identification of possible molecular targets (e.g., enzymes) that are significantly involved in the disease and the screening of molecules that possibly interfere with these targets. However, regarding trypanosomiasis, restricted success has been achieved, possibly due to a lack of translation between the activity in the molecular target, and the result on the proliferation of the parasite. In fact, the number of robust and validated molecular targets against these diseases is very limited, and in addition, for several drugs currently used in clinic, the mode of action is not yet completely understood or it comprises several targets [10,104]. By far, the most widely used methodologies to identify anti-trypanosomal drugs are based on the phenotypic methods. They consist of the screening of the compounds directly against the different forms of the parasite, and most of the time, for the bioactive compounds, there is no knowledge regarding their underlying mode of action or their molecular target. Through this screening, the effects on the parasite and the host cell viability (toxicity) can be assessed simultaneously [10].
In this review, 150 terpenic compounds obtained by isolation or derivatization from different plant species, were grouped into four classes of terpenes, namely, monoterpenes and iridoids, sesquiterpenes, diterpenes and triterpenes. The scope of this review was limited to compounds that exhibited in vitro or in vivo activity against the diverse forms of T. brucei and/or T. cruzi, displaying IC50 values in low micromolar range, most of them below 10 μM.
Regarding the anti-T. brucei activity, it can be observed that most of the selected compounds were active on the trypomastigote bloodstream form of the parasite (in vitro assays), and several possible hits can be identified. A high number of compounds exhibited very low IC50 values (0.05 < IC50 < 3.0 μM), when tested against T. b. rhodesiense or T. b. brucei, also presenting low cytotoxicity on the mammalian cells (SI values higher than 10). Some of the most promising hits are depicted in Figure 16. It is interesting to note that the majority of the bioactive compounds are sesquiterpenes and more specifically, sesquiterpenic lactones. Indeed, the α,β-unsaturated lactone function is very common in biologically active molecules, being responsible not only for their activity but also for the cytotoxicity of the compounds. The presence of this chemical function promotes the Michael-type addition to a suitable nucleophile (for example, the thiol group in proteins), and may irreversibly alkylate critical enzymes and transcription factors that control gene regulation, protein synthesis, cell metabolism, and ultimately, the cell division [105,106]. After the preliminary in vitro studies, two compounds (3 and 7) were further evaluated using in vivo studies on T. brucei mouse models achieving very good results.
Concerning T. cruzi assays, it is curious to notice that there are much fewer research papers reporting the bioactivity of compounds against this parasite, probably because the in vitro assays using intracellular amastigotes were not so straightforward. Nevertheless, it was possible to select some promising hits that were very active against T. cruzi amastigotes, according to the criteria established by DNDi (Figure 17). Some of the most active compounds (1, 51, and 74) were further evaluated in vivo using a T. cruzi mouse model. The best result was obtained for compound 51, and a significant reduction in parasitemia levels was observed in treated mice (1 mg/Kg/day, for 5 consecutive days), similarly to that obtained with the control group treated with the reference drug benznidazole.
Although most of the reported compounds were remarkably active against some infectious forms of the parasites, some of them also displayed some cytotoxicity on the mammalian cells tested. Furthermore, there is a notorious lack of additional studies on structure-activity relationships (SAR) and on the possible mechanisms of action. In addition, some reasons could be addressed to justify the absence of results for in vivo assays, including that the majority of compounds are isolated in very low amounts, a fact that precludes this type of assay where a large amount of compound is always needed.
5. Conclusions
The data presented in this review gathered the recent scientific research and experimental evidence on the most promising terpenoids derived from plants, and active against T. brucei and T. cruzi parasites. These data represent the immense efforts of various research groups all over the world, and ultimately the collected information is highly pertinent and can be used, for example, to support the selection of other plants to study, using the chemotaxonomic approach. However, all of these studies are strictly academic and no further translation to drug development has been achieved. There is still a limited collaboration of academic institutions with the pharmaceutical industry, and importantly, obtaining opportunities for research funding is, nowadays, even more challenging. All these aspects have limited or made it difficult for academic researchers to advance promising hits for further development.
In the future, natural products research must be a multidisciplinary process combining phytochemistry with innovative technological resources, in a way that will be significantly different from the past. These technologies must include high-throughput screening and in silico methodologies, as well as new extraction and dereplication procedures, new analytical tools, metabolic engineering, omics-based analysis, informatics, and big data analysis, in order to overcome the constraints of the classic natural products research.
Author Contributions
Conceptualization, N.D.; writing—original draft preparation, R.D. and C.R.; writing—review and editing, N.D., C.R., E.M. and A.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Ending the Neglect to Attain the Sustainable Development Goals. A Road Map for Neglected Tropical Diseases 2021–2030; Ntuli, M.M., Ed.; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Hotez, P.J.; Aksoy, S.; Brindley, P.J.; Kamhawi, S. What constitutes a neglected tropical disease? PLoS Negl. Trop. Dis. 2020, 14, e0008001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisuzzaman; Tsuji, N. Schistosomiasis and hookworm infection in humans: Disease burden, pathobiology and anthelmintic vaccines. Parasitol. Int. 2020, 75, 102051. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Corbeil, A.; Do Monte-Neto, R.L.; Fernandez-Prada, C. Of drugs and trypanosomatids: New tools and knowledge to reduce bottlenecks in drug discovery. Genes 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, D.H.; Asamoa-Bah, A.; Fenwick, A.; Savioli, L.; Hotez, P. The history of the neglected tropical disease movement. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 169–175. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Neglected tropical diseases: Impact of COVID-19 and WHO’s response—Maladies tropicales négligées: Impact de la COVID-19 et réponse de l’OMS. Wkly. Epidemiol. Rec. 2020, 95, 461–468. [Google Scholar]
- Cheuka, P.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2017, 22, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capela, R.; Moreira, R.; Lopes, F. An overview of drug resistance in protozoal diseases. Int. J. Mol. Sci. 2019, 20, 5748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Herrera, A.; Cortez-Maya, S.; Bocanegra-Garcia, V.; Banik, B.K.; Rivera, G. Recent Advances in the Development of Broad-Spectrum Antiprotozoal Agents. Curr. Med. Chem. 2020, 28, 583–606. [Google Scholar] [CrossRef] [PubMed]
- Field, M.C.; Horn, D.; Fairlamb, A.H.; Ferguson, M.A.J.; Gray, D.W.; Read, K.D.; De Rycker, M.; Torrie, L.S.; Wyatt, P.G.; Wyllie, S.; et al. Anti-trypanosomatid drug discovery: An ongoing challenge and a continuing need. Nat. Rev. Microbiol. 2017, 15, 217–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournet, A.; Munoz, V.; Muñoz, V. Natural products as trypanocidal, antileishmanial and antimalarial drugs. Curr. Top. Med. Chem. 2002, 2, 1215–1237. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; Da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads Against Protozoan Neglected Diseases—Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Tullius Scotti, M.; Scotti, L.; Ishiki, H.; Fávaro Ribeiro, F.; da Cruz, R.M.D.; de Oliveira, M.P.; Jaime Bezerra Mendonça, F. Natural Products as a Source for Antileishmanial and Antitrypanosomal Agents. Comb. Chem. High Throughput Screen. 2016, 19, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Isah, M.B.; Ibrahim, M.A.; Mohammed, A.; Aliyu, A.B.; Masola, B.; Coetzer, T.H.T. A systematic review of pentacyclic triterpenes and their derivatives as chemotherapeutic agents against tropical parasitic diseases. Parasitology 2016, 143, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Muschietti, L.V.; Ulloa, J.L. Natural Sesquiterpene Lactones as Potential Trypanocidal Therapeutic Agents: A Review. Nat. Prod. Commun. 2016, 11, 1569–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockram, P.E.; Smith, T.K. Active Natural Product Scaffolds against Trypanosomatid Parasites: A Review. J. Nat. Prod. 2018, 81, 2138–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungogo, M.A.; Ebiloma, G.U.; Ichoron, N.; Igoli, J.O.; de Koning, H.P.; Balogun, E.O. A Review of the Antimalarial, Antitrypanosomal, and Antileishmanial Activities of Natural Compounds Isolated From Nigerian Flora. Front. Chem. 2020, 8, 1221. [Google Scholar] [CrossRef]
- Chan-Bacab, M.J.; Reyes-Estebanez, M.M.; Camacho-Chab, J.C.; Ortega-Morales, B.O. Microorganisms as a potential source of molecules to control trypanosomatid diseases. Molecules 2021, 26, 1388. [Google Scholar] [CrossRef]
- Hzounda Fokou, J.B.; Dize, D.; Etame Loe, G.M.; Nko’o, M.H.J.; Ngene, J.P.; Ngoule, C.C.; Boyom, F.F. Anti-leishmanial and anti-trypanosomal natural products from endophytes. Parasitol. Res. 2021, 120, 785–796. [Google Scholar] [CrossRef]
- Lenzi, J.; Costa, T.M.; Alberton, M.D.; Goulart, J.A.G.; Tavares, L.B.B. Medicinal fungi: A source of antiparasitic secondary metabolites. Appl. Microbiol. Biotechnol. 2018, 102, 5791–5810. [Google Scholar] [CrossRef]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.M.; Yang, L.Y.; Huang, S.S.; Chigor, V.N.; Eze, E.A.; Pan, L.X.; Zhang, T.; Yang, D.F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty 2021, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Lucas, L.A.; Cândido, A.C.B.B.; Santos, M.F.C.; Caffrey, C.R.; Bastos, J.K.; Ambrósio, S.R.; Magalhães, L.G. Antiparasitic Properties of Propolis Extracts and Their Compounds. Chem. Biodivers. 2021, 18, 310. [Google Scholar] [CrossRef]
- Asfaram, S.; Fakhar, M.; Keighobadi, M.; Akhtari, J. Promising Anti-Protozoan Activities of Propolis (Bee Glue) as Natural Product: A Review. Acta Parasitol. 2021, 66, 1–12. [Google Scholar] [CrossRef]
- Simoben, C.V.; Ntie-Kang, F.; Akone, S.H.; Sippl, W. Compounds from African Medicinal Plants with Activities Against Selected Parasitic Diseases: Schistosomiasis, Trypanosomiasis and Leishmaniasis. Nat. Products Bioprospect. 2018, 8, 151–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naß, J.; Efferth, T. The activity of Artemisia spp. and their constituents against Trypanosomiasis. Phytomedicine 2018, 47, 184–191. [Google Scholar] [CrossRef] [PubMed]
- de Morais, M.C.; de Souza, J.V.; da Silva Maia Bezerra Filho, C.; Dolabella, S.S.; de Sousa, D.P. Trypanocidal essential oils: A review. Molecules 2020, 25, 4568. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; van Staden, J. Antiprotozoal alkaloid principles of the plant family Amaryllidaceae. Bioorg. Med. Chem. Lett. 2019, 29, 126642. [Google Scholar] [CrossRef] [PubMed]
- Montesino, N.L.; Schmidt, T.J. Salvia species as sources of natural products with antiprotozoal activity. Int. J. Mol. Sci. 2018, 19, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.K.; da Trindade, R.; Alves, N.S.; Figueiredo, P.L.; Maia, J.G.S.; Setzer, W.N. Essential Oils from Neotropical Piper Species and Their Biological Activities. Int. J. Mol. Sci. 2017, 18, 2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Human African Trypanosomiasis (Sleeping Sickness). Available online: https://www.who.int/health-topics/human-african-trypanosomiasis#tab=tab_1 (accessed on 13 January 2022).
- World Health Organization. Trypanosomiasis, Human African (Sleeping Sickness). Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 4 March 2021).
- Drugs for Neglected Diseases Initiative (DNDi) Sleping Sickness. Available online: https://dndi.org/diseases/sleeping-sickness/facts/ (accessed on 7 January 2022).
- Centers for Disease Control and Prevention (CDC) Parasites—African Trypanosomiasis (also Known as Sleeping Sickness). Available online: https://www.cdc.gov/parasites/sleepingsickness/ (accessed on 6 January 2022).
- Keiser, J.; Stich, A.; Burri, C. New drugs for the treatment of human African trypanosomiasis: Research and development. Parasitol. Today 2001, 17, 42–49. [Google Scholar] [CrossRef]
- Brand, S.; Norcross, N.R.; Thompson, S.; Harrison, J.R.; Smith, V.C.; Robinson, D.A.; Torrie, L.S.; McElroy, S.P.; Hallyburton, I.; Norval, S.; et al. Lead optimization of a pyrazole sulfonamide series of trypanosoma brucei N -myristoyltransferase inhibitors: Identification and evaluation of CNS penetrant compounds as potential treatments for stage 2 human african trypanosomiasis. J. Med. Chem. 2014, 57, 9855–9869. [Google Scholar] [CrossRef] [PubMed]
- Bijlmakers, M.J. Ubiquitination and the Proteasome as Drug Targets in Trypanosomatid Diseases. Front. Chem. 2021, 8, e630888. [Google Scholar] [CrossRef] [PubMed]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An Overview on Target-Based Drug Design against Kinetoplastid Protozoan Infections: Human African. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Khan, S.A.; Alshammari, M.K.; Alqahtani, A.M.; Alanazi, T.A.; Kamal, M.; Jawaid, T.; Ghoneim, M.M.; Alshehri, S.; Shakeel, F. Discovery, Development, Inventions and Patent Review of Fexinidazole: The First All-Oral Therapy for Human African Trypanosomiasis. Pharmaceuticals 2022, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Steketee, P.C.; Giordani, F.; Vincent, I.M.; Crouch, K.; Achcar, F.; Dickens, N.J.; Morrison, L.J.; MacLeod, A.; Barrett, M.P. Transcriptional differentiation of Trypanosoma brucei during in vitro acquisition of resistance to acoziborole. PLoS Negl. Trop. Dis. 2021, 15, e0009939. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chagas Disease (also Known as American Trypanosomiasis). Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 4 March 2021).
- Dumonteil, E.; Herrera, C. The Case for the Development of a Chagas Disease Vaccine: Why? How? When? Trop. Med. Infect. Dis. 2021, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Lidani, K.C.F.; Andrade, F.A.; Bavia, L.; Damasceno, F.S.; Beltrame, M.H.; Messias-Reason, I.J.; Sandri, T.L. Chagas disease: From discovery to a worldwide health problem. J. Phys. Oceanogr. 2019, 49, 166. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.M. Chagas Disease: Epidemiology and Barriers to Treatment. Am. J. Med. 2020, 133, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Silva, E.; Morais, L.H.; Clarke, G.; Savino, W.; Peixoto, C. Targeting the Gut Microbiota in Chagas Disease: What Do We Know so Far? Front. Microbiol. 2020, 11, 3083. [Google Scholar] [CrossRef]
- Onyekwelu, K. Life Cycle of Trypanosoma cruzi in the Invertebrate and the Vertebrate Hosts. In Biology of Trypanosoma Cruzi; De Souza, W., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-83968-204-9. [Google Scholar]
- Francisco, A.F.; Jayawardhana, S.; Olmo, F.; Lewis, M.D.; Wilkinson, S.R.; Taylor, M.C.; Kelly, J.M. Challenges in Chagas Disease Drug Development. Molecules 2020, 25, 2799. [Google Scholar] [CrossRef]
- Villalta, F.; Rachakonda, G. Advances in preclinical approaches to Chagas disease drug discovery. Expert Opin. Drug Discov. 2019, 14, 1161–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Huertas, P.; Cardona-Castro, N. Advances in the treatment of Chagas disease: Promising new drugs, plants and targets. Biomed. Pharmacother. 2021, 142, 112020. [Google Scholar] [CrossRef] [PubMed]
- Torrico, F.; Gascón, J.; Barreira, F.; Blum, B.; Almeida, I.C.; Alonso-Vega, C.; Barboza, T.; Bilbe, G.; Correia, E.; Garcia, W.; et al. New regimens of benznidazole monotherapy and in combination with fosravuconazole for treatment of Chagas disease (BENDITA): A phase 2, double-blind, randomised trial. Lancet Infect. Dis. 2021, 21, 1129–1140. [Google Scholar] [CrossRef]
- Don, R.; Ioset, J.-R. Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infections. Parasitology 2014, 141, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Van Huijsduijnen, R.H.; Kaneko, T.; Kita, K.; Mowbray, C.E.; Schmatz, D.; Warner, P.; Slingsby, B.T. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat. Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.C.; Lazarin-Bidóia, D.; Desoti, V.C.; Falzirolli, H.; da Silva, C.C.; Ueda-Nakamura, T.; de O. Silva, S.; Nakamura, C.V. 1,3,4-Thiadiazole derivatives of R-(+)-limonene benzaldehyde-thiosemicarbazones cause death in Trypanosoma cruzi through oxidative stress. Microbes Infect. 2016, 18, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Kaiser, M.; Demirci, B.; Demirci, F.; Hüsnü Can Baser, K. Antiprotozoal Activity of Turkish Origanum onites Essential Oil and Its Components. Molecules 2019, 24, 4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngahang Kamte, S.L.; Ranjbarian, F.; Cianfaglione, K.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Afshar, F.H.; Iannarelli, R.; Benelli, G.; Cappellacci, L.; et al. Identification of highly effective antitrypanosomal compounds in essential oils from the Apiaceae family. Ecotoxicol. Environ. Saf. 2018, 156, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Kwofie, K.D.; Tung, N.H.; Suzuki-Ohashi, M.; Amoa-Bosompem, M.; Adegle, R.; Sakyiamah, M.M.; Ayertey, F.; Owusu, K.B.A.; Tuffour, I.; Atchoglo, P.; et al. Antitrypanosomal activities and mechanisms of action of novel tetracyclic iridoids from Morinda lucida Benth. Antimicrob. Agents Chemother. 2016, 60, 3283–3290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sut, S.; Dall’Acqua, S.; Baldan, V.; Ngahang Kamte, S.L.; Ranjbarian, F.; Biapa Nya, P.C.; Vittori, S.; Benelli, G.; Maggi, F.; Cappellacci, L.; et al. Identification of tagitinin C from Tithonia diversifolia as antitrypanosomal compound using bioactivity-guided fractionation. Fitoterapia 2018, 124, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Sesquiterpene lactones from Vernonia cinerascens Sch. Bip. and their in vitro antitrypanosomal activity. Molecules 2018, 23, 248. [Google Scholar] [CrossRef] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Anti-trypanosomatid elemanolide sesquiterpene lactones from Vernonia lasiopus O. Hoffm. Molecules 2017, 22, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimani, N.M.; Matasyoh, J.C.; Kaiser, M.; Brun, R.; Schmidt, T.J. Antiprotozoal Sesquiterpene Lactones and Other Constituents from Tarchonanthus camphoratus and Schkuhria pinnata. J. Nat. Prod. 2018, 81, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Skaf, J.; Hamarsheh, O.; Berninger, M.; Balasubramanian, S.; Oelschlaeger, T.A.; Holzgrabe, U. Improving anti-trypanosomal activity of alkamides isolated from Achillea fragrantissima. Fitoterapia 2018, 125, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Laurella, L.C.; Cerny, N.; Bivona, A.E.; Alberti, A.S.; Giberti, G.; Malchiodi, E.L.; Martino, V.S.; Catalan, C.A.; Alonso, M.R.; Cazorla, S.I.; et al. Assessment of sesquiterpene lactones isolated from Mikania plants species for their potential efficacy against Trypanosoma cruzi and Leishmania sp. PLoS Negl. Trop. Dis. 2017, 11, e0005929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrelli, R.; Ranjbarian, F.; Dall’Acqua, S.; Papa, F.; Iannarelli, R.; Ngahang Kamte, S.L.; Vittori, S.; Benelli, G.; Maggi, F.; Hofer, A.; et al. An overlooked horticultural crop, Smyrnium olusatrum, as a potential source of compounds effective against African trypanosomiasis. Parasitol. Int. 2017, 66, 146–151. [Google Scholar] [CrossRef] [PubMed]
- De Mieri, M.; Monteleone, G.; Ismajili, I.; Kaiser, M.; Hamburger, M. Antiprotozoal Activity-Based Profiling of a Dichloromethane Extract from Anthemis nobilis Flowers. J. Nat. Prod. 2017, 80, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.C.; Souza, R.D.J.; De Moraes, M.H.; Steindel, M.; Biavatti, M.W. A new furanoheliangolide sesquiterpene lactone from Calea pinnatifida (R. Br.) Less. (Asteraceae) and evaluation of its trypanocidal and leishmanicidal activities. J. Braz. Chem. Soc. 2017, 28, 367–375. [Google Scholar] [CrossRef]
- Ulloa, J.L.; Spina, R.; Casasco, A.; Petray, P.B.; Martino, V.; Sosa, M.A.; Frank, F.M.; Muschietti, L.V. Germacranolide-type sesquiterpene lactones from Smallanthus sonchifolius with promising activity against Leishmania mexicana and Trypanosoma cruzi. Parasites Vectors 2017, 10, 567. [Google Scholar] [CrossRef] [PubMed]
- Nyongbela, K.D.; Ntie-Kang, F.; Hoye, T.R.; Efange, S.M.N. Antiparasitic Sesquiterpenes from the Cameroonian Spice Scleria striatinux and Preliminary In Vitro and In Silico DMPK Assessment. Nat. Products Bioprospect. 2017, 7, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elso, O.G.; Bivona, A.E.; Alberti, A.S.; Cerny, N.; Fabian, L.; Morales, C.; Catalán, C.A.N.; Malchiodi, E.L.; Cazorla, S.I.; Sülsen, V.P. Trypanocidal activity of four sesquiterpene lactones isolated from Asteraceae species. Molecules 2020, 25, 2014. [Google Scholar] [CrossRef]
- Gonçalves-Santos, E.; Vilas-Boas, D.F.; Diniz, L.F.; Veloso, M.P.; Mazzeti, A.L.; Rodrigues, M.R.; Oliveira, C.M.; Fernandes, V.H.C.; Novaes, R.D.; Chagas-Paula, D.A.; et al. Sesquiterpene lactone potentiates the immunomodulatory, antiparasitic and cardioprotective effects on anti-Trypanosoma cruzi specific chemotherapy. Int. Immunopharmacol. 2019, 77, 105961. [Google Scholar] [CrossRef] [PubMed]
- Galkina, A.; Krause, N.; Lenz, M.; Daniliuc, C.G.; Kaiser, M.; Schmidt, T.J. Antitrypanosomal Activity of Sesquiterpene Lactones from Helianthus tuberosus L. Including a new furanoheliangolide with an unusual structure. Molecules 2019, 24, 1068. [Google Scholar] [CrossRef] [Green Version]
- Sosa, A.; Salamanca Capusiri, E.; Amaya, S.; Bardón, A.; Giménez-Turba, A.; Vera, N.; Borkosky, S. Trypanocidal activity of South American Vernonieae (Asteraceae) extracts and its sesquiterpene lactones. Nat. Prod. Res. 2021, 35, 5224–5228. [Google Scholar] [CrossRef]
- Elso, O.G.; Clavin, M.; Hernandez, N.; Sgarlata, T.; Bach, H.; Catalan, C.A.N.; Aguilera, E.; Alvarez, G.; Sülsen, V.P. Antiprotozoal Compounds from Urolepis hecatantha (Asteraceae). Evid. -Based Complement. Altern. Med. 2021, 2021, 6622894. [Google Scholar] [CrossRef] [PubMed]
- Milagre, M.M.; Branquinho, R.T.; Gonçalves, M.F.; de Assis, G.; de Oliveira, M.T.; Reis, L.; Saúde-Guimarães, D.A.; de Lana, M. Activity of the sesquiterpene lactone goyazensolide against Trypanosoma cruzi in vitro and in vivo. Parasitology 2020, 147, 108–119. [Google Scholar] [CrossRef]
- Murakami, C.; Cabral, R.S.A.; Gomes, K.S.; Costa-Silva, T.A.; Amaral, M.; Romanelli, M.; Tempone, A.G.; Lago, J.H.G.; da Bolzani, V.S.; Moreno, P.R.H.; et al. Hedyosulide, a novel trypanosomicidal sesterterpene lactone from Hedyosmum brasiliense Mart. ex Miq. Phytochem. Lett. 2019, 33, 6–11. [Google Scholar] [CrossRef]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef] [PubMed]
- Londero, V.S.; Costa-Silva, T.A.; Tempone, A.G.; Namiyama, G.M.; Thevenard, F.; Antar, G.M.; Baitello, J.B.; Lago, J.H.G. Anti-Trypanosoma cruzi activity of costic acid isolated from Nectandra barbellata (Lauraceae) is associated with alterations in plasma membrane electric and mitochondrial membrane potentials. Bioorg. Chem. 2020, 95, 103510. [Google Scholar] [CrossRef]
- Mofidi Tabatabaei, S.; Nejad Ebrahimi, S.; Salehi, P.; Sonboli, A.; Tabefam, M.; Kaiser, M.; Hamburger, M.; Moridi Farimani, M. Antiprotozoal Germacranolide Sesquiterpene Lactones from Tanacetum sonbolii. Planta Med. 2019, 85, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.N.; Just, J.; Dasari, R.; Smith, J.A.; Bissember, A.C.; Kornienko, A.; Rogelj, S. Activity of natural and synthetic polygodial derivatives against Trypanosoma cruzi amastigotes, trypomastigotes and epimastigotes. Nat. Prod. Res. 2021, 35, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Bombaça, A.C.S.; Von Dossow, D.; Barbosa, J.M.C.; Paz, C.; Burgos, V.; Menna-Barreto, R.F.S. Trypanocidal activity of natural sesquiterpenoids involves mitochondrial dysfunction, ROS production and autophagic phenotype in trypanosomacruzi. Molecules 2018, 23, 2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, G.E.G.; Morais, T.R.; Gomes, K.d.S.; Costa-Silva, T.A.; Tempone, A.G.; Lago, J.H.G.; Caseli, L. Antitrypanosomal activity of epi-polygodial from Drimys brasiliensis and its effects in cellular membrane models at the air-water interface. Bioorg. Chem. 2019, 84, 186–191. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.L.; Amaral, M.; Hasegawa, F.R.; Lago, J.H.G.; Tempone, A.G.; Sartorelli, P. (-)-T-Cadinol—A Sesquiterpene Isolated From Casearia sylvestris (Salicaceae)—Displayed In Vitro Activity and Causes Hyperpolarization of the Membrane Potential of Trypanosoma cruzi. Front. Pharmacol. 2021, 12, 734127. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.A.; Ashour, A.A.; Qiu, L. New sesquiterpene glycoside ester with antiprotozoal activity from the flowers of Calendula officinalis L. Nat. Prod. Res. 2021, 35, 5250–5254. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Parai, D.; Dhar, P.; Roy, M.; Barik, R.; Chattopadhyay, S.; Mukherjee, S.K. Andrographolide induces oxidative stress-dependent cell death in unicellular protozoan parasite Trypanosoma brucei. Acta Trop. 2017, 176, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ueno, A.K.; Barcellos, A.F.; Costa-Silva, T.A.; Mesquita, J.T.; Ferreira, D.D.; Tempone, A.G.; Romoff, P.; Antar, G.M.; Lago, J.H.G. Antitrypanosomal activity and evaluation of the mechanism of action of diterpenes from aerial parts of Baccharis retusa (Asteraceae). Fitoterapia 2018, 125, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.C.F.S.; Morais, G.O.; da Silva, M.M.; Kovatch, P.Y.; Ferreira, D.S.; Esperandim, V.R.; Pagotti, M.C.; Magalhães, L.G.; Heleno, V.C.G. In vitro anti-trypanosomal potential of kaurane and pimarane semi-synthetic derivatives. Nat. Prod. Res. 2020, 36, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Kuźma; Kaiser, M.; Wysokińska, H. The production and antiprotozoal activity of abietane diterpenes in Salvia austriaca hairy roots grown in shake flasks and bioreactor. Prep. Biochem. Biotechnol. 2017, 47, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.S.; Da Costa, F.B.; Brun, R.; Kaiser, M.; Schmidt, T.J. ent-pimarane and ent-kaurane diterpenes from Aldama discolor (Asteraceae) and their antiprotozoal activity. Molecules 2016, 21, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirttimaa, M.; Nasereddin, A.; Kopelyanskiy, D.; Kaiser, M.; Yli-Kauhaluoma, J.; Oksman-Caldentey, K.M.; Brun, R.; Jaffe, C.L.; Moreira, V.M.; Alakurtti, S. Abietane-Type Diterpenoid Amides with Highly Potent and Selective Activity against Leishmania donovani and Trypanosoma cruzi. J. Nat. Prod. 2016, 79, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Farimani, M.M.; Khodaei, B.; Moradi, H.; Aliabadi, A.; Ebrahimi, S.N.; De Mieri, M.; Kaiser, M.; Hamburger, M. Phytochemical Study of Salvia leriifolia Roots: Rearranged Abietane Diterpenoids with Antiprotozoal Activity. J. Nat. Prod. 2018, 81, 1384–1390. [Google Scholar] [CrossRef] [PubMed]
- Zadali, R.; Nejad Ebrahimi, S.; Tofighi, Z.; Es-haghi, A.; Hamburger, M.; Kaiser, M.; D’ Ambola, M.; De Tommasi, N.; Hadjiakhoondi, A. Antiprotozoal activity of diterpenoids isolated from Zhumeria majdae- absolute configuration by circular dichroism. DARU J. Pharm. Sci. 2020, 28, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Tabefam, M.; Farimani, M.M.; Danton, O.; Ramseyer, J.; Kaiser, M.; Ebrahimi, S.N.; Salehi, P.; Batooli, H.; Potterat, O.; Hamburger, M. Antiprotozoal Diterpenes from Perovskia abrotanoides. Planta Med. 2018, 84, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Nvau, J.B.; Alenezi, S.; Ungogo, M.A.; Alfayez, I.A.M.; Natto, M.J.; Gray, A.I.; Ferro, V.A.; Watson, D.G.; de Koning, H.P.; Igoli, J.O. Antiparasitic and Cytotoxic Activity of Bokkosin, A Novel Diterpene-Substituted Chromanyl Benzoquinone From Calliandra portoricensis. Front. Chem. 2020, 8, 574103. [Google Scholar] [CrossRef] [PubMed]
- Anyam, J.V.; Daikwo, P.E.; Ungogo, M.A.; Nweze, N.E.; Igoli, N.P.; Gray, A.I.; De Koning, H.P.; Igoli, J.O. Two New Antiprotozoal Diterpenes From the Roots of Acacia nilotica. Front. Chem. 2021, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Catteau, L.; Schioppa, L.; Beaufay, C.; Girardi, C.; Hérent, M.F.; Frédérich, M.; Quetin-Leclercq, J. Antiprotozoal activities of Triterpenic Acids and Ester Derivatives Isolated from the Leaves of Vitellaria paradoxa. Planta Med. 2021, 87, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Labib, R.; Ebada, S.; Youssef, F.; Ashour, M.; Ross, S. Ursolic acid, a natural pentacylcic triterpene from Ochrosia elliptica and its role in the management of certain neglected tropical diseases. Pharmacogn. Mag. 2016, 12, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.G.; Ali, Z.; Fantoukh, O.; Raman, V.; Kamdem, R.S.T.; Khan, I. Glycosides of ursane-type triterpenoid, benzophenone, and iridoid from Vangueria agrestis (Fadogia agrestis) and their anti-infective activities. Nat. Prod. Res. 2020, 34, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.S.; Barbosa-Filho, J.M.; Lanfredi-Rangel, A.; Guimarães, E.T.; Moreira, D.R.M.; Soares, M.B.P. Antiparasitic evaluation of betulinic acid derivatives reveals effective and selective anti-Trypanosoma cruzi inhibitors. Exp. Parasitol. 2016, 166, 108–115. [Google Scholar] [CrossRef]
- Sousa, P.L.; da Silva Souza, R.O.; Tessarolo, L.D.; de Menezes, R.R.P.P.B.; Sampaio, T.L.; Canuto, J.A.; Martins, A.M.C. Betulinic acid induces cell death by necrosis in Trypanosoma cruzi. Acta Trop. 2017, 174, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Steverding, D.; Sidjui, L.S.; Ferreira, É.R.; Ngameni, B.; Folefoc, G.N.; Mahiou-Leddet, V.; Ollivier, E.; Stephenson, G.R.; Storr, T.E.; Tyler, K.M. Trypanocidal and leishmanicidal activity of six limonoids. J. Nat. Med. 2020, 74, 606–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carothers, S.; Nyamwihura, R.; Collins, J.; Zhang, H.; Park, H.; Setzer, W.N.; Ogungbe, I.V. Bauerenol acetate, the pentacyclic triterpenoid from Tabernaemontana longipes, is an antitrypanosomal agent. Molecules 2018, 23, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabefam, M.; Farimani, M.M.; Danton, O.; Ramseyer, J.; Nejad Ebrahimi, S.; Neuburger, M.; Kaiser, M.; Salehi, P.; Potterat, O.; Hamburger, M. Antiprotozoal Isoprenoids from Salvia hydrangea. J. Nat. Prod. 2018, 81, 2682–2691. [Google Scholar] [CrossRef] [PubMed]
- Muganza, D.M.; Fruth, B.; Nzunzu, J.L.; Tuenter, E.; Foubert, K.; Cos, P.; Maes, L.; Kanyanga, R.C.; Exarchou, V.; Apers, S.; et al. In vitro antiprotozoal activity and cytotoxicity of extracts and isolated constituents from Greenwayodendron suaveolens. J. Ethnopharmacol. 2016, 193, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Szabó, L.U.; Kaiser, M.; Mäser, P.; Schmidt, T.J. Antiprotozoal nor-triterpene alkaloids from Buxus sempervirens L. Antibiotics 2021, 10, 696. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2018, 18, 41–58. [Google Scholar] [CrossRef]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G.G. Application of sesquiterpene lactone: A new promising way for cancer therapy based on anticancer activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef]
- Moujir, L.; Callies, O.; Sousa, P.M.C.; Sharopov, F.; Seca, A.M.L. Applications of sesquiterpene lactones: A review of some potential success cases. Appl. Sci. 2020, 10, 3001. [Google Scholar] [CrossRef]
Figure 1.
The life cycle of T. brucei.

Figure 2.
The life cycle of T. cruzi.

Figure 3.
Structures of monoterpenes (1–6) and iridoids (7–9).

Figure 4.
Structures of sesquiterpenes 10–27.

Figure 5.
Structures of sesquiterpenes 28–44.

Figure 6.
Structures of sesquiterpenes 45–50.

Figure 7.
Structures of sesquiterpenes 51–58.

Figure 8.
Structures of sesquiterpenes 59–71.

Figure 9.
Structures of sesquiterpenes 72–87.

Figure 10.
Structures of diterpenes 88–97.

Figure 11.
Structures of diterpenes 98–104.

Figure 12.
Structures of diterpenes 105–115.

Figure 13.
Structures of triterpenes 116–121.

Figure 14.
Structures of triterpenes 122–135.

Figure 15.
Structures of triterpenes 136–150.

Figure 16.
Promising hit compounds for further research into treating Human African Trypanosomiasis.
Figure 16.
Promising hit compounds for further research into treating Human African Trypanosomiasis.

Figure 17.
Promising hit compounds for further research into treating Human American Trypanosomiasis. All the selected compounds were tested against T. cruzi intracellular amastigote forms.
Figure 17.
Promising hit compounds for further research into treating Human American Trypanosomiasis. All the selected compounds were tested against T. cruzi intracellular amastigote forms.


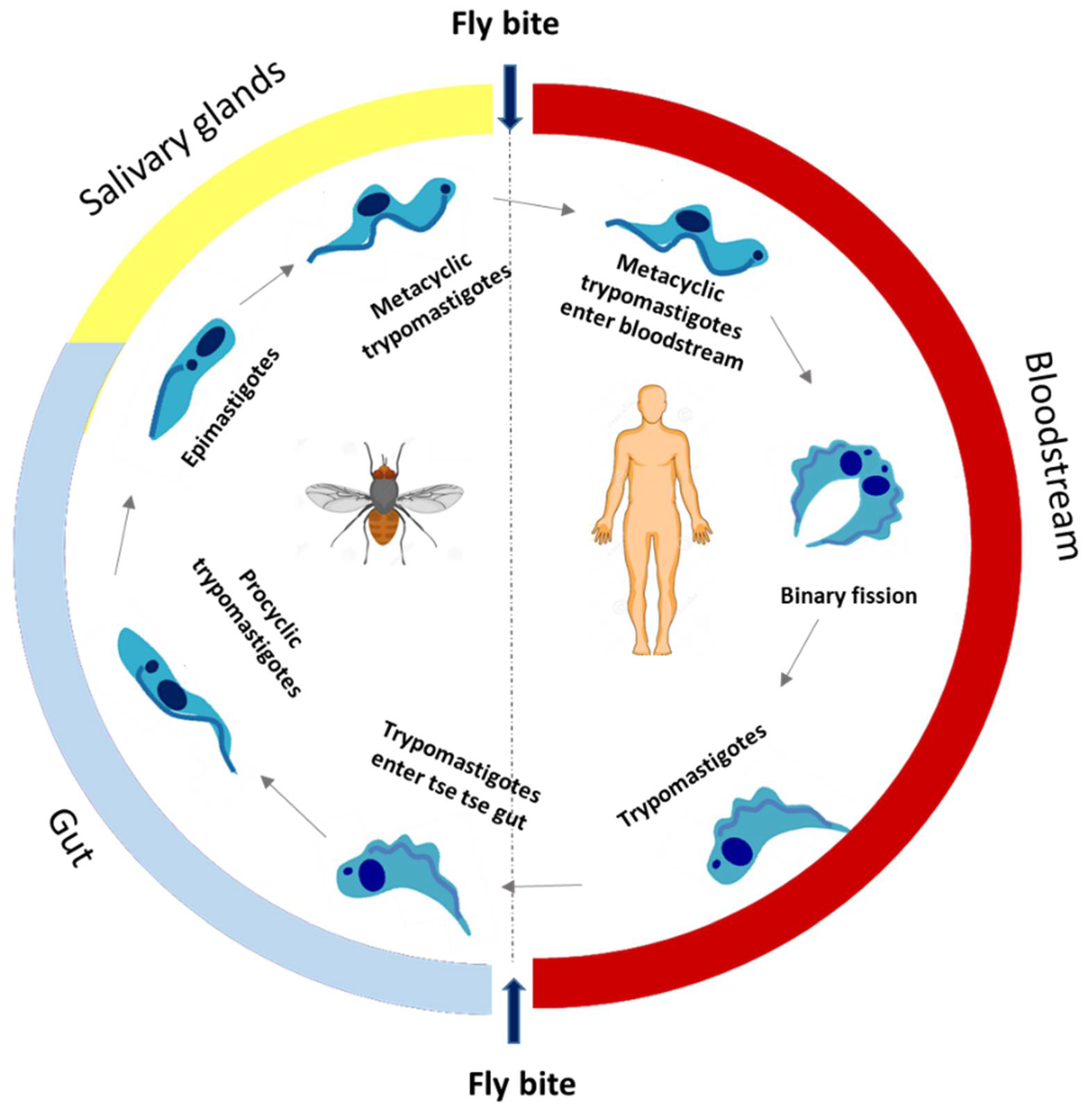{kind=link}
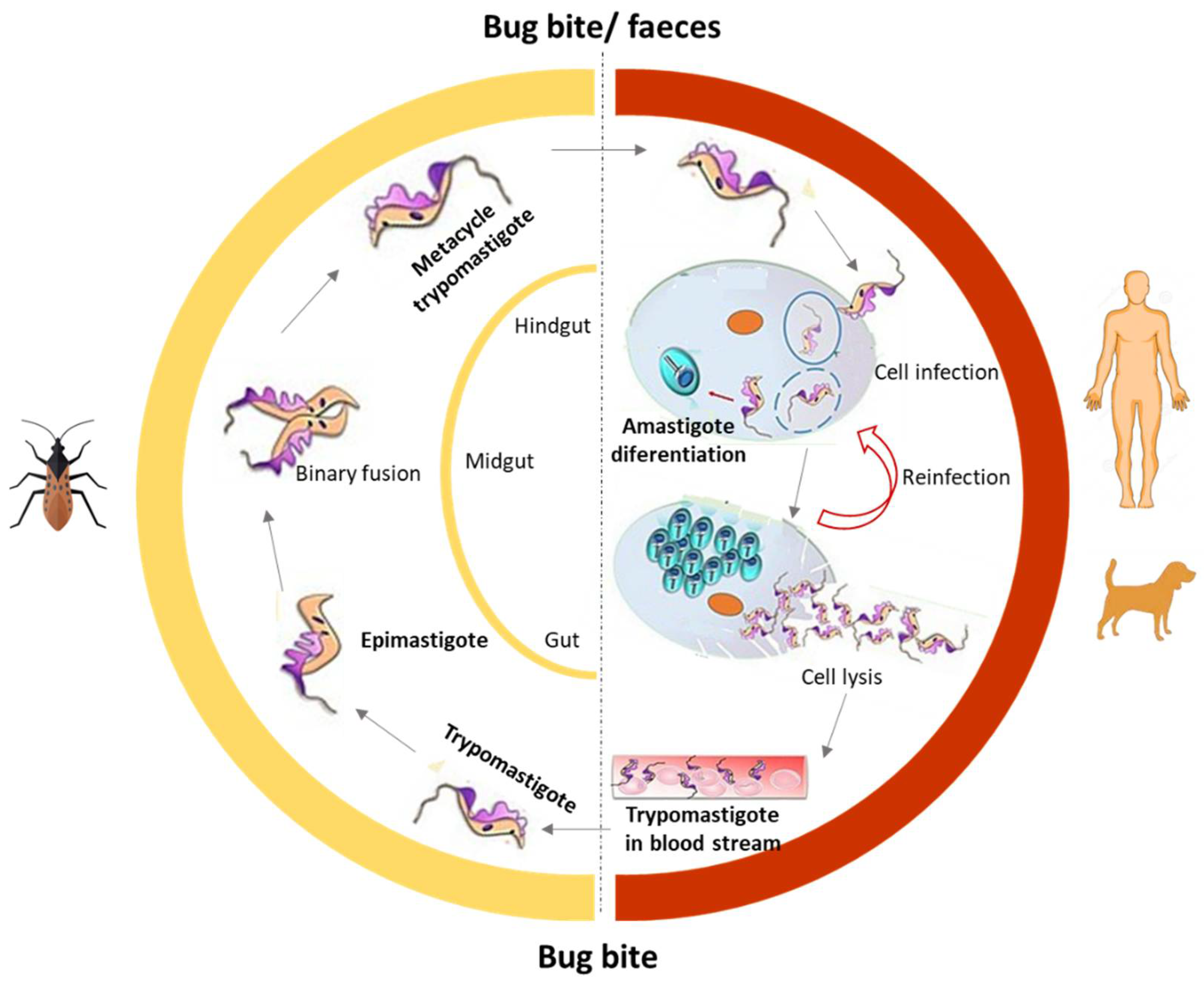{kind=link}
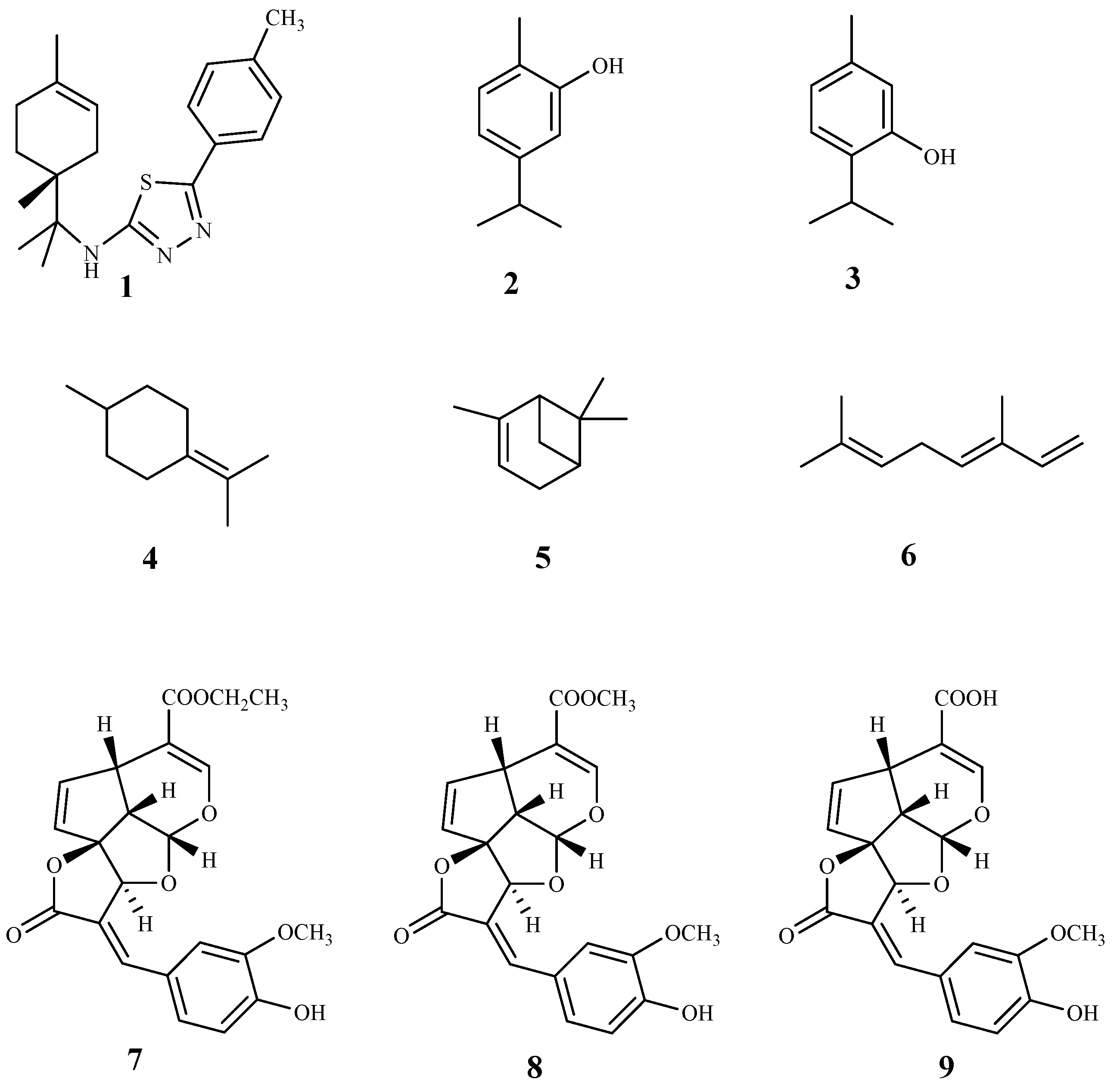{kind=link}
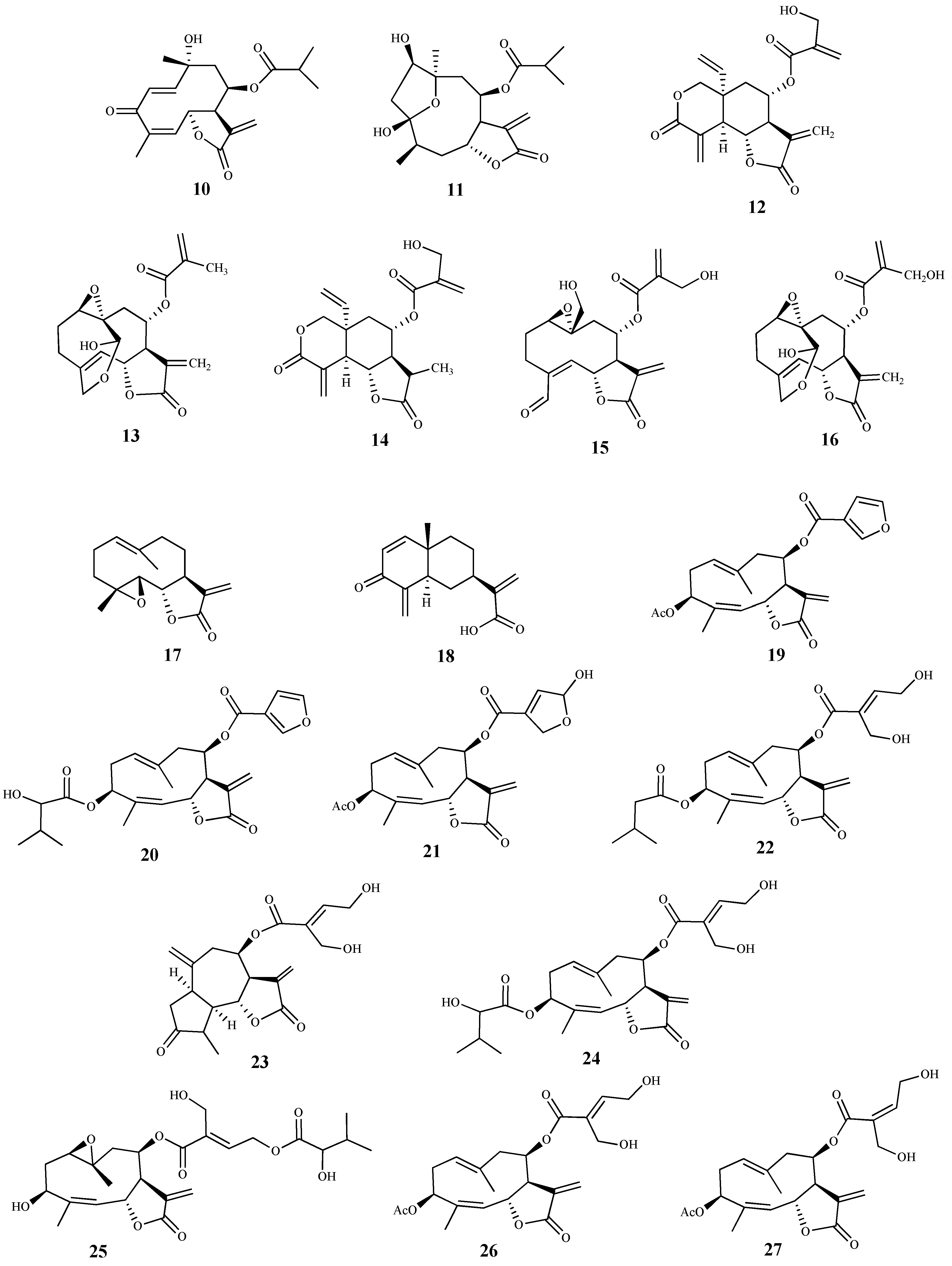{kind=link}
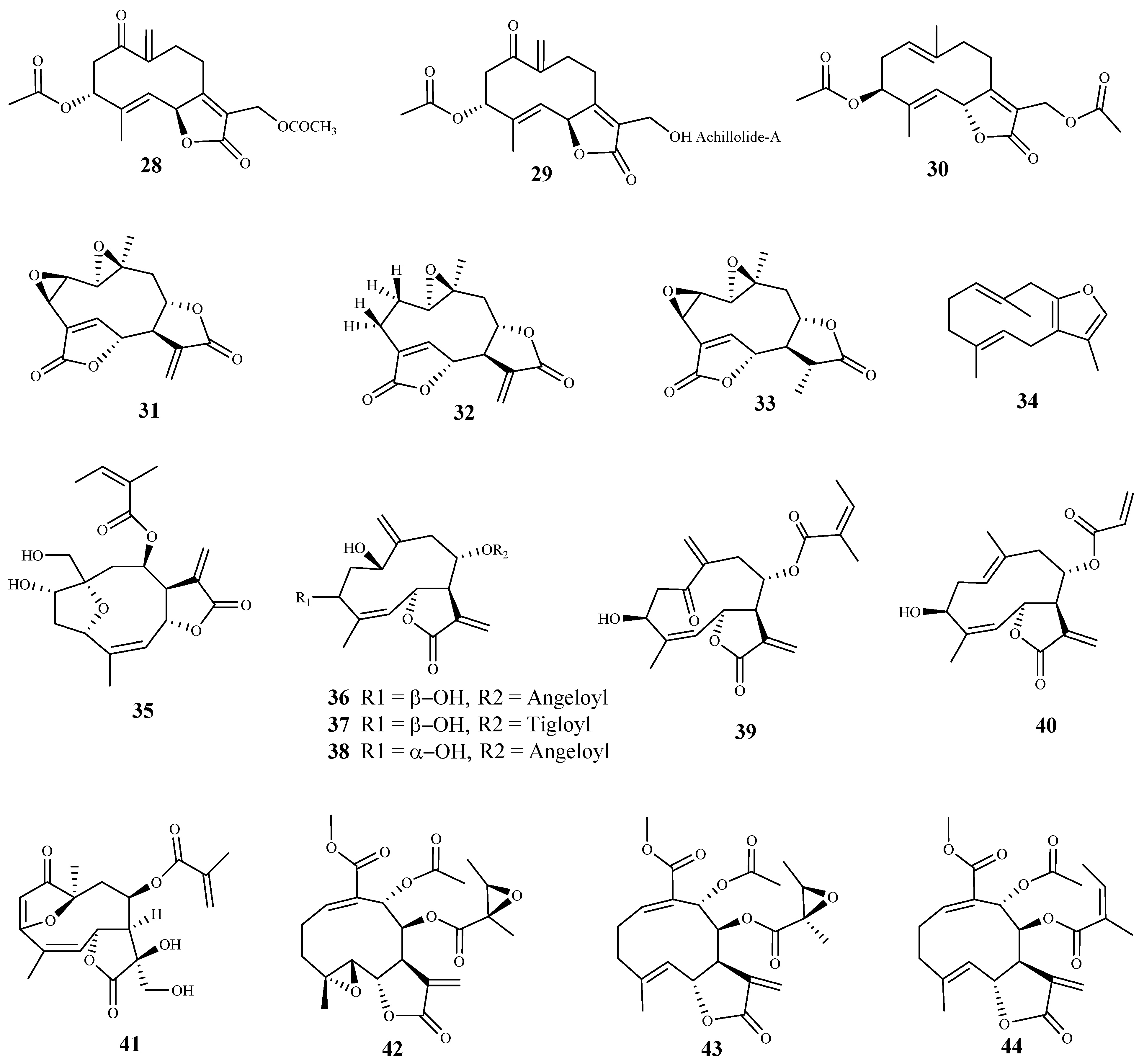{kind=link}
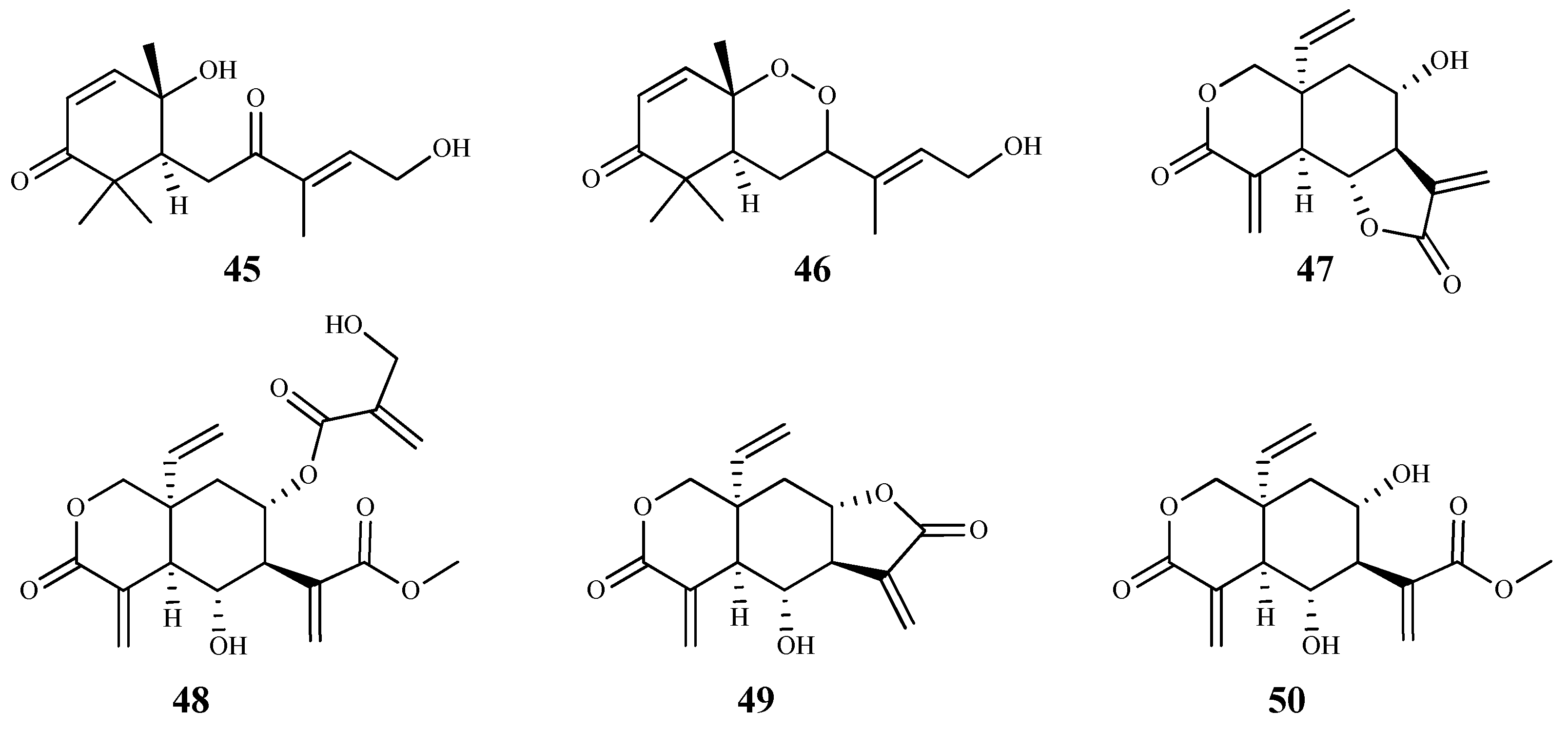{kind=link}
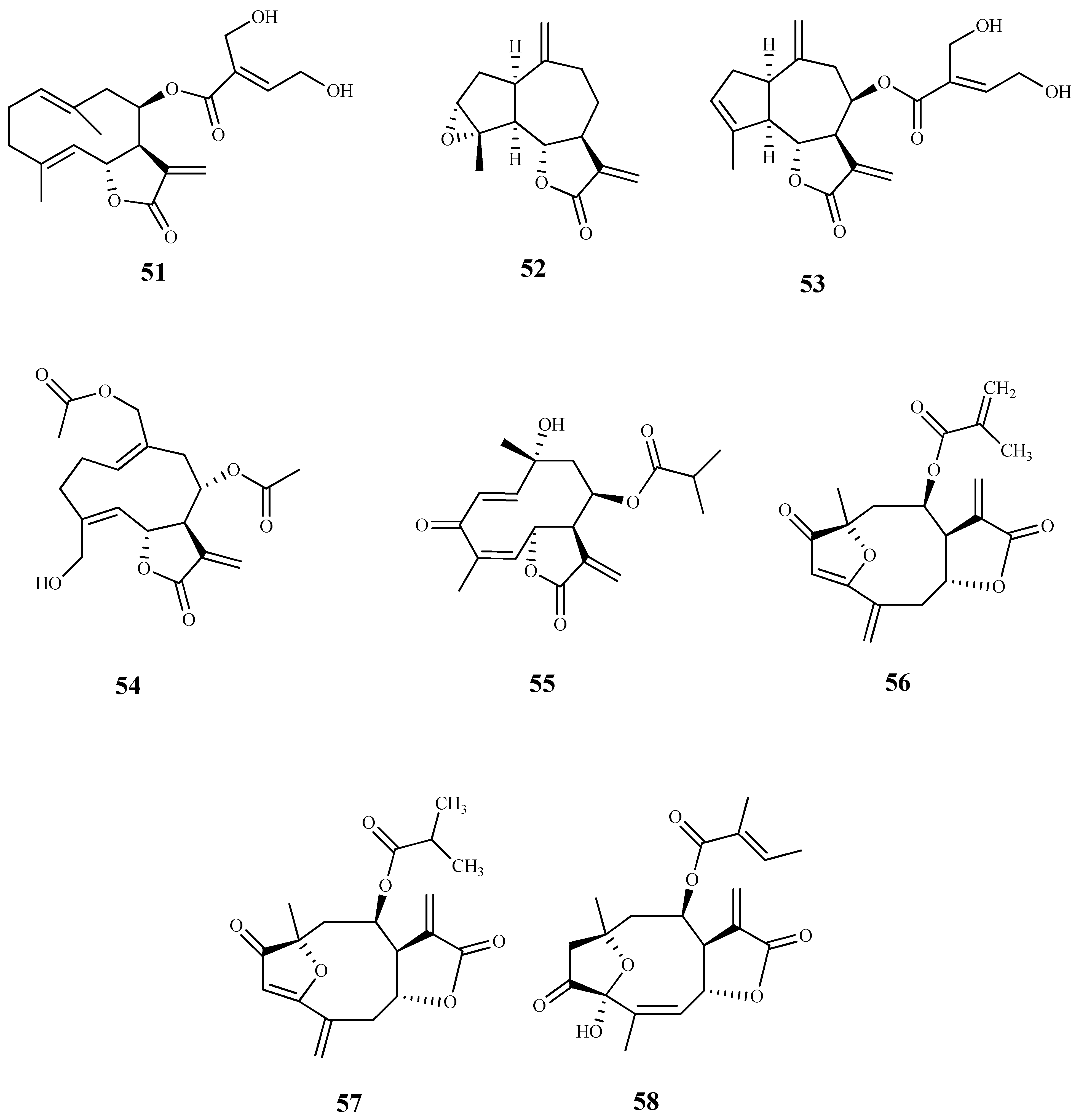{kind=link}
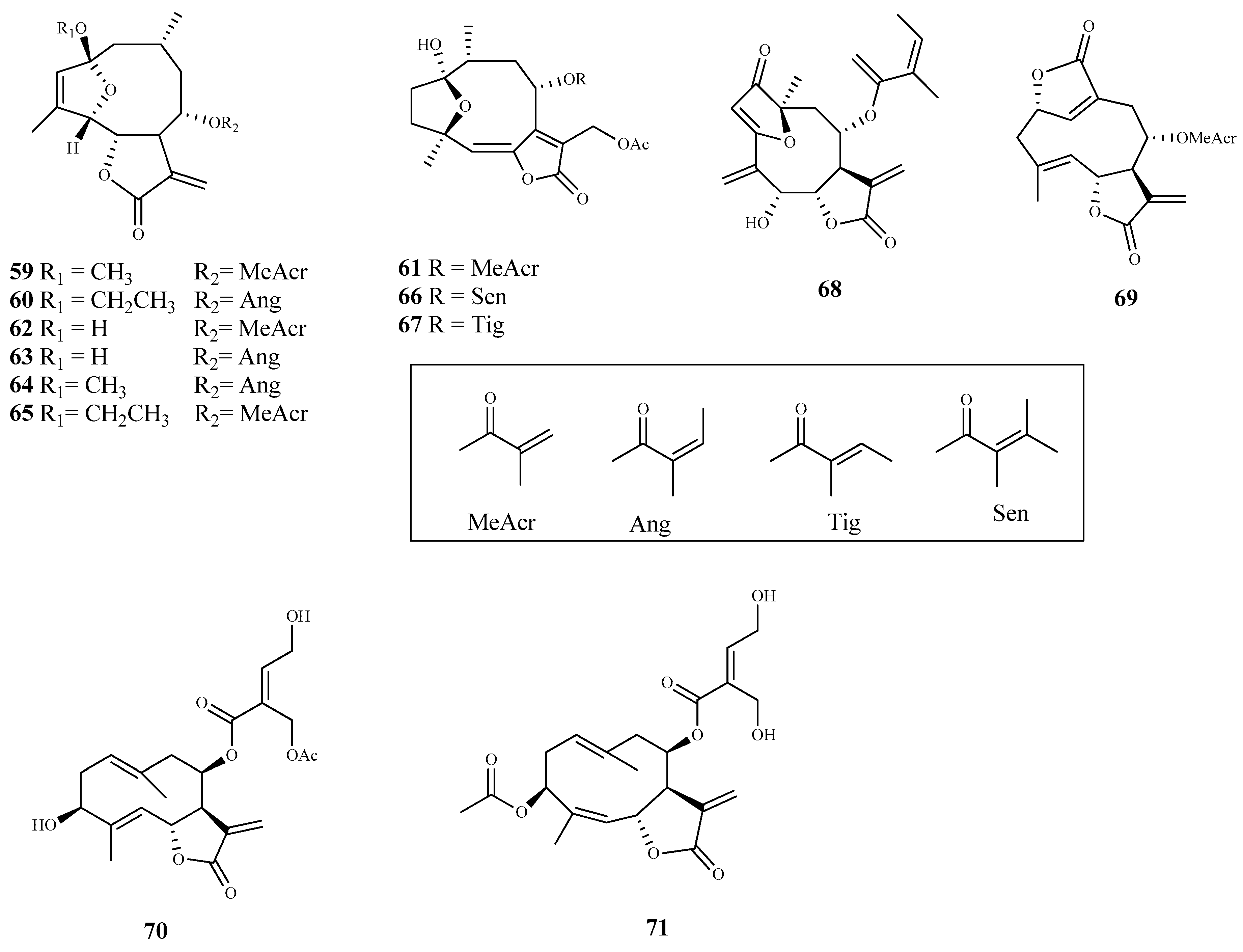{kind=link}
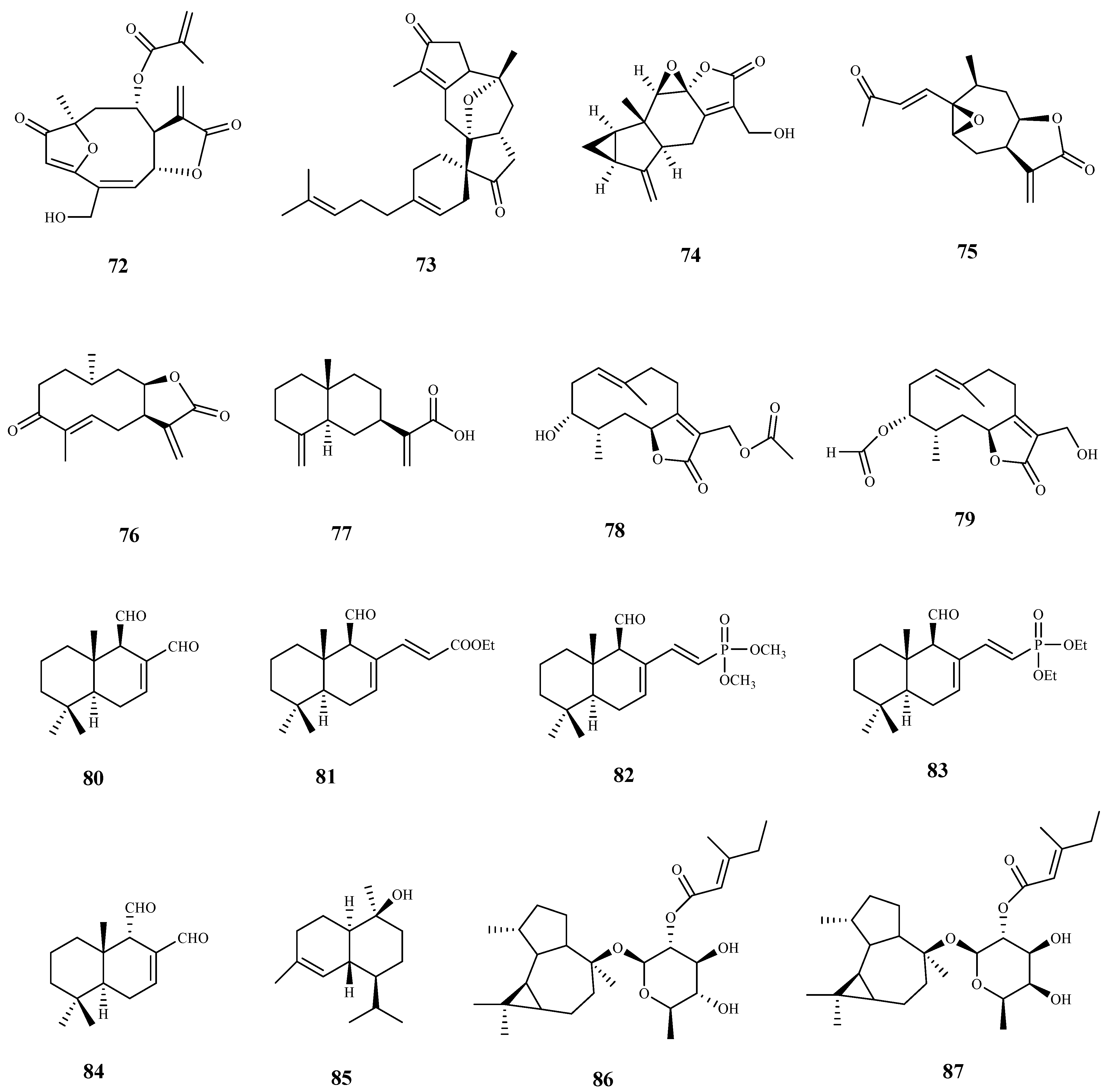{kind=link}
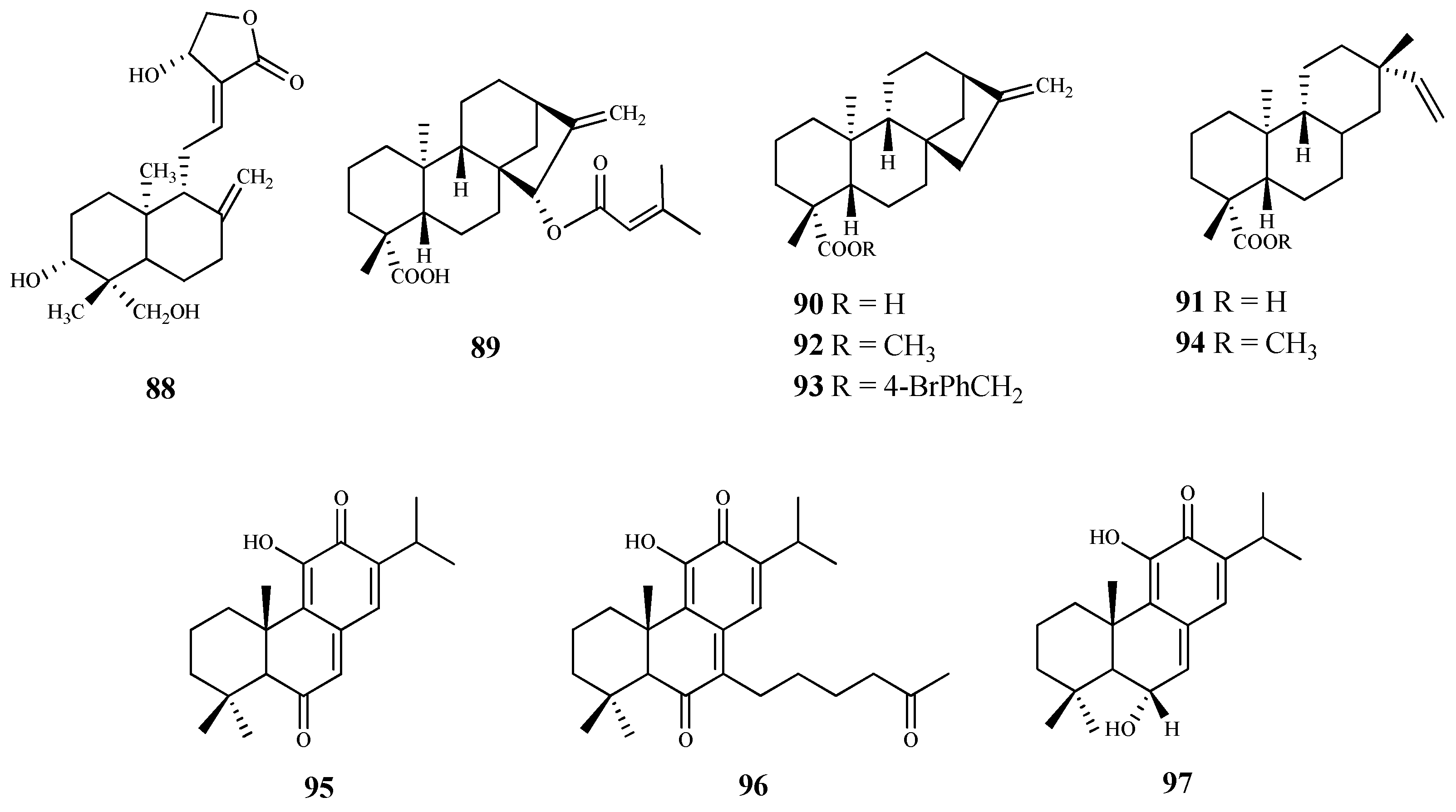{kind=link}
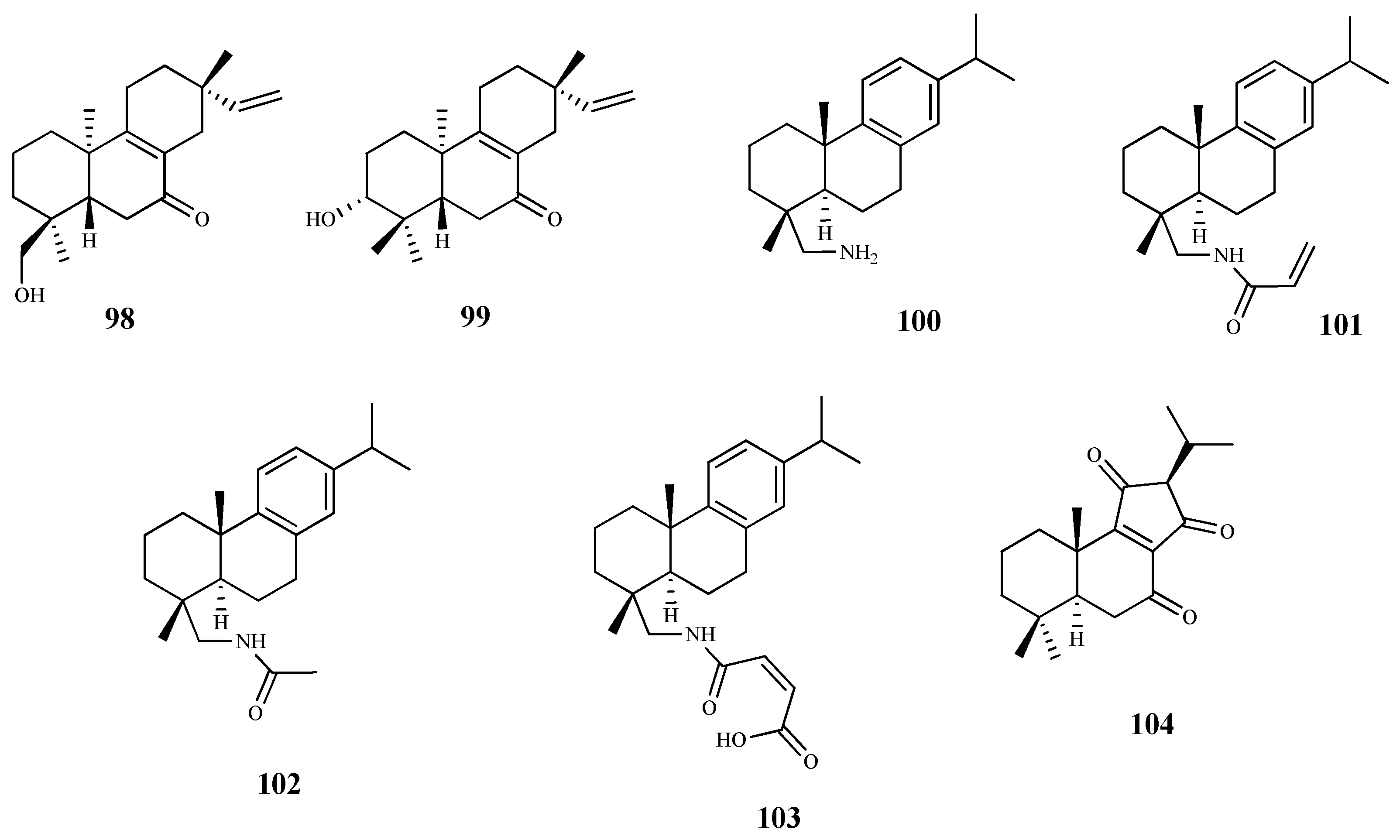{kind=link}
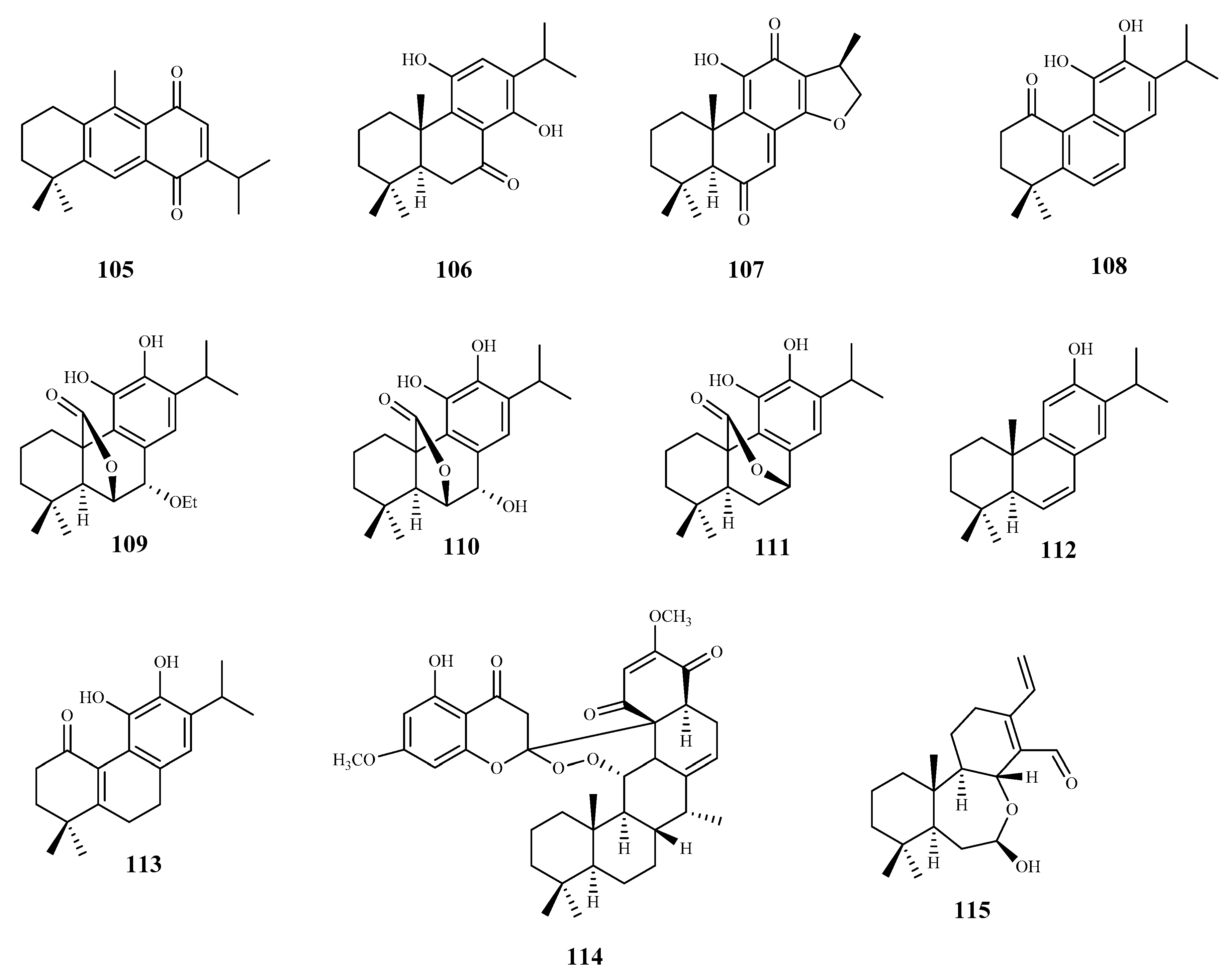{kind=link}
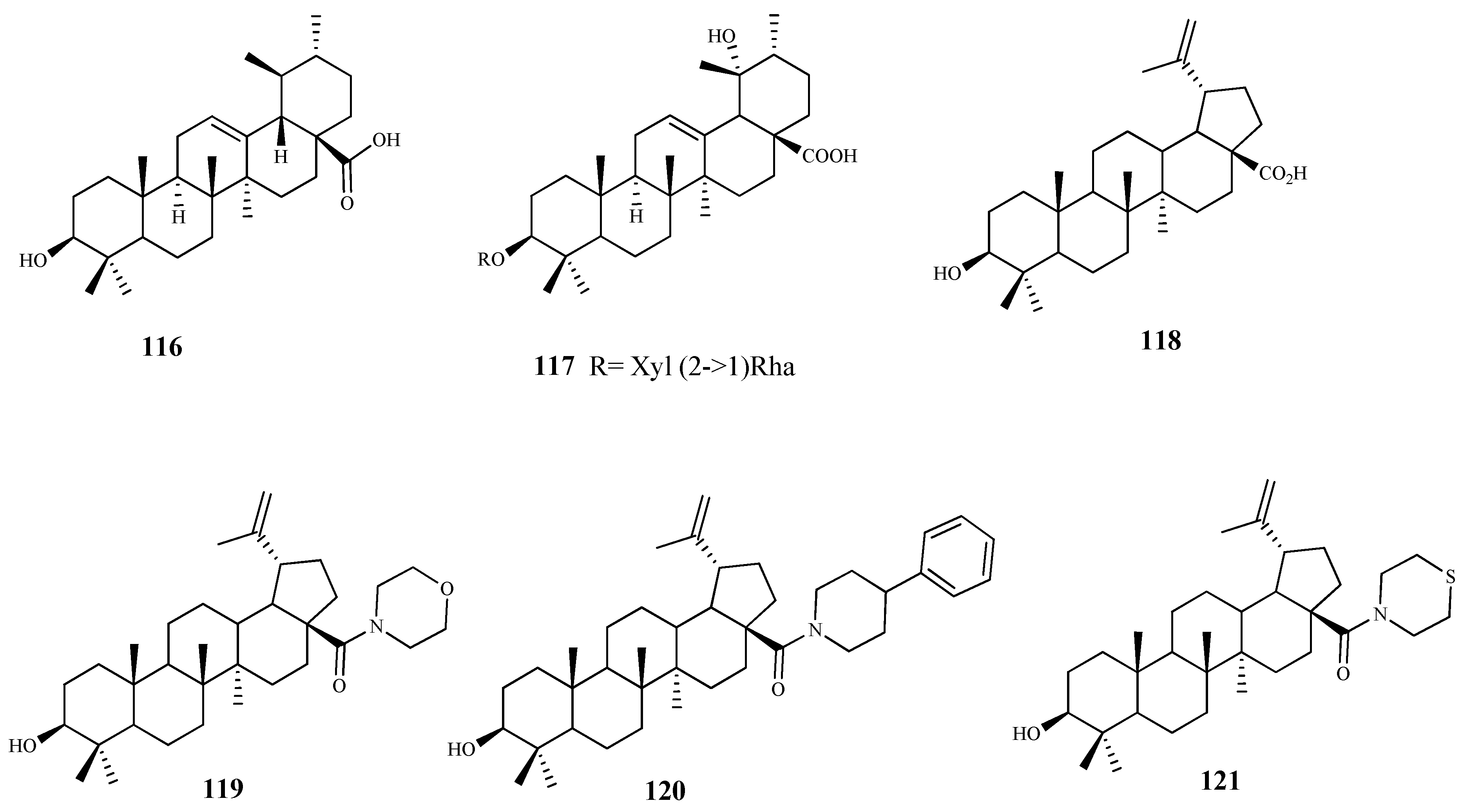{kind=link}
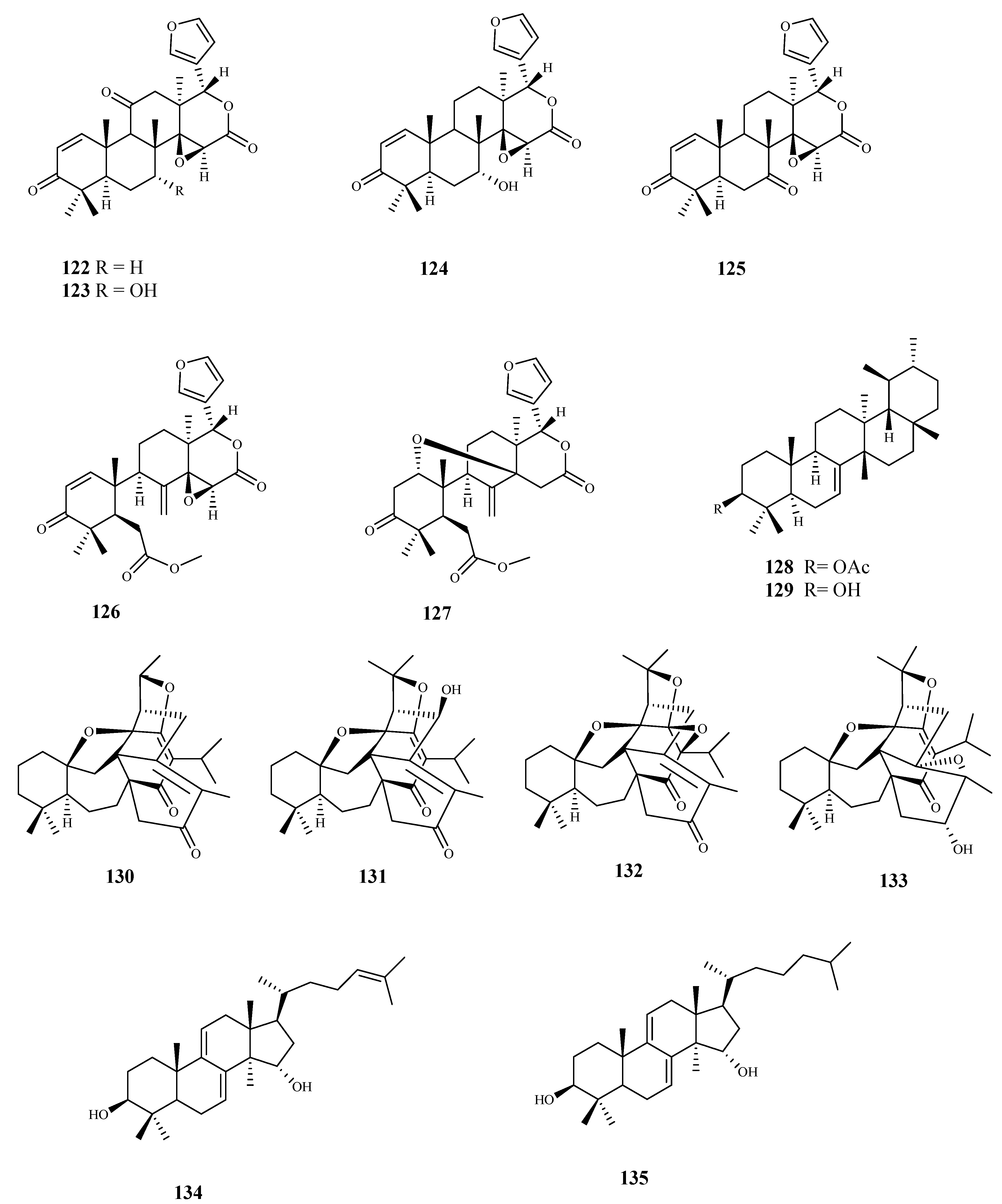{kind=link}
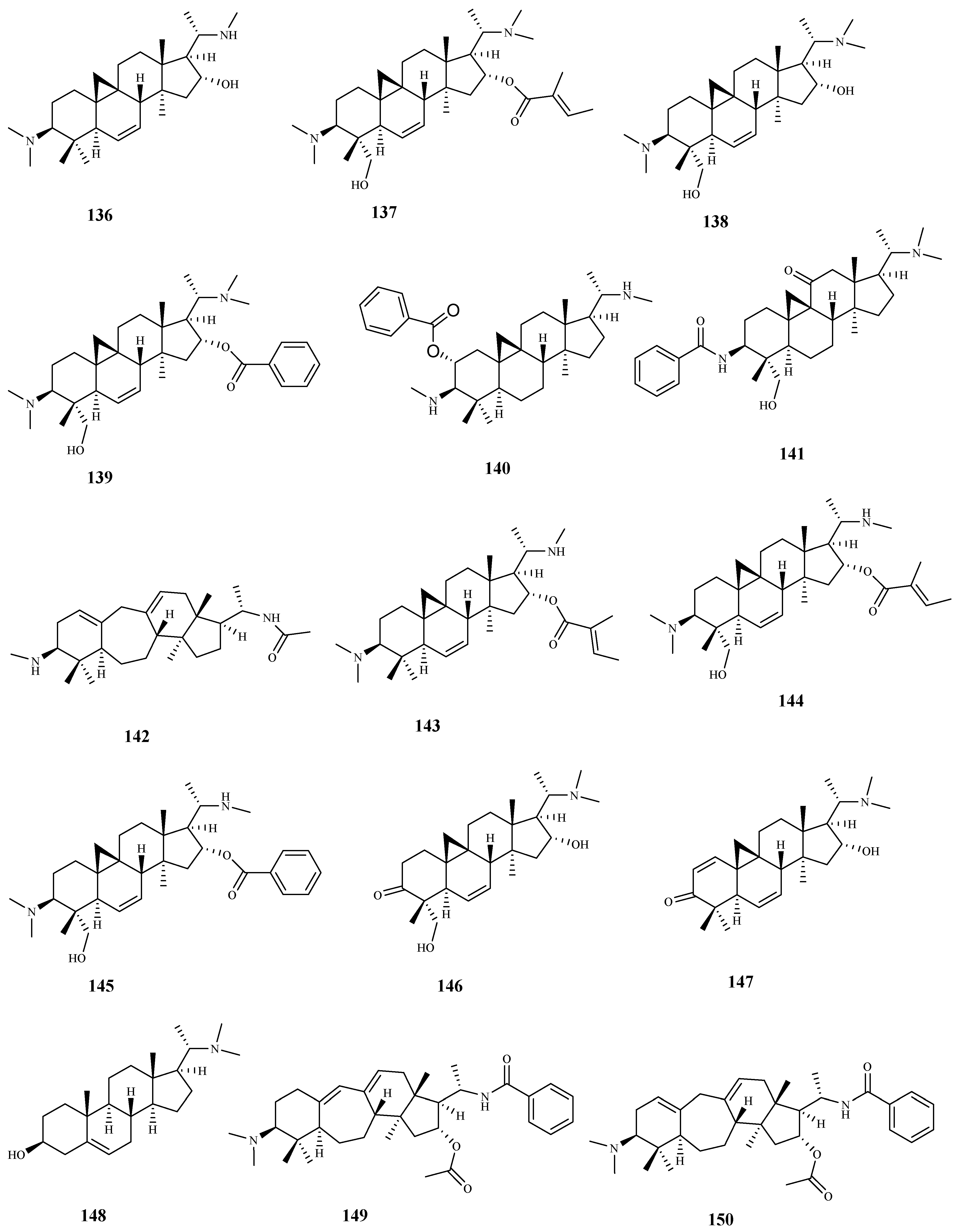{kind=link}
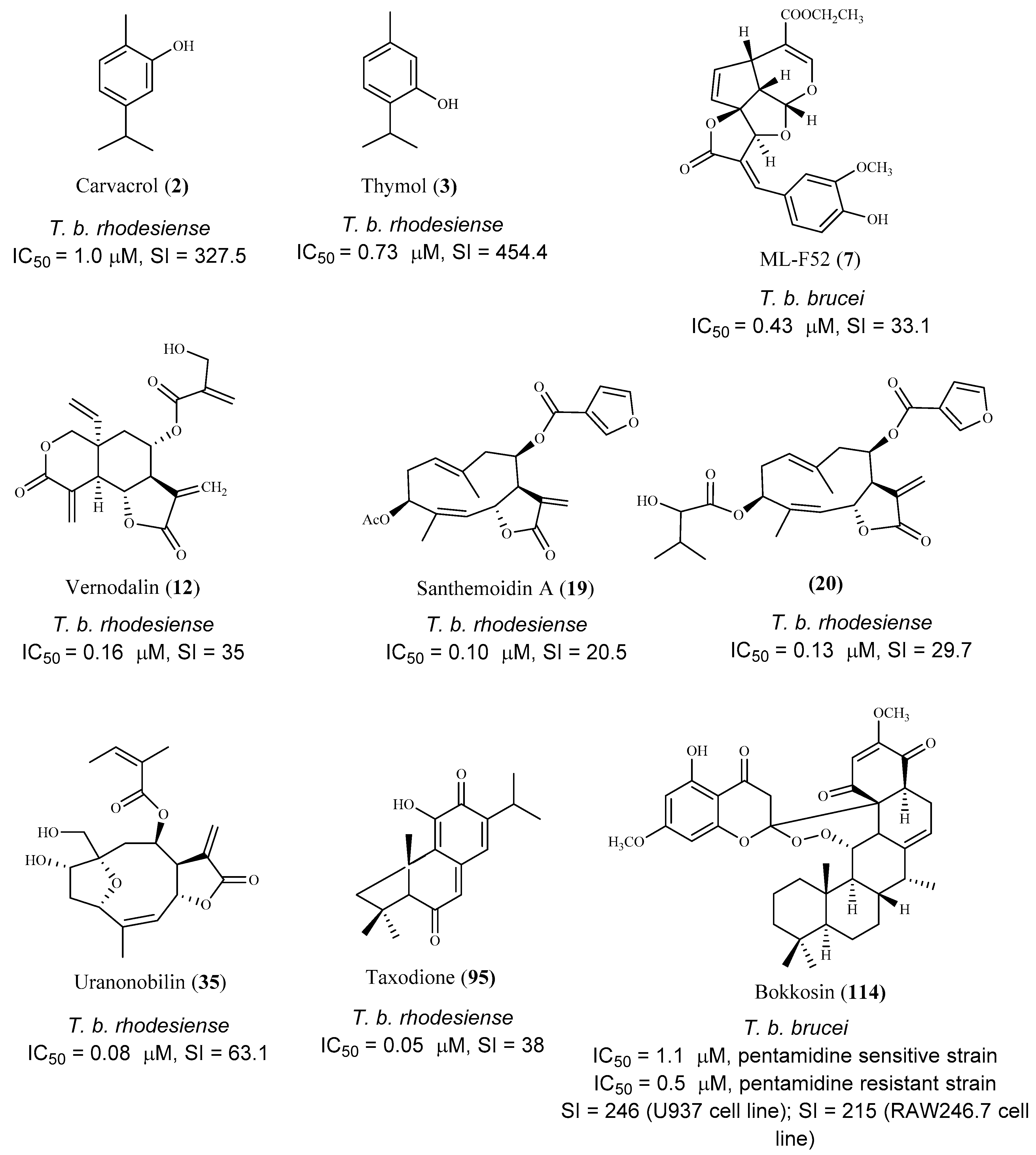{kind=link}
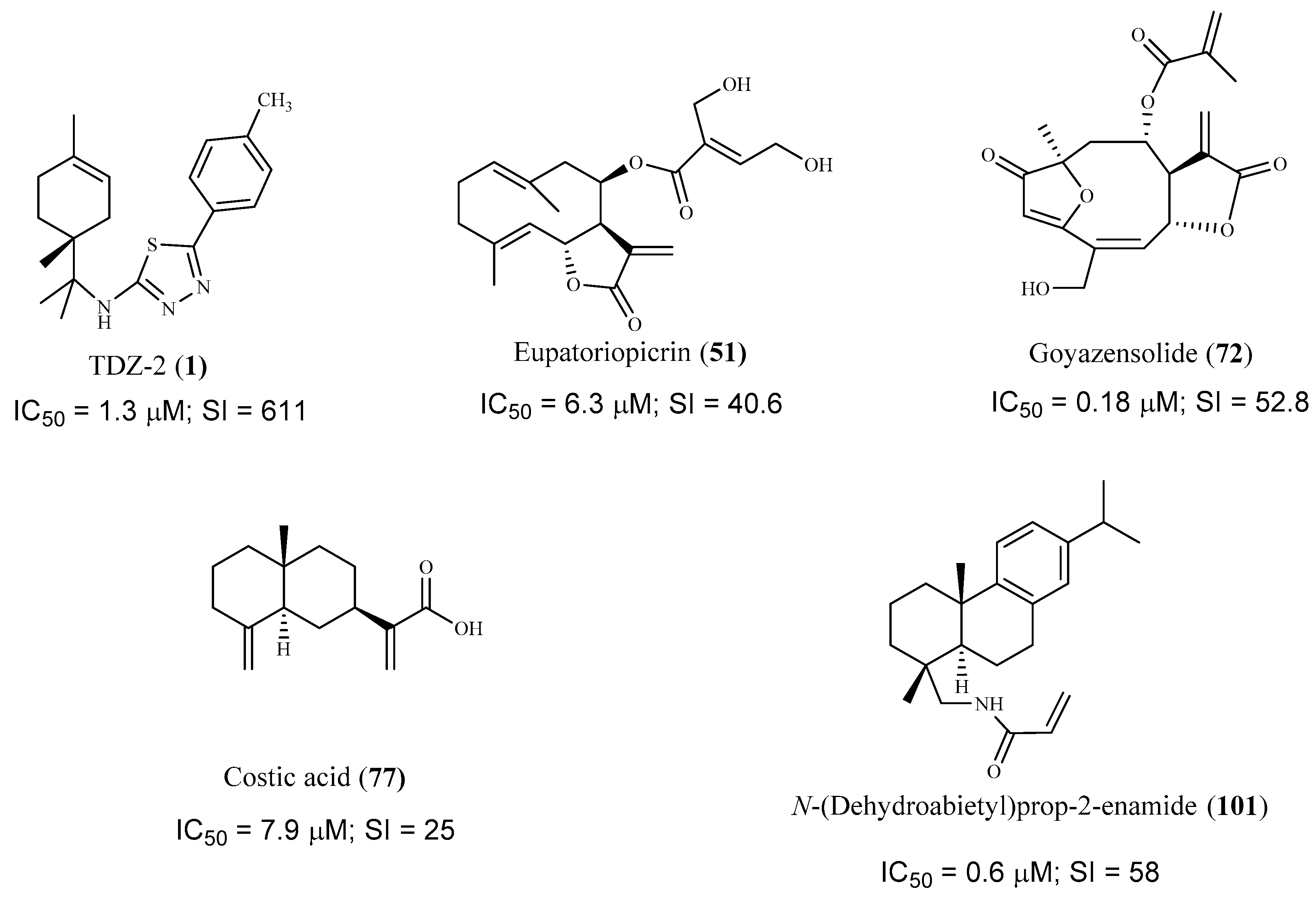{kind=link}
Table 1.
Plant terpenoids with anti-T. brucei activity (2016–2021).
| Compound a | Plant | Parasite (Form) b | IC50 (µM) | SI | Refs. |
|---|---|---|---|---|---|
| Monoterpenes and Iridoids | |||||
| carvacrol (2) | Origanum onites L. | T. b. rhodesiense (tryp.) | 1.0 | 327.5 | [54] |
| thymol (3) | Origanum onites L. | T. b. rhodesiense (tryp.) | 0.73 | 454.4 | [54] |
| terpinolene (4) | Crithmum maritimum L. | T. brucei (tryp.) | 0.26 | 180.0 | [55] |
| α-pinene (5) | Crithmum maritimum L. | T. brucei (tryp.) | 7.4 | >100 | [55] |
| β-ocimene (6) | Crithmum maritimum L. | T. brucei (tryp.) | 8.0 | >91 | [55] |
| ML-F52 (7) | Morinda lucida Benth. | T. b. brucei (tryp.) | 0.43 | 33.1 | [56] |
| molucidin (8) | Morinda lucida Benth. | T. b. brucei (tryp.) | 1.27 | 3.7 | [56] |
| ML-2-3 (9) | Morinda lucida Benth. | T. b. brucei (tryp.) | 3.75 | >13.3 | [56] |
| Sesquiterpenes | |||||
| tagitinin C (10) | Tithonia diversifolia (Hemsl) A. Grey | T. brucei (tryp.) | 0.012 | 3.0 | [57] |
| tagitinin A (11) | Tithonia diversifolia (Hemsl) A. Grey | T. brucei (tryp.) | 0.97 | 1.3 | [57] |
| vernodalin (12) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 0.16 | 35.0 | [58,59] |
| 11β, 13-dihydrovernolide (13) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 0.5 | 13.0 | [58,59] |
| 11β, 13-dihydrovernodaline (14) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 1.1 | 4.2 | [58,59] |
| vernocinerascolide (15) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 4.8 | 27.0 | [58,59] |
| 11β, 13-dihydrohydroxtvernolide (16) | Vernonia cinerascens Sch.Bip. | T. b. rhodesiense (tryp.) | 5.0 | 4.3 | [58,59] |
| parthenolide (17) | Tarchonanthus camphoratus L. | T. b. rhodesiense (tryp.) | 0.39 | 18.6 | [60] |
| 3-oxo-1,2-dehydrocostic acid (18) | Tarchonanthus camphoratus L. | T. b. rhodesiense (tryp.) | 2.8 | 6.2 | [60] |
| santhemoidin A (19) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.10 | 20.5 | [60] |
| 3β-(2″-hydroxyisovaleroyloxy)-8β-(3-furoyloxy)costunolide (20) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.13 | 29.7 | [60] |
| 2″-dehydroeucannabinolidesemiacetal (21) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.35 | 11.5 | [60] |
| 3-desacetyl-3-isovaleroyleu-cannabinolide (22) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.52 | 13.0 | [60] |
| 3-oxo-4β,15-dihydroliqustrin-[4′,5′-dihydroxytigloyloxy] (23) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.60 | 19.2 | [60] |
| schkuhrin II (24) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.82 | 13.4 | [60] |
| 1(10)-epoxy-3β-hydroxy-8β-[5′-hydroxy-4′-(2″- hydroxyisovaleroyloxy)tigloyloxy]costunolide (25) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.91 | 15.8 | [60] |
| eucannabinolide (26) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 0.92 | 15.8 | [60] |
| 2′(3′)-Z-eucannabinolide (27) | Schkuhria pinnata (Lam.) Kuntze ex Thell. | T. b. rhodesiense (tryp.) | 1.7 | 31.1 | [60] |
| 28 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 3.03 | n.d | [61] |
| 29 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 10.97 | n.d | [61] |
| 30 | Achillea fragrantissima (Forssk.) Sch.Bip. | T. b. brucei (tryp.) | 10.97 | n.d | [61] |
| isofuranodiene (34) | Smyrnium olusatrum L. | T. brucei (tryp.) | 3.0 | 30.0 | [63] |
| furanonobilin (35) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.08 | 63.1 | [64] |
| hydroxyisonobilin (36) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.61 | 8.3 | [64] |
| 8-tigloylhydroxyisonobilin (37) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.36 | 14.1 | [64] |
| 3-epi-hydroxyisonobilin (38) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.88 | 8.3 | [64] |
| nobilinon A (39) | Anthemis nobilis L. | T. b. rhodesiense (tryp.) | 0.4 | 3.8 | [64] |
| 45 | Scleria striatinux De Wild. | T. b. rhodesiense (tryp.) | 0.002 | 8.3 | [67] |
| 46 | Scleria striatinux De Wild. | T. b. rhodesiense (tryp.) | 0.025 | 3.4 | [67] |
| vernolepin (47) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.185 | 14.5 | [59] |
| vernodalol (48) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.26 | 14.4 | [59] |
| vernomenin (49) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 0.51 | 4.5 | [59] |
| 8-desacylvernodalol (50) | Vernonia lasiopus (O.Hoffm.) H.Rob. | T. b. rhodesiense (tryp.) | 2.53 | 13.7 | [59] |
| 4,15-iso-atriplicolide methacrylate (56) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.077 | 6.7 | [70] |
| 4,15-iso-atriplicolide isobutryrate (57) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.26 | 3.38 | [70] |
| heliantuberolide-8-O-tiglate is (58) | Helianthus tuberosus L. | T. b. rhodesiense (tryp.) | 0.92 | 4.24 | [70] |
| 78 | Tanacetum sonbolii Mozaff. | T. b. rhodesiense (tryp.) | 5.1 | 3.9 | [77] |
| 79 | Tanacetum sonbolii Mozaff. | T. b. rhodesiense (tryp.) | 10.2 | 4.0 | [77] |
| Diterpenes | |||||
| andrographolide (88) | Andrographis paniculate (Burm. F.) Wall. Ex Nees | T. brucei procyclic trypomastigotes | 8.3 | 8.5 | [83] |
| taxodione (95) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 0.05 | 38.0 | [86] |
| 7-(20-oxohexyl)-taxodione (96) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 0.62 | 5.0 | [86] |
| taxodone (97) | Salvia austriaca Jacq. | T. b. rhodesiense (tryp.) | 1.67 | 2.4 | [86] |
| ent-7-oxo-pimara-8,15-diene-18-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | T. b. rhodesiense (tryp.) | 24.3 | 2.0 | [87] |
| leriifolione (104) | Salvia leriifolia Benth. | T. b. rhodesiense (tryp.) | 1.0 | 2.6 | [89] |
| 12, 16-dideoxy aegyptinone B (105) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 3.6 | 1.7 | [90] |
| 11,14-dihydroxy-8, 11,13- abietatrien-7-one (106) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 1.8 | 21.9 | [90] |
| lanugon Q (107) | Zhumeria majdae Rech. F | T. b. rhodesiense (tryp.) | 0.1 | 15.4 | [90] |
| miltiodiol (108) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 0.5 | 10.5 | [91] |
| 7α-ethoxyrosmanol (109) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) (trypomastigote) | 0.8 | 14.9 | [91] |
| rosmanol (110) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 3.8 | 1.5 | [91] |
| carnosol (111) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 5.4 | 2.4 | [91] |
| Δ9-dehydro-ferruginol (112) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 7.2 | 12.3 | [91] |
| 11,12-dihydroxy-20-norabieta-5(10),8,11,13-tetraen-1-one (113) | Perovskia abrotanoides Kar. | T. b. rhodesiense (tryp.) | 12.0 | 12.5 | [91] |
| bokkosin (114) | Calliandra portoricensis Hassk. | T. b. brucei ((ryp.) | 1.1 | 246 | [92] |
| 8-oxacassa-13,15-dien-7-ol-17-al (115) | Acacia nilotica L. | T. b. brucei (tryp.) | 1.4 | 21.1 | [93] |
| Triterpenes | |||||
| ursolic acid (116) | Vitellaria paradoxa C. F. Gaertn | T. brucei (tryp.) | 2.4 | 4.6 | [94] |
| 3-O-[α-L-rhamnopyranosyl-(1→2)-β-D- xylopyranosyl]pomolic acid (117) | Vangueria agrestis (Schweinf. ex Hiern) Lantz | T. brucei (tryp.) | 11.1 | n.d. | [96] |
| kotschyienone A (122) | Pseudocedrela kotschyi (Schweinf.) Harms | T. brucei (tryp.) | 2.5 | 12.6 | [99] |
| 7-deacetyl-7-oxogedunin (125) | Pseudocedrela kotschyi (Schweinf.) Harms | T. brucei (tryp.) | 3.18 | >31.4 | [99] |
| baurenol acetate (128) | Tabernaemontana longipes Donn.Sm. | T. brucei (tryp.) | 3.1 | >25.8 | [100] |
| baurenol (129) | Tabernaemontana longipes Donn.Sm. | T. brucei (tryp.) | 2.7 | >29.6 | [100] |
| polycarpol (134) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | T. b. brucei (tryp.) | 8.1 | 1.0 | [102] |
| dihydropolycarpol (135) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | T. b. brucei (tryp.) | 8.1 | 2.4 | [102] |
| cyclovirobuxeine-B (136) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.5 | 25.0 | [103] |
| cyclomicrophylline-A (138) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 2.3 | 42.0 | [103] |
| N-benzoyl-O-acetyl-cycloxo-buxoline-F (141) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 2.4 | 30.0 | [103] |
| N20-acetylbuxadine- G (142) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.3 | 33.0 | [103] |
| O-benzoyl-cycloprotobuxoline-D (146) | Buxus sempervirens L. | T. b. rhodesiense (tryp.) | 1.1 | 12.0 | [103] |
a Some names are not indicated in the corresponding papers; b Tryp: trypomastigotes bloodstream forms: n.d.: not defined/not determined.
Table 2.
Plant terpenoids with anti-T. cruzi activity (2016–2021).
| Compound a | Plant | Parasite Form | IC50 (µM) | SI | Refs. |
|---|---|---|---|---|---|
| Monoterpenes and Iridoids | |||||
| N-{1-methyl-1-[(1R)-4-methylcyclohex-3-en-1- yl]ethyl}-5-(4-methylphenyl)-1,3,4-thiadiazol-2-amine (1) | n.d | Amastigote | 1.3 | 611.2 | [53] |
| Sesquiterpenes | |||||
| mikanolide (31) | Mikania variifolia Hieron. and Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 2.41 7.24 15.5 | 31.9 10.6 4.3 | [62] |
| deoxymikanolide (32) | Mikania variifolia Hieron. and Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 0.29 5.43 22.8 | 992.5 54.0 12.5 | [62] |
| dihydromikanolide (33) | Mikania micrantha Kunth | Epimastigote Trypomastigote Amastigote | 8.55 1.03 29.1 | 5.2 49.0 1.5 | [62] |
| nobilinon A (39) | Anthemis nobilis L. | Intracellular amastigote | 2.8 | 0.5 | [64] |
| nobilinon B (40) | Anthemis nobilis L. | Intracellular amastigote | 4.2 | 6.1 | [64] |
| 11,13-dihydroxy-calaxin (41) | Calea pinnatifida (R. Br.) Less. | Amastigote | 8.30 | 1.88 | [65] |
| enhydrin (42) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 0.78 | n.d. | [66] |
| uvedalin (43) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 0.79 | n.d. | [66] |
| polymatin (44) | Smallanthus sonchifolius (Poepp.) H. Rob. | Epimastigote | 1.38 | n.d. | [66] |
| 45 | Scleria striatinux De Wild. | Amastigote | 0.025 | 0.74 | [67] |
| 46 | Scleria striatinux De Wild. | Amastigote | 0.085 | 1.0 | [67] |
| eupatoriopicrin (51) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 19.9 6.3 | 12.9 40.6 | [68] |
| estafietin (52) | Astearaceae species | Trypomastigote bloodstream form | 117 | 6.8 | [68] |
| eupahakonenin B (53) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 33.0 89.3 | 10.4 3.8 | [68] |
| minimolide (54) | Astearaceae species | Trypomastigote bloodstream form Intracellular amastigote | 21.0 25.1 | 12.8 10.7 | [68] |
| tagitinin C (55) | Tithonia diversifolia (Hemsl.) A. Gray | Epimastigote | 1.15 | 5.69 | |
| 4,15-iso-atriplicolide methacrylate (56) | Helianthus tuberosus L. | Trypomastigote | 1.6 | 0.32 | [73] |
| 4,15-iso-atriplicolide isobutryrate (57) | Helianthus tuberosus L. | Trypomastigote | 3.1 | 0.28 | [73] |
| heliantuberolide-8-O-tiglate (58) | Helianthus tuberosus L. | Trypomastigote | 5.7 | 0.68 | [73] |
| (2-methoxy-2,5-epoxy-8-metha- cryloxygermacra-3Z,11(13)-dien-6,12-olide (59) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 1.5 | > 14 | [71] |
| (2-ethoxy-2, 5-epoxy-8-angeloxygermacra-3Z,11(13)-dien-6,12-olide (60) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 2.1 | > 14 | [71] |
| 8a-methacryloxyhirsutinolide 13-O-acetate (61) | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 2.0 | > 14 | [71] |
| 62 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 3.7 | 14.3 | [71] |
| 66 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 10.7 | 9.0 | [71] |
| 67 | Vernonanthura nebularum (Cabrera) H. Rob. | Epimastigote | 8.1 | 13.9 | [71] |
| 68 | Centratherum puctatum ssp. Punctatum Cass. | Epimastigote | 6.8 | 1.6 | [71] |
| 69 | Elephantopus mollis Kunth | Epimastigote | 4.7 | 11.5 | [71] |
| eucannabinolide (70) | Urolepis hecatantha (DC.) R.King & H.Rob. | Epimastigote | 10 | 1.5 | [72] |
| santhemoidin C (71) | Urolepis hecatantha (DC.) R.King & H.Rob. | Epimastigote | 18 | 0.83 | [72] |
| goyazensolide (72) | Lychnophora passerina (Mart ex DC) Gardn. | Intracellular amastigote | 0.181/24 h 0.020/48 h | 52.82/24h 915.0/48h | [73] |
| hedyosulide (73) | Hedyosmum brasiliense Mart. ex Miq. | Trypomastigote Intracellular amastigote | 28.1 21.6 | >7 >9 | [74] |
| 8-epi-xanthatin-1β,5β-epoxide (75) | Inula viscosa (L.) Greuter | Epimastigote | 4.99 | 3.67 | [75] |
| inuloxin A (76) | Inula viscosa (L.) Greuter | Epimastigote | 15.52 | 3.38 | [75] |
| costic acid (77) | Nectandra barbellata Coe-Teix. | Intracellular amastigote | 7.9 | >25 | [76] |
| polygodial (80) | Polygonum hydropiper L. | Epimastigote Trypomastigote Amastigote | 51.0 68.2 34.4 | n.d | [79] |
| Polygodial derivative (81) | n.d | Epimastigote Trypomastigote Amastigote | 13.0 8.4 9.9 | n.d | [79] |
| Polygodial derivative (82) | n.d | Epimastigote Trypomastigote Amastigote | 12.3 6.4 6.7 | n.d | [79] |
| Polygodial derivative (83) | n.d | Epimastigote Trypomastigote Amastigote | 7.2 6.9 8.3 | n.d | [79] |
| epi-polygodial (84) | Drimys brasiliensis Miers | Trypomastigote | 5.01 | >40 | [80] |
| (-)-T-cadinol (85) | Casearia sylvestris Sw. | Trypomastigote Amastigote | 18.2 15.8 | >15 | [81] |
| Diterpenes | |||||
| ent-15β-senecioyl-oxy-kaur-16-en-19-oic acid (89) | Baccharis retusa DC. | Trypomastigote | 3.8 | 50.0 | [84] |
| taxodione (95) | Salvia austriaca Jacq. | Amastigote | 7.11 | 0.27 | [86] |
| 7-(20-oxohexyl)-taxodione (96) | Salvia austriaca Jacq. | Amastigote | 7.76 | 0.4 | [86] |
| Taxodone (97) | Salvia austriaca Jacq. | Amastigote | 7.63 | 0.5 | [86] |
| ent-7-oxo-pimara-8,15-diene-18-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | Amastigote | 15.4 | 3.0 | [87] |
| ent-2S,4S-2-19-epoxy-pimara-8(3),15-diene-7β-ol (98) | Aldama discolors (Baker) E.E.Schill. & Panero | Amastigote | 19.4 | 4.0 | [87] |
| dehydroabietylamine derivative(103) | n.d | Amastigote | 0.6 | 58.0 | [88] |
| Leriifolione (104) | Salvia leriifolia Benth. | Amastigote | 2.6 | 0.6 | [89] |
| Triterpenes | |||||
| Betulinic acid (118) | n.d | trypomastigote | 19.5 | 18.8 | [97] |
| Betulinic acid derivative (119) | n.d | trypomastigote | 1.8 | 17.3 | [97] |
| Betulinic acid derivative (120) | n.d | trypomastigote | 5.0 | 10.7 | [97] |
| Betulinic acid derivative (121) | n.d | trypomastigote | 5.4 | 5.3 | [97] |
| Perovskone C (130) | Salvia hydrangea DC. ex Benth. | Amastigote | 3.5 | 10.7 | [101] |
| perovskone D (131) | Salvia hydrangea DC. ex Benth. | Amastigote | 3.8 | 3.6 | [101] |
| perovskone E (132) | Salvia hydrangea DC. ex Benth. | Amastigote | 11.5 | 6.3 | [101] |
| perovskone F (133) | Salvia hydrangea DC. ex Benth. | Amastigote | 19.8 | 2.4 | [101] |
| polycarpol (134) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | trypomastigote | 1.4 | 2.0 | [102] |
| dihydropolycarpol (135) | Greenwayodendron suaveolens (Engl. & Diels) Verdc. | trypomastigote | 2.4 | 8.1 | [102] |
a Some names are not indicated in the corresponding papers; n.d.: not defined/not determined.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Durão, R.; Ramalhete, C.; Madureira, A.M.; Mendes, E.; Duarte, N. Plant Terpenoids as Hit Compounds against Trypanosomiasis. Pharmaceuticals 2022, 15, 340. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15030340
AMA Style
Durão R, Ramalhete C, Madureira AM, Mendes E, Duarte N. Plant Terpenoids as Hit Compounds against Trypanosomiasis. Pharmaceuticals. 2022; 15(3):340. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15030340
Chicago/Turabian StyleDurão, Raquel, Cátia Ramalhete, Ana Margarida Madureira, Eduarda Mendes, and Noélia Duarte. 2022. "Plant Terpenoids as Hit Compounds against Trypanosomiasis" Pharmaceuticals 15, no. 3: 340. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15030340
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.