
Nutraceutical Interventions for Post-Traumatic Stress Disorder in Animal Models: A Focus on the Hypothalamic–Pituitary–Adrenal Axis

Abstract
:1. Introduction
2. Methods
3. Role of the HPA Axis in Stress Response
4. Role of the HPA Axis in PTSD
4.1. Changes in the HPA Axis in PTSD-Related Brain Regions
4.2. Involvement of the HPA Axis in Inflammation in PTSD
5. Effects of Herbal Medicines on PTSD via Amelioration of Disturbances in the HPA Axis
5.1. Bioactive Compounds
5.2. Herbal Extracts
5.3. Herbal Formula
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- What is Posttraumatic Stress Disorder (PTSD)? Available online: https://www.psychiatry.org/patients-families/ptsd/what-is-ptsd (accessed on 31 August 2020).
- Guillen-Burgos, H.F.; Gutierrez-Ruiz, K. Genetic Advances in Post-traumatic Stress Disorder. Rev. Colomb. Psiquiatr. (Engl. Ed.) 2018, 47, 108–118. [Google Scholar] [CrossRef]
- Fischer, S.; Schumacher, T.; Knaevelsrud, C.; Ehlert, U.; Schumacher, S. Genes and hormones of the hypothalamic-pituitary-adrenal axis in post-traumatic stress disorder. What is their role in symptom expression and treatment response? J. Neural Transm. 2021, 128, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Aliev, G.; Beeraka, N.M.; Nikolenko, V.N.; Svistunov, A.A.; Rozhnova, T.; Kostyuk, S.; Cherkesov, I.; Gavryushova, L.V.; Chekhonatsky, A.A.; Mikhaleva, L.M.; et al. Neurophysiology and Psychopathology Underlying PTSD and Recent Insights into the PTSD Therapies-A Comprehensive Review. J. Clin. Med. 2020, 9, 2951. [Google Scholar] [CrossRef] [PubMed]
- Schottenbauer, M.A.; Glass, C.R.; Arnkoff, D.B.; Tendick, V.; Gray, S.H. Nonresponse and dropout rates in outcome studies on PTSD: Review and methodological considerations. Psychiatry 2008, 71, 134–168. [Google Scholar] [CrossRef] [PubMed]
- Machado-Vieira, R.; Baumann, J.; Wheeler-Castillo, C.; Latov, D.; Henter, I.D.; Salvadore, G.; Zarate, C.A. The Timing of Antidepressant Effects: A Comparison of Diverse Pharmacological and Somatic Treatments. Pharmaceuticals 2010, 3, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Koek, R.J.; Schwartz, H.N.; Scully, S.; Langevin, J.P.; Spangler, S.; Korotinsky, A.; Jou, K.; Leuchter, A. Treatment-refractory posttraumatic stress disorder (TRPTSD): A review and framework for the future. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 70, 170–218. [Google Scholar] [CrossRef] [PubMed]
- Numata, T.; Gunfan, S.; Takayama, S.; Takahashi, S.; Monma, Y.; Kaneko, S.; Kuroda, H.; Tanaka, J.; Kanemura, S.; Nara, M.; et al. Treatment of posttraumatic stress disorder using the traditional Japanese herbal medicine saikokeishikankyoto: A randomized, observer-blinded, controlled trial in survivors of the great East Japan earthquake and tsunami. Evid.-Based Complement. Altern. Med. 2014, 2014, 683293. [Google Scholar] [CrossRef]
- Ma, K.; Wang, X.; Feng, S.; Xia, X.; Zhang, H.; Rahaman, A.; Dong, Z.; Lu, Y.; Li, X.; Zhou, X.; et al. From the perspective of Traditional Chinese Medicine: Treatment of mental disorders in COVID-19 survivors. Biomed. Pharm. 2020, 132, 110810. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, C. Ellen Kamhi, phd, rn: Herbal Support for the HPA Axis. Integr. Med. 2016, 15, 42–46. [Google Scholar]
- Liu, L.; Liu, C.; Wang, Y.; Wang, P.; Li, Y.; Li, B. Herbal Medicine for Anxiety, Depression and Insomnia. Curr. Neuropharmacol. 2015, 13, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, L.; Zhang, Y.Z.; Liu, X.M.; Yu, N.J.; Chen, H.X.; Zhao, N.; Yuan, L.; Li, Y.F. Total flavonoids extracted from xiaobuxin-tang on the hyperactivity of hypothalamic-pituitary-adrenal axis in chronically stressed rats. Evid.-Based Complement. Altern. Med. 2011, 2011, 367619. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gu, J.; Wang, X.; Xie, K.; Luan, Q.; Wan, N.; Zhang, Q.; Jiang, H.; Liu, D. Antidepressant-like activity of resveratrol treatment in the forced swim test and tail suspension test in mice: The HPA axis, BDNF expression and phosphorylation of ERK. Pharm. Biochem. Behav. 2013, 112, 104–110. [Google Scholar] [CrossRef]
- Kitchener, P.; Di Blasi, F.; Borrelli, E.; Piazza, P.V. Differences between brain structures in nuclear translocation and DNA binding of the glucocorticoid receptor during stress and the circadian cycle. Eur. J. Neurosci. 2004, 19, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Coric, V.; Feldman, H.H.; Oren, D.A.; Shekhar, A.; Pultz, J.; Dockens, R.C.; Wu, X.; Gentile, K.A.; Huang, S.P.; Emison, E.; et al. Multicenter, randomized, double-blind, active comparator and placebo-controlled trial of a corticotropin-releasing factor receptor-1 antagonist in generalized anxiety disorder. Depress. Anxiety 2010, 27, 417–425. [Google Scholar] [CrossRef]
- White, M.R.; Graziano, M.J.; Sanderson, T.P. Toxicity of Pexacerfont, a Corticotropin-Releasing Factor Type 1 Receptor Antagonist, in Rats and Dogs. Int. J. Toxicol. 2019, 38, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Arato, M.; Banki, C.M.; Bissette, G.; Nemeroff, C.B. Elevated CSF CRF in suicide victims. Biol. Psychiatry 1989, 25, 355–359. [Google Scholar] [CrossRef]
- Raadsheer, F.C.; van Heerikhuize, J.J.; Lucassen, P.J.; Hoogendijk, W.J.; Tilders, F.J.; Swaab, D.F. Corticotropin-releasing hormone mRNA levels in the paraventricular nucleus of patients with Alzheimer’s disease and depression. Am. J. Psychiatry 1995, 152, 1372–1376. [Google Scholar]
- Michopoulos, V.; Norrholm, S.D.; Jovanovic, T. Diagnostic Biomarkers for Posttraumatic Stress Disorder: Promising Horizons from Translational Neuroscience Research. Biol. Psychiatry 2015, 78, 344–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Elia, A.T.D.; Juruena, M.F.; Coimbra, B.M.; Mello, M.F.; Mello, A.F. Posttraumatic stress disorder (PTSD) and depression severity in sexually assaulted women: Hypothalamic-pituitary-adrenal (HPA) axis alterations. BMC Psychiatry 2021, 21, 174. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Morinobu, S.; Fuchikami, M.; Kurata, A.; Kozuru, T.; Yamawaki, S. Effects of single prolonged stress and D-cycloserine on contextual fear extinction and hippocampal NMDA receptor expression in a rat model of PTSD. Neuropsychopharmacology 2008, 33, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.R.; Noble, L.J.; McIntyre, C.K. Using the Single Prolonged Stress Model to Examine the Pathophysiology of PTSD. Front. Pharm. 2017, 8, 615. [Google Scholar] [CrossRef] [PubMed]
- Deslauriers, J.; Toth, M.; Der-Avakian, A.; Risbrough, V.B. Current Status of Animal Models of Posttraumatic Stress Disorder: Behavioral and Biological Phenotypes, and Future Challenges in Improving Translation. Biol. Psychiatry 2018, 83, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.H. Effects of Epigallocatechin Gallate on Behavioral and Cognitive Impairments, Hypothalamic-Pituitary-Adrenal Axis Dysfunction, and Alternations in Hippocampal BDNF Expression Under Single Prolonged Stress. J. Med. Food 2018, 21, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.H. Tetramethylpyrazine reverses anxiety-like behaviors in a rat model of post-traumatic stress disorder. Korean J. Physiol. Pharm. 2018, 22, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Arborelius, L.; Owens, M.J.; Plotsky, P.M.; Nemeroff, C.B. The role of corticotropin-releasing factor in depression and anxiety disorders. J. Endocrinol. 1999, 160, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Choi, G.M.; Sur, B. Antidepressant-Like Effects of Hesperidin in Animal Model of Post-Traumatic Stress Disorder. Chin. J. Integr. Med. 2021, 27, 39–46. [Google Scholar] [CrossRef]
- Higuchi, Y.; Soga, T.; Parhar, I.S. Regulatory Pathways of Monoamine Oxidase A during Social Stress. Front. Neurosci. 2017, 11, 604. [Google Scholar] [CrossRef] [Green Version]
- Homberg, J.R. Serotonergic modulation of conditioned fear. Scientifica 2012, 2012, 821549. [Google Scholar] [CrossRef] [Green Version]
- Raymundi, A.M.; da Silva, T.R.; Sohn, J.M.B.; Bertoglio, L.J.; Stern, C.A. Effects of (9)-tetrahydrocannabinol on aversive memories and anxiety: A review from human studies. BMC Psychiatry 2020, 20, 420. [Google Scholar] [CrossRef]
- De Quervain, D.; Wolf, O.T.; Roozendaal, B. Glucocorticoid-induced enhancement of extinction-from animal models to clinical trials. Psychopharmacology 2019, 236, 183–199. [Google Scholar] [CrossRef] [Green Version]
- De Quervain, D.; Schwabe, L.; Roozendaal, B. Stress, glucocorticoids and memory: Implications for treating fear-related disorders. Nat. Rev. Neurosci. 2017, 18, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Logue, M.W.; van Rooij, S.J.H.; Dennis, E.L.; Davis, S.L.; Hayes, J.P.; Stevens, J.S.; Densmore, M.; Haswell, C.C.; Ipser, J.; Koch, S.B.J.; et al. Smaller Hippocampal Volume in Posttraumatic Stress Disorder: A Multisite ENIGMA-PGC Study: Subcortical Volumetry Results from Posttraumatic Stress Disorder Consortia. Biol. Psychiatry 2018, 83, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, S.J.; Kennis, M.; Sjouwerman, R.; van den Heuvel, M.P.; Kahn, R.S.; Geuze, E. Smaller hippocampal volume as a vulnerability factor for the persistence of post-traumatic stress disorder. Psychol. Med. 2015, 45, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Morey, R.A.; Gold, A.L.; LaBar, K.S.; Beall, S.K.; Brown, V.M.; Haswell, C.C.; Nasser, J.D.; Wagner, H.R.; McCarthy, G.; for the Mid-Atlantic MIRECC Workgroup. Amygdala volume changes in posttraumatic stress disorder in a large case-controlled veterans group. Arch. Gen. Psychiatry 2012, 69, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Rauch, S.L.; Shin, L.M.; Segal, E.; Pitman, R.K.; Carson, M.A.; McMullin, K.; Whalen, P.J.; Makris, N. Selectively reduced regional cortical volumes in post-traumatic stress disorder. Neuroreport 2003, 14, 913–916. [Google Scholar]
- Im, J.J.; Namgung, E.; Choi, Y.; Kim, J.Y.; Rhie, S.J.; Yoon, S. Molecular Neuroimaging in Posttraumatic Stress Disorder. Exp. Neurobiol. 2016, 25, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Jankord, R.; Herman, J.P. Limbic regulation of hypothalamo-pituitary-adrenocortical function during acute and chronic stress. Ann. N. Y. Acad. Sci. 2008, 1148, 64–73. [Google Scholar] [CrossRef]
- Wang, Z.; Neylan, T.C.; Mueller, S.G.; Lenoci, M.; Truran, D.; Marmar, C.R.; Weiner, M.W.; Schuff, N. Magnetic resonance imaging of hippocampal subfields in posttraumatic stress disorder. Arch. Gen. Psychiatry 2010, 67, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.P.; Hayes, S.; Miller, D.R.; Lafleche, G.; Logue, M.W.; Verfaellie, M. Automated measurement of hippocampal subfields in PTSD: Evidence for smaller dentate gyrus volume. J. Psychiatr. Res. 2017, 95, 247–252. [Google Scholar] [CrossRef]
- Tata, D.A.; Anderson, B.J. The effects of chronic glucocorticoid exposure on dendritic length, synapse numbers and glial volume in animal models: Implications for hippocampal volume reductions in depression. Physiol. Behav. 2010, 99, 186–193. [Google Scholar] [CrossRef]
- Roozendaal, B.; Phillips, R.G.; Power, A.E.; Brooke, S.M.; Sapolsky, R.M.; McGaugh, J.L. Memory retrieval impairment induced by hippocampal CA3 lesions is blocked by adrenocortical suppression. Nat. Neurosci. 2001, 4, 1169–1171. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. Glucocorticoid-sensitive hippocampal neurons are involved in terminating the adrenocortical stress response. Proc. Natl. Acad. Sci. USA 1984, 81, 6174–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; Schafer, M.K.; Young, E.A.; Thompson, R.; Douglass, J.; Akil, H.; Watson, S.J. Evidence for hippocampal regulation of neuroendocrine neurons of the hypothalamo-pituitary-adrenocortical axis. J. Neurosci. 1989, 9, 3072–3082. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M.; Uno, H.; Rebert, C.S.; Finch, C.E. Hippocampal damage associated with prolonged glucocorticoid exposure in primates. J. Neurosci. 1990, 10, 2897–2902. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Jiang, Y.; Wang, J.; Liu, Y.; Xiao, M.; Song, C.; Bai, Y.; Yinuo Han, N.; Han, F. Synapse impairment associated with enhanced apoptosis in post-traumatic stress disorder. Synapse 2020, 74, e22134. [Google Scholar] [CrossRef]
- Chen, H.; Lombes, M.; Le Menuet, D. Glucocorticoid receptor represses brain-derived neurotrophic factor expression in neuron-like cells. Mol. Brain 2017, 10, 12. [Google Scholar] [CrossRef]
- Cheng, W.; Han, F.; Shi, Y. Neonatal isolation modulates glucocorticoid-receptor function and synaptic plasticity of hippocampal and amygdala neurons in a rat model of single prolonged stress. J. Affect. Disord. 2019, 246, 682–694. [Google Scholar] [CrossRef]
- Wang, H.T.; Han, F.; Shi, Y.X. Activity of the 5-HT1A receptor is involved in the alteration of glucocorticoid receptor in hippocampus and corticotropin-releasing factor in hypothalamus in SPS rats. Int. J. Mol. Med. 2009, 24, 227–231. [Google Scholar]
- Zhang, L.; Lu, L.; Bu, X.; Li, H.; Tang, S.; Gao, Y.; Liang, K.; Zhang, S.; Hu, X.; Wang, Y.; et al. Alterations in hippocampal subfield and amygdala subregion volumes in posttraumatic subjects with and without posttraumatic stress disorder. Hum. Brain Mapp. 2021, 42, 2147–2158. [Google Scholar] [CrossRef]
- Roozendaal, B.; McEwen, B.S.; Chattarji, S. Stress, memory and the amygdala. Nat. Rev. Neurosci. 2009, 10, 423–433. [Google Scholar] [CrossRef]
- Neves, L.T.; Neves, P.F.R.; Paz, L.V.; Zancan, M.; Milanesi, B.B.; Lazzari, G.Z.; da Silva, R.B.; de Oliveira, M.; Venturin, G.T.; Greggio, S.; et al. Increases in dendritic spine density in BLA without metabolic changes in a rodent model of PTSD. Brain Struct. Funct. 2019, 224, 2857–2870. [Google Scholar] [CrossRef]
- Zhang, H.H.; Meng, S.Q.; Guo, X.Y.; Zhang, J.L.; Zhang, W.; Chen, Y.Y.; Lu, L.; Yang, J.L.; Xue, Y.X. Traumatic Stress Produces Delayed Alterations of Synaptic Plasticity in Basolateral Amygdala. Front. Psychol. 2019, 10, 2394. [Google Scholar] [CrossRef] [Green Version]
- Roozendaal, B. Stress and memory: Opposing effects of glucocorticoids on memory consolidation and memory retrieval. Neurobiol. Learn. Mem. 2002, 78, 578–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roozendaal, B. 1999 Curt P. Richter award. Glucocorticoids and the regulation of memory consolidation. Psychoneuroendocrinology 2000, 25, 213–238. [Google Scholar] [CrossRef]
- Han, F.; Ding, J.; Shi, Y. Expression of amygdala mineralocorticoid receptor and glucocorticoid receptor in the single-prolonged stress rats. BMC Neurosci. 2014, 15, 77. [Google Scholar] [CrossRef] [Green Version]
- Korte, S.M.; de Boer, S.F.; de Kloet, E.R.; Bohus, B. Anxiolytic-like effects of selective mineralocorticoid and glucocorticoid antagonists on fear-enhanced behavior in the elevated plus-maze. Psychoneuroendocrinology 1995, 20, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Lee, H.J.; Han, J.S.; Packard, M.G. Amygdala is critical for stress-induced modulation of hippocampal long-term potentiation and learning. J. Neurosci. 2001, 21, 5222–5228. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Koo, J.W.; Lee, H.J.; Han, J.S. Amygdalar inactivation blocks stress-induced impairments in hippocampal long-term potentiation and spatial memory. J. Neurosci. 2005, 25, 1532–1539. [Google Scholar] [CrossRef]
- Roozendaal, B.; de Quervain, D.J.; Ferry, B.; Setlow, B.; McGaugh, J.L. Basolateral amygdala-nucleus accumbens interactions in mediating glucocorticoid enhancement of memory consolidation. J. Neurosci. 2001, 21, 2518–2525. [Google Scholar] [CrossRef] [Green Version]
- Mahan, A.L.; Ressler, K.J. Fear conditioning, synaptic plasticity and the amygdala: Implications for posttraumatic stress disorder. Trends Neurosci. 2012, 35, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.A.; LeDoux, J.E. Differential contribution of dorsal and ventral medial prefrontal cortex to the acquisition and extinction of conditioned fear in rats. Behav. Neurosci. 1995, 109, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Robbins, T.W.; Roberts, A.C. Dissociation in prefrontal cortex of affective and attentional shifts. Nature 1996, 380, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Quirk, G.J.; Likhtik, E.; Pelletier, J.G.; Pare, D. Stimulation of medial prefrontal cortex decreases the responsiveness of central amygdala output neurons. J. Neurosci. 2003, 23, 8800–8807. [Google Scholar] [CrossRef]
- Newport, D.J.; Nemeroff, C.B. Neurobiology of posttraumatic stress disorder. Curr. Opin. Neurobiol. 2000, 10, 211–218. [Google Scholar] [CrossRef]
- McDonald, A.J.; Mascagni, F.; Guo, L. Projections of the medial and lateral prefrontal cortices to the amygdala: A Phaseolus vulgaris leucoagglutinin study in the rat. Neuroscience 1996, 71, 55–75. [Google Scholar] [CrossRef]
- Licznerski, P.; Duric, V.; Banasr, M.; Alavian, K.N.; Ota, K.T.; Kang, H.J.; Jonas, E.A.; Ursano, R.; Krystal, J.H.; Duman, R.S.; et al. Decreased SGK1 Expression and Function Contributes to Behavioral Deficits Induced by Traumatic Stress. PLoS Biol. 2015, 13, e1002282. [Google Scholar] [CrossRef] [Green Version]
- Diorio, D.; Viau, V.; Meaney, M.J. The role of the medial prefrontal cortex (cingulate gyrus) in the regulation of hypothalamic-pituitary-adrenal responses to stress. J. Neurosci. 1993, 13, 3839–3847. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Han, F.; Shi, Y. Changes in the glucocorticoid receptor and Ca(2)(+)/calreticulin-dependent signalling pathway in the medial prefrontal cortex of rats with post-traumatic stress disorder. J. Mol. Neurosci. 2015, 56, 24–34. [Google Scholar] [CrossRef]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Chrousos, G.P. The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N. Engl. J. Med. 1995, 332, 1351–1362. [Google Scholar] [CrossRef]
- Tian, R.; Hou, G.; Li, D.; Yuan, T.F. A possible change process of inflammatory cytokines in the prolonged chronic stress and its ultimate implications for health. Sci. World J. 2014, 2014, 780616. [Google Scholar] [CrossRef] [PubMed]
- Elenkov, I.J.; Webster, E.L.; Torpy, D.J.; Chrousos, G.P. Stress, corticotropin-releasing hormone, glucocorticoids, and the immune/inflammatory response: Acute and chronic effects. Ann. N. Y. Acad. Sci. 1999, 876, 1–13. [Google Scholar] [CrossRef]
- Vashist, S.K.; Schneider, E.M. Depression: An Insight and Need for Personalized Psychological Stress Monitoring and Management. J. Basic Appl. Sci. 2014, 10, 177–182. [Google Scholar] [CrossRef]
- De Pablos, R.M.; Villaran, R.F.; Arguelles, S.; Herrera, A.J.; Venero, J.L.; Ayala, A.; Cano, J.; Machado, A. Stress increases vulnerability to inflammation in the rat prefrontal cortex. J. Neurosci. 2006, 26, 5709–5719. [Google Scholar] [CrossRef] [PubMed]
- McCanlies, E.C.; Araia, S.K.; Joseph, P.N.; Mnatsakanova, A.; Andrew, M.E.; Burchfiel, C.M.; Violanti, J.M. C-reactive protein, interleukin-6, and posttraumatic stress disorder symptomology in urban police officers. Cytokine 2011, 55, 74–78. [Google Scholar] [CrossRef]
- Michopoulos, V.; Powers, A.; Gillespie, C.F.; Ressler, K.J.; Jovanovic, T. Inflammation in Fear- and Anxiety-Based Disorders: PTSD, GAD, and Beyond. Neuropsychopharmacology 2017, 42, 254–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtado, M.; Katzman, M.A. Neuroinflammatory pathways in anxiety, posttraumatic stress, and obsessive compulsive disorders. Psychiatry Res. 2015, 229, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xie, J.; Sun, Y. TLR4/MyD88/NF-kappaB-Mediated Inflammation Contributes to Cardiac Dysfunction in Rats of PTSD. Cell. Mol. Neurobiol. 2020, 40, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Averill, L.A.; Purohit, P.; Averill, C.L.; Boesl, M.A.; Krystal, J.H.; Abdallah, C.G. Glutamate dysregulation and glutamatergic therapeutics for PTSD: Evidence from human studies. Neurosci. Lett. 2017, 649, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Xiao, B.; Wen, L. Loss of Glial Cells of the Hippocampus in a Rat Model of Post-traumatic Stress Disorder. Neurochem. Res. 2015, 40, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Knox, D.; Perrine, S.A.; George, S.A.; Galloway, M.P.; Liberzon, I. Single prolonged stress decreases glutamate, glutamine, and creatine concentrations in the rat medial prefrontal cortex. Neurosci. Lett. 2010, 480, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledgerwood, L.; Richardson, R.; Cranney, J. D-cycloserine and the facilitation of extinction of conditioned fear: Consequences for reinstatement. Behav. Neurosci. 2004, 118, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.L.; Ressler, K.J.; Lu, K.T.; Davis, M. Facilitation of conditioned fear extinction by systemic administration or intra-amygdala infusions of D-cycloserine as assessed with fear-potentiated startle in rats. J. Neurosci. 2002, 22, 2343–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heresco-Levy, U.; Vass, A.; Bloch, B.; Wolosker, H.; Dumin, E.; Balan, L.; Deutsch, L.; Kremer, I. Pilot controlled trial of D-serine for the treatment of post-traumatic stress disorder. Int. J. Neuropsychopharmacol. 2009, 12, 1275–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Pothula, S.; Wu, M.; Kang, H.; Girgenti, M.J.; Picciotto, M.R.; DiLeone, R.J.; Taylor, J.R.; Duman, R.S. Positive modulation of N-methyl-D-aspartate receptors in the mPFC reduces the spontaneous recovery of fear. Mol. Psychiatry 2022, 27, 2580–2589. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.H.; Madhana, R.M.; Athira, K.V.; Kasala, E.R.; Bodduluru, L.N.; Pitta, S.; Mahareddy, J.R.; Lahkar, M. Resveratrol ameliorates depressive-like behavior in repeated corticosterone-induced depression in mice. Steroids 2015, 101, 37–42. [Google Scholar] [CrossRef]
- Magaji, M.G.; Iniaghe, L.O.; Abolarin, M.; Abdullahi, O.I.; Magaji, R.A. Neurobehavioural evaluation of resveratrol in murine models of anxiety and schizophrenia. Metab. Brain Dis. 2017, 32, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Qiu, Z.K.; He, J.L.; Liu, X.; Chen, J.S.; Wang, Y.L. Resveratrol ameliorated the behavioral deficits in a mouse model of post-traumatic stress disorder. Pharm. Biochem. Behav. 2017, 161, 68–76. [Google Scholar] [CrossRef]
- Zhao, Z.; Kim, Y.W.; Wu, Y.; Zhang, J.; Lee, J.H.; Li, X.; Cho, I.J.; Park, S.M.; Jung, D.H.; Yang, C.H.; et al. Korean Red Ginseng attenuates anxiety-like behavior during ethanol withdrawal in rats. J. Ginseng Res. 2014, 38, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Cha, H.Y.; Park, J.H.; Hong, J.T.; Yoo, H.S.; Song, S.; Hwang, B.Y.; Eun, J.S.; Oh, K.W. Anxiolytic-like effects of ginsenosides on the elevated plus-maze model in mice. Biol. Pharm. Bull. 2005, 28, 1621–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.H. Effect of ginsenoside Re on depression- and anxiety-like behaviors and cognition memory deficit induced by repeated immobilization in rats. J. Microbiol. Biotechnol. 2012, 22, 708–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Sur, B.; Cho, S.G.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Ginsenoside Rb1 rescues anxiety-like responses in a rat model of post-traumatic stress disorder. J. Nat. Med. 2016, 70, 133–144. [Google Scholar] [CrossRef]
- Lu, D.; Zhu, L.H.; Shu, X.M.; Zhang, C.J.; Zhao, J.Y.; Qi, R.B.; Wang, H.D.; Lu, D.X. Ginsenoside Rg1 relieves tert-Butyl hydroperoxide-induced cell impairment in mouse microglial BV2 cells. J. Asian Nat. Prod. Res. 2015, 17, 930–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, K.; Chen, L.; Ou Yang, L.; Huang, Y.; Zhao, Y. Preventive effects of ginsenoside Rg1 on post-traumatic stress disorder (PTSD)-like behavior in male C57/B6 mice. Neurosci. Lett. 2015, 605, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Song, Z.; Shen, F.; Xie, P.; Wang, J.; Zhu, A.S.; Zhu, G. Ginsenoside Rg1 Prevents PTSD-Like Behaviors in Mice Through Promoting Synaptic Proteins, Reducing Kir4.1 and TNF-alpha in the Hippocampus. Mol. Neurobiol. 2021, 58, 1550–1563. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Gastrodin reversed the traumatic stress-induced depressed-like symptoms in rats. J. Nat. Med. 2016, 70, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Donato, F.; de Gomes, M.G.; Goes, A.T.; Filho, C.B.; Del Fabbro, L.; Antunes, M.S.; Souza, L.C.; Boeira, S.P.; Jesse, C.R. Hesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: Possible role of l-arginine-NO-cGMP pathway and BDNF levels. Brain Res. Bull. 2014, 104, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Choi, G.M.; Shim, I.; Lee, H. Genistein Prevents Single Prolonged Stress-Induced Cognitive Impairment in a Post-Traumatic Stress Disorder Rat Model via Activation of the Serotonergic System. J. Med. Food 2020, 23, 476–484. [Google Scholar] [CrossRef]
- Dekanski, D.; Selakovic, V.; Piperski, V.; Radulovic, Z.; Korenic, A.; Radenovic, L. Protective effect of olive leaf extract on hippocampal injury induced by transient global cerebral ischemia and reperfusion in Mongolian gerbils. Phytomedicine 2011, 18, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Pourkhodadad, S.; Alirezaei, M.; Moghaddasi, M.; Ahmadvand, H.; Karami, M.; Delfan, B.; Khanipour, Z. Neuroprotective effects of oleuropein against cognitive dysfunction induced by colchicine in hippocampal CA1 area in rats. J. Physiol. Sci. 2016, 66, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.H. Effect of oleuropein on cognitive deficits and changes in hippocampal brain-derived neurotrophic factor and cytokine expression in a rat model of post-traumatic stress disorder. J. Nat. Med. 2018, 72, 44–56. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, W.; Tang, Y.; Bai, W.; Yang, F.; Xie, L.; Li, X.; Zhou, S.; Pan, S.; Chen, Q.; et al. l-Tetrahydropalmatine, an active component of Corydalis yanhusuo W.T. Wang, protects against myocardial ischaemia-reperfusion injury in rats. PLoS ONE 2012, 7, e38627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Sur, B.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. L-tetrahydropalmatine ameliorates development of anxiety and depression-related symptoms induced by single prolonged stress in rats. Biomol. Ther. 2014, 22, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Chun, H.J.; Lee, K.M.; Jung, Y.S.; Lee, J. Silibinin suppresses astroglial activation in a mouse model of acute Parkinson’s disease by modulating the ERK and JNK signaling pathways. Brain Res. 2015, 1627, 233–242. [Google Scholar] [CrossRef]
- Lee, B.; Choi, G.M.; Sur, B. Silibinin prevents depression-like behaviors in a single prolonged stress rat model: The possible role of serotonin. BMC Complement. Med. 2020, 20, 70. [Google Scholar] [CrossRef] [Green Version]
- Lopresti, A.L. Curcumin for neuropsychiatric disorders: A review of in vitro, animal and human studies. J. Psychopharmacol. 2017, 31, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Lee, H. Systemic Administration of Curcumin Affect Anxiety-Related Behaviors in a Rat Model of Posttraumatic Stress Disorder via Activation of Serotonergic Systems. Evid.-Based Complement. Altern. Med. 2018, 2018, 9041309. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, X.; Xu, W.; Farzaneh, F.; Xu, R. The structure and pharmacological functions of coumarins and their derivatives. Curr. Med. Chem. 2009, 16, 4236–4260. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Umbelliferone modulates depression-like symptoms by altering monoamines in a rat post-traumatic stress disorder model. J. Nat. Med. 2020, 74, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Zhu, S.; Ma, J.; Zhu, L.; Liu, Y.; Ou, G.; Li, R.; Wang, Y.; Liang, Y.; Jin, X.; et al. Intranasal temperature-sensitive hydrogels of cannabidiol inclusion complex for the treatment of post-traumatic stress disorder. Acta Pharm. Sin. B 2021, 11, 2031–2047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Yuan, L.; Yang, M.; Luo, Z.P.; Zhao, Y.M. The effect of Morinda officinalis How, a Chinese traditional medicinal plant, on the DRL 72-s schedule in rats and the forced swimming test in mice. Pharm. Biochem. Behav. 2002, 72, 39–43. [Google Scholar] [CrossRef]
- Qiu, Z.K.; Liu, C.H.; Gao, Z.W.; He, J.L.; Liu, X.; Wei, Q.L.; Chen, J.S. The inulin-type oligosaccharides extract from morinda officinalis, a traditional Chinese herb, ameliorated behavioral deficits in an animal model of post-traumatic stress disorder. Metab. Brain Dis. 2016, 31, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Bae, B.S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Sur, B.; Lee, H.; Oh, S. Korean Red Ginseng prevents posttraumatic stress disorder-triggered depression-like behaviors in rats via activation of the serotonergic system. J. Ginseng Res. 2020, 44, 644–654. [Google Scholar] [CrossRef] [PubMed]
- May, B.H.; Lu, C.; Lu, Y.; Zhang, A.L.; Xue, C.C. Chinese herbs for memory disorders: A review and systematic analysis of classical herbal literature. J. Acupunct. Meridian Stud. 2013, 6, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.L.; Ito, H.; Masuoka, T.; Kamei, C.; Hatano, T. Effect of Polygala tenuifolia root extract on scopolamine-induced impairment of rat spatial cognition in an eight-arm radial maze task. Biol. Pharm. Bull. 2007, 30, 1727–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, I.J.; Son, S.U.; Park, H.; Kim, Y.; Park, S.H.; Swanberg, K.; Shin, J.Y.; Ha, S.K.; Cho, Y.; Bang, S.Y.; et al. Preclinical evidence of rapid-onset antidepressant-like effect in Radix Polygalae extract. PLoS ONE 2014, 9, e88617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneikert, J.; Hubner, S.; Martin, E.; Cato, A.C. A nuclear action of the eukaryotic cochaperone RAP46 in downregulation of glucocorticoid receptor activity. J. Cell Biol. 1999, 146, 929–940. [Google Scholar] [CrossRef]
- Shin, J.Y.; Shin, J.W.; Ha, S.K.; Kim, Y.; Swanberg, K.M.; Lee, S.; Kim, T.W.; Maeng, S. Radix Polygalae Extract Attenuates PTSD-like Symptoms in a Mouse Model of Single Prolonged Stress and Conditioned Fear Possibly by Reversing BAG1. Exp. Neurobiol. 2018, 27, 200–209. [Google Scholar] [CrossRef]
- Lee, B.; Hong, R.; Lim, P.; Cho, D.; Yeom, M.; Lee, S.; Kang, K.S.; Lee, S.C.; Shim, I.; Lee, H.; et al. The ethanolic extract of Aralia continentalis ameliorates cognitive deficits via modifications of BDNF expression and anti-inflammatory effects in a rat model of post-traumatic stress disorder. BMC Complement. Altern. Med. 2019, 19, 11. [Google Scholar] [CrossRef]
- Qiu, Z.K.; Zhang, G.H.; He, J.L.; Ma, J.C.; Zeng, J.; Shen, D.; Shen, Y.G.; Chen, J.S.; Liu, C.Y. Free and Easy Wanderer Plus (FEWP) improves behavioral deficits in an animal model of post-traumatic stress disorder by stimulating allopregnanolone biosynthesis. Neurosci. Lett. 2015, 602, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Qu, Y.; Wang, J.; Gao, F.; Ji, M.; Xie, P.; Zhu, A.; Tan, B.; Wang, X.; Zhu, G. Anshen Dingzhi prescription in the treatment of PTSD in mice: Investigation of the underlying mechanism from the perspective of hippocampal synaptic function. Phytomedicine 2022, 101, 154139. [Google Scholar] [CrossRef] [PubMed]


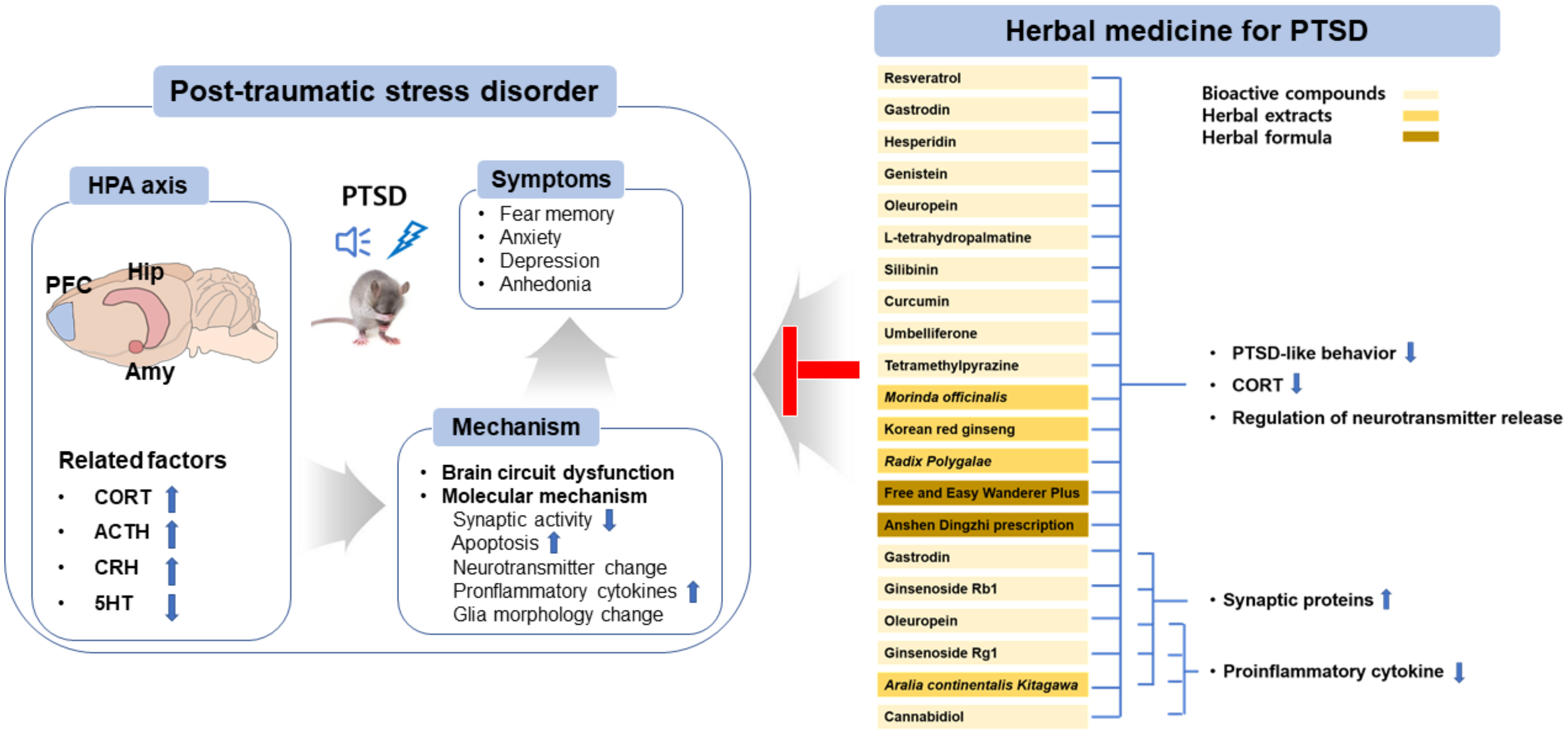{kind=link}
| Herbal Medicine | Model | Treatment (Concentration/ Period) | Action Mechanism | Author (Year) Reference | |
|---|---|---|---|---|---|
| Resveratrol | FS | 20, 40 mg/kg (i.g.) for 17 days | ∙ Anti-anxiety and anti-fear memory behavior ∙ Decreased HPA- axis hormones (CORT, CFH, ACTH) ∙ Increased progesterone and allopregnanolone | Zhang (2017) [89] | |
| Ginsenoside Rb1 (GRb1) | SPS | 30 mg/kg (i.p.) for 14 days | ∙ Anti-anxiety behavior ∙ Increased NPY, decreased TH, increased BDNF mRNA ∙ Decreased CORT | Lee (2015) [93] | |
| Ginsenoside Rg1 (Rg1) | FS | 5 mg/kg (p.o.) for 7 days | ∙ Anti-anxiety behavior ∙ Decreased CORT and CRH | Wang (2015) [95] | |
| Ginsenoside Rg1 (Rg1) | SPS | 20 mg/kg (i.p.) for 14 days | ∙ Anti-depression behavior, increased fear extinction ∙ Decreased hip pro-inflammatory cytokines (IL-1β and TNF-α) ∙ Increased synaptic proteins (PSD95, Arc, GluN2A, GluA1) in hipo | Zhang (2021) [96] | |
| Gastrodin (GAS) | SPS | 100 mg/kg (i.p.) for 14 days | ∙ Anti-depression behavior ∙ Increased body weight ∙ Decreased CORT, NE, and TH ∙ Increase NPY and BDNF | Lee (2016) [97] | |
| Bioactive compounds | Hesperidin (HSD) | SPS | 100 mg/kg (i.p.) for 14 days | ∙Anti-depression, anti-anxiety, and anti-fear memory ∙ Increased sucrose intake ∙ Decreased CORT ∙ Increased 5-HT in the hip and amy ∙ Decreased 5-HIAA/5-HT and MAO-A activity ∙ Increased DA in the hip ∙ Increased TPH1 in the hip | Lee (2021) [27] |
| Genistein (GEN) | SPS | 10 mg/kg (i.p.) for 14 days | ∙ Increased cognitive memory ∙ Decreased CORT ∙ Increased 5-HT level in the hip and mPFC ∙ Decreased 5-HIAA/5-HT ratio and MAO-B activity | Lee. (2020) [99] | |
| Oleuropein (OLE) | SPS | 10 mg/kg (i.p.) for 14 days | ∙ Improved cognitive memory ∙ Decreased CORT ∙ Increased BDNF and CREB in the hip ∙ Decreased TNF-α and Il-1β | Lee. (2018) [102] | |
| L-tetrahydropalmatine (THP) | SPS | 50 mg/kg (i.p.) for 14 days | ∙ Anti-anxiety and anti-depression-like behaviors ∙ Decreased CORT ∙ Increased NYP and decreased CRH in the hypo | Lee. (2020) [104] | |
| Silibinin (SIL) | SPS | 100 mg/kg (i.p.) for 14 days | ∙ Anti-anxiety, anti-depression-like behavior, and anti-fear memory ∙ Decreased CORT ∙ Increased 5-HT in the hip and amy ∙ Decreased 5-HIAA ∙ Increased DA in the hip ∙ Increased TPH-1 mRNA in the hip | Lee. (2020) [106] | |
| Curcumin (CUR), | SPS | 100 mg/kg (i.p.) for 14 days | ∙Anti-anxiety, anti-depression-like behaviors, and anti-fear memory ∙ Decreased CORT ∙ Increased 5-HT in the hip and amy ∙ Increased TPH-1 mRNA in the hip | Lee, (2018) [108] | |
| Umbelliferone (UMB) | SPS | 60 mg/kg (i.p.) for 14 days | ∙ Anti-anxiety, anti-depression-like behaviors, and anti-fear memory ∙ Decreased CORT ∙ Increased 5-HT in the hip and amy ∙ Decreased 5-HIAA/5-HT ratio and MAO-A ∙ Increased DA in the hip | Lee, (2019) [110] | |
| Bioactive compounds | Cannabidiol (CBD) | FS | 30mg/kg (nasal administration) for 10 days | ∙ Anti-anxiety and anti-fear memory ∙ Neuroprotection effects ∙ Decreased c-fos activation in the PFC, hip CA1 and amy ∙ Decrease TNF-α ∙ Decrease 5-HT1A receptor in the hip | Pang (2021) [111] |
| Tetramethylpyrazine (TMP) | SPS | 40 mg/kg (i.p.) for 14 days | ∙ Anti-anxiety and anti-fear memory ∙ Decreased CORT and ACTH ∙ Increased 5-HT in the mPFC and hip ∙ Increased TRP and 5-HIAA mRNA in the hip | Lee, (2018) [25] | |
| Morinda officinalis (MO) | SPS | 25, 50 mg/kg (i.p.) for 13 days | ∙ Anti-anxiety and anti-fear memory ∙ Increased allopregnanolone biosynthesis in the hip, PFC, and amy | Qiu (2016) [113] | |
| Herbal extracts | Korean red ginseng (KRG) | SPS | 100 mg/kg (i.p.) for 14 days | ∙ Anti-depression and anti-fear memory ∙ Decreased CORT ∙ Increased sucrose performance ∙Increased 5-HT in the hip and mPFC, and decreased 5-HIAA/5-HT ∙ Decreased NE and increased DA in the hip ∙ Decreased TPH-1, TPH-2, and MAO-B the hip | Lee (2020) [115] |
| Radix polygalae (RP) | CF + SPS | 0.1 mg/kg (p.o.) for 13 days | ∙ Anti-fear memory ∙ Increased BAG1 in the hip | Shin (2018) [120] | |
| Aralia continentalis Kitagawa (AC) | SPS | 100 mg/kg (i.p.) for 21 days | ∙ Increase recognition memory ∙ Decreased CORT ∙ Increased BDNF and CREB in the hip ∙ Decreased TNF-α and IL-6 in the hip | Lee, (2019) [121] | |
| Herbal formula | Free and Easy Wanderer Plus (FEWP) | SPS | 5, 10 mg/kg (drinking water) for 14 days | ∙ Anti-anxiety and anti-fear memory ∙ Increased allopregnanolone in the PFC, hip, and amy | Qiu (2015) [122] |
| Anshen Dingzhi prescription (ADP) | SPS | 18.4, 36.8 mg/kg (i.g.) for 14 days | ∙ Anti-anxiety and anti-fear memory ∙ Neuroprotective effect ∙Regulated synaptic proteins (PSD95, BDNF-TrkB signaling pathway, μ-calpain/PTEN-AKT-mTOR signaling pathway) | Yang (2022) [123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, M.; Park, H.R.; Yang, E.J. Nutraceutical Interventions for Post-Traumatic Stress Disorder in Animal Models: A Focus on the Hypothalamic–Pituitary–Adrenal Axis. Pharmaceuticals 2022, 15, 898. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15070898
Cai M, Park HR, Yang EJ. Nutraceutical Interventions for Post-Traumatic Stress Disorder in Animal Models: A Focus on the Hypothalamic–Pituitary–Adrenal Axis. Pharmaceuticals. 2022; 15(7):898. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15070898
Chicago/Turabian StyleCai, Mudan, Hee Ra Park, and Eun Jin Yang. 2022. "Nutraceutical Interventions for Post-Traumatic Stress Disorder in Animal Models: A Focus on the Hypothalamic–Pituitary–Adrenal Axis" Pharmaceuticals 15, no. 7: 898. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15070898