
Anti-Inflammatory Effect of Columbianadin against D-Galactose-Induced Liver Injury In Vivo via the JAK2/STAT3 and JAK2/p38/NF-κB Pathways

Abstract
:1. Introduction
2. Results
2.1. Influence on Body Weight and Liver Indices
2.2. Influence on Serum Biochemicals and Inflammatory Cytokines
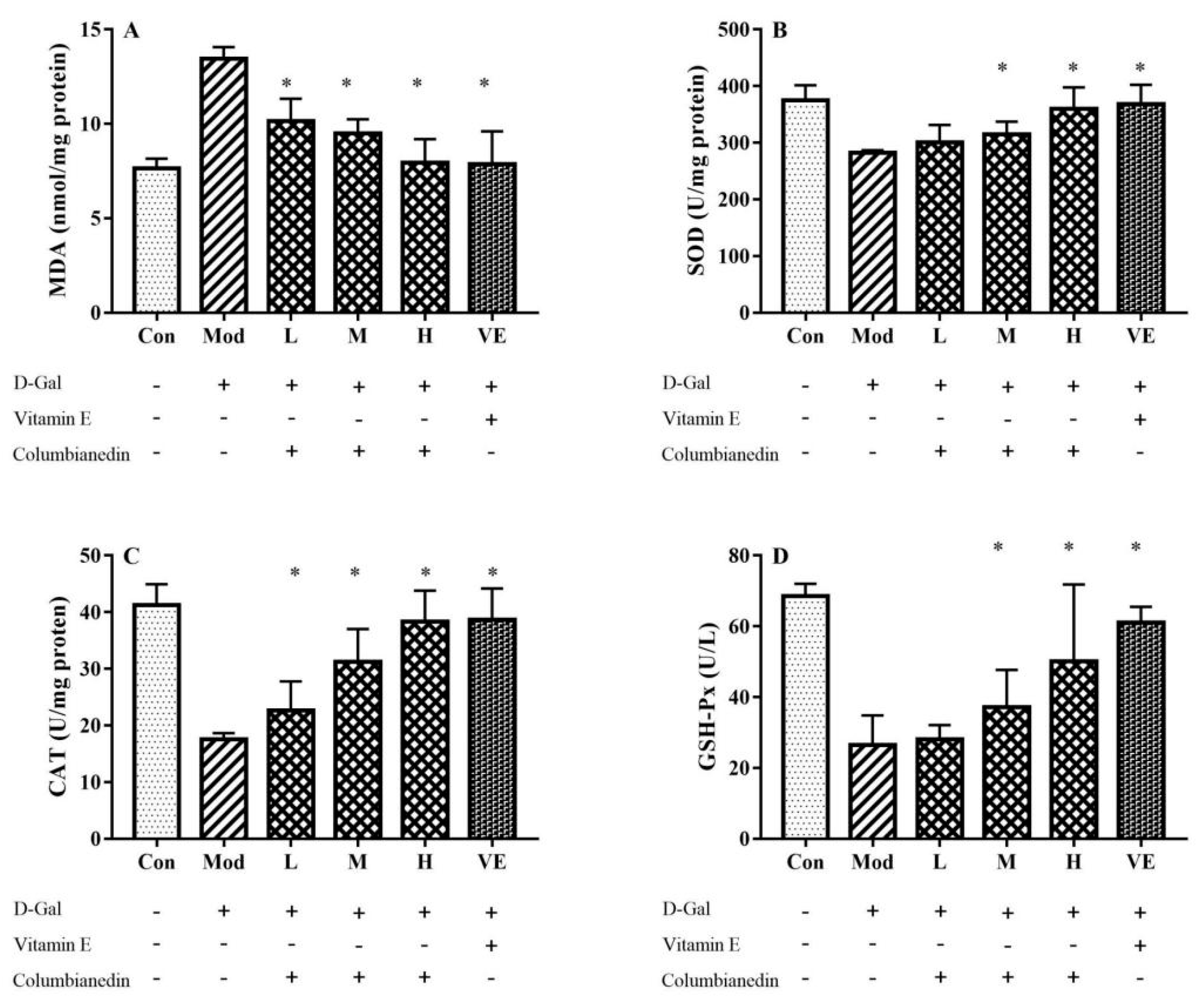
2.3. Influence on D-Gal-Induced Oxidative Stress in Liver Tissue
2.4. Influence on Inflammatory Cytokines in Liver Tissue
2.5. Influence on Liver Histology
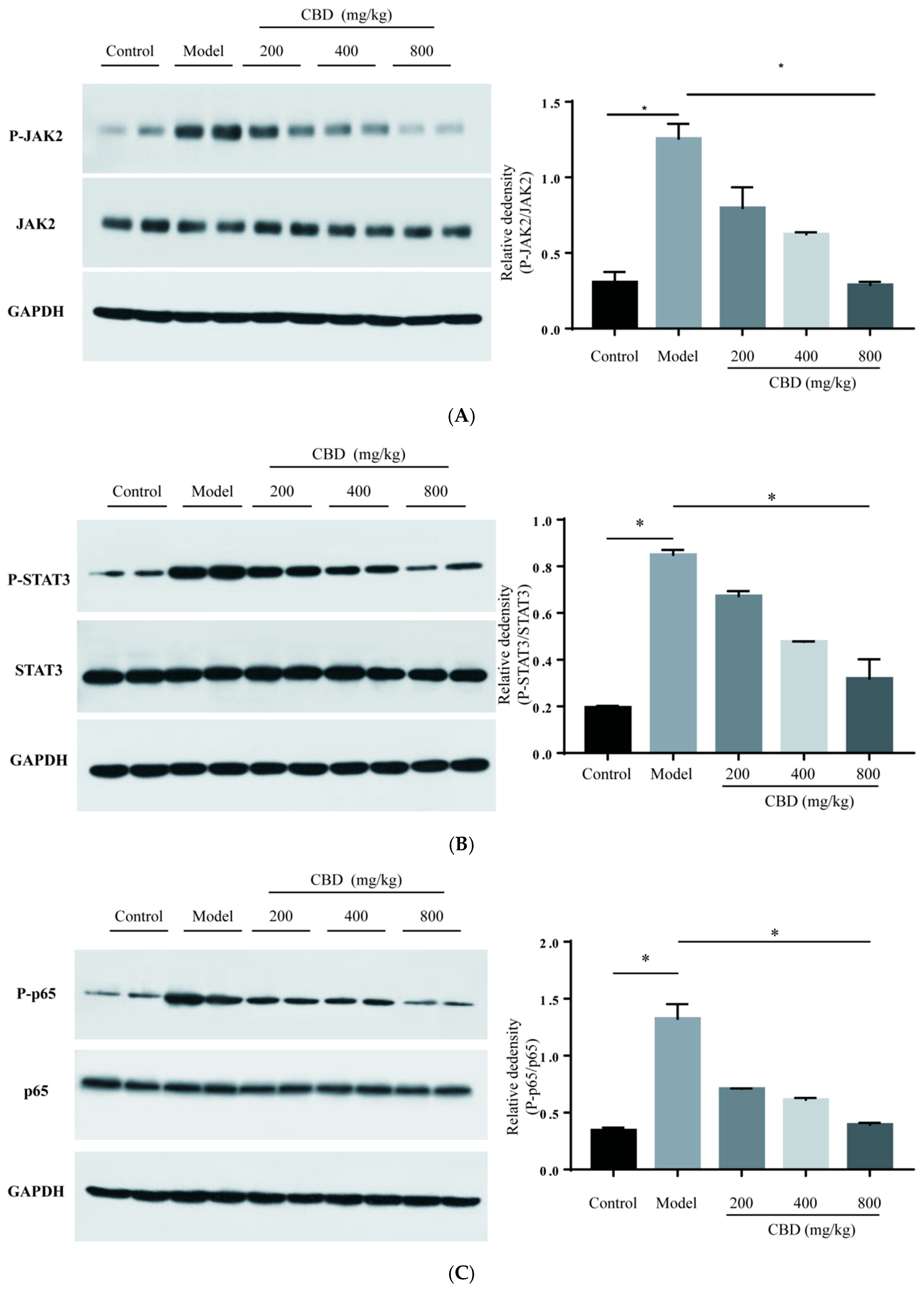
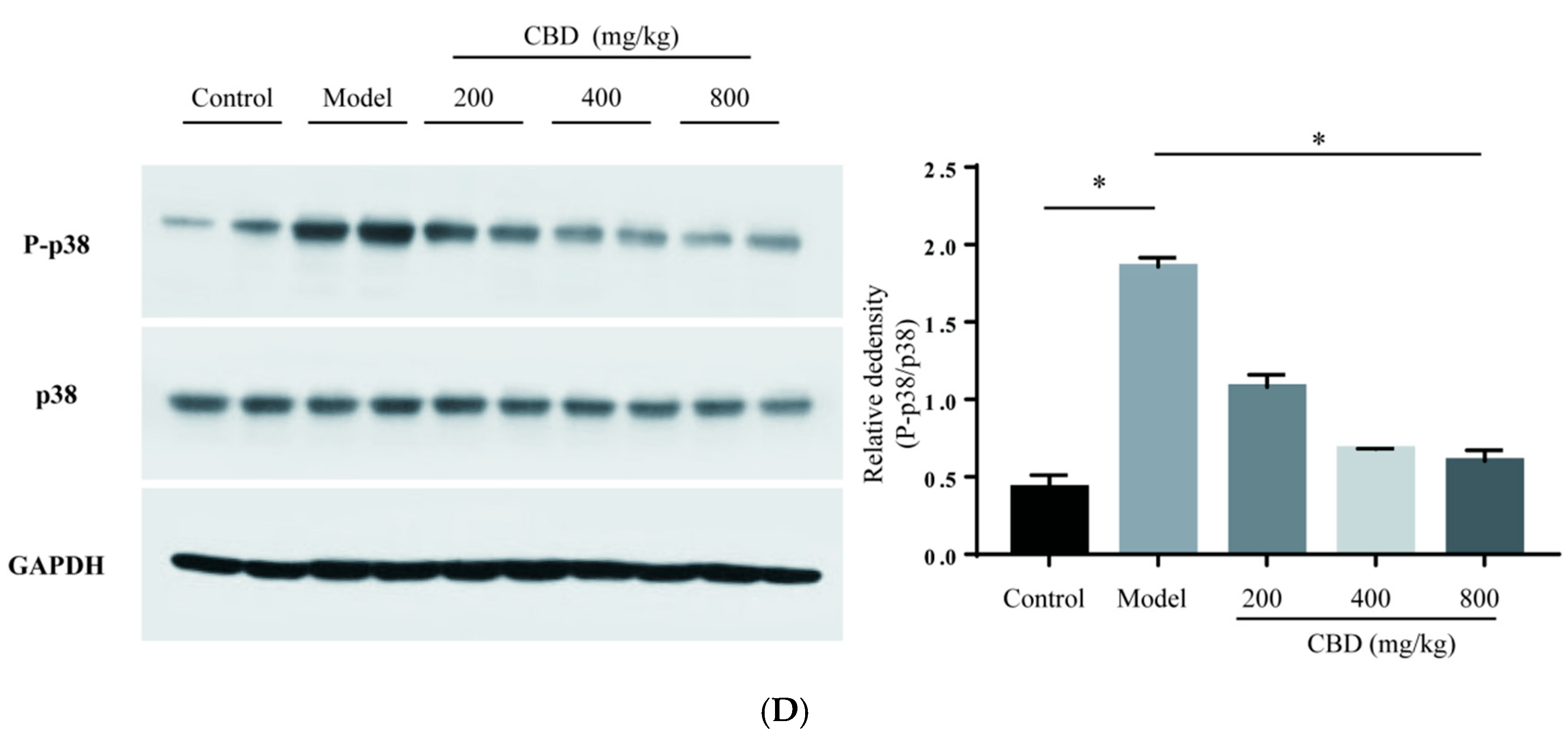
2.6. Influence on the Expression of Proteins in JAK2/STAT3 and JAK2/p38/NF-κB Signaling Pathways
3. Discussion
4. Methods and Materials
4.1. Reagents
4.2. Animal Experiment
4.3. Determination of Body Weight and Organ Indices
4.4. Determination of Biochemical Indices in Serum and Liver
4.5. Tissue Preparation and Histological Analysis
4.6. Western Blot Analysis
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALT | Alanine aminotransferase | JNK | c-Jun N-terminal kinase |
| APR | Angelicae pubescentis radix | MAPK | Mitogen-activated protein kinase |
| AST | Aspartate aminotransferase | NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| BW | Body weight | OE | Osthole |
| CAT | Catalase | p38 | Protein 38 |
| CBA | Columbianetin acetate | p65 | Protein 65 |
| CBN | Columbianadin | P-p38 | Phosphorylated protein 38 |
| CBT | Columbianetin | P-p65 | Phosphorylated protein 65 |
| D-Gal | D-galactose | P-JAK | Phosphorylated Janus kinase |
| ELISA | Enzyme-linked immunosorbent assay | P-STAT | Phosphorylated signal transducer and activator of transcription |
| Erk | Extracellular signal-regulated protein kinase | PVDF | Polyvinylidene difluoride |
| GSH | Glutathione | ROS | Reactive oxygen species |
| GPx | Glutathione peroxidase | SDS-PAGE | Sulfate–polyacrylamide gel electrophoresis |
| H&E | Hematoxylin and eosin | SOD | Superoxide dismutase |
| ICR | Institute of Cancer Research | STAT | Signal transducer and activator of transcription |
| IL-1 | Interleukin-1 | TLR | Toll-like receptor |
| IL-6 | Interleukin-6 | TNF-α | Tumor necrosis factor-α |
| JAK | Janus kinase | VE | Vitamin E |
References
- Ge, Y.Y.; Li, Z.; Zhang, L.; Li, J.; He, J.; Hao, J.; Gao, X.M.; Chang, Y.X. Pharmacokinetics and tissue distribution study of bisabolangelone from Angelicae pubescentis Radix in rat using LC-MS/MS. Biomed. Chromatogr. 2019, 33, e4433. [Google Scholar] [CrossRef]
- El-Gaphar, O.; Abo-Youssef, A.M.; Halal, G.K. Levetiracetam mitigates lipopolysaccharide-induced JAK2/STAT3 and TLR4/MAPK signaling pathways activation in a rat model of adjuvant-induced arthritis. Eur. J. Pharmacol. 2018, 826, 85–95. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Zhang, B.G.; Yang, X.W. Studies on the chemical constituents of the root of Angelica pubescens f. biserrata. Pharm. J. Chin. Peoples Lib. Army 2008, 6, 392–398. [Google Scholar]
- Zhang, Y.; Sun, J.; Tu, P.F.; Xue, P.F. Chemical constituents of Angelica dahurica. J. Inn. Mong. Med. Coll. 2012, 034, 277–280. [Google Scholar]
- Tornquist, K.; Vuorela, H. The furanocoumarin columbianadin inhibits depolarization induced Ca2+ uptake in rat pituitary GH3 cells. Planta Med. 1990, 56, 127–129. [Google Scholar] [CrossRef]
- Li, R.Z.; He, Y.Q.; Chiao, M.; Xu, Y.; Ge, L.P. Studies of the active constituents of the Chinese drug “Duhuo” Angelica pubescens. Acta Pharm. Sin. 1989, 24, 546–551. [Google Scholar]
- Yang, X.-w.; Xu, B.; Ran, F.-x.; Wang, R.-q.; Wu, J.; Cui, J.-r. Inhibitory effects of 11 coumarin compounds against growth of human bladder carcinoma cell line E-J in vitro. Zhong Xi Yi Jie He Xue Bao = J. Chin. Integr. Med. 2007, 5, 56–60. [Google Scholar] [CrossRef]
- Kang, K.-H.; Kong, C.-S.; Seo, Y.; Kim, M.-M.; Kim, S.-K. Anti-inflammatory effect of coumarins isolated from Corydalis heterocarpa in HT-29 human colon carcinoma cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2009, 47, 2129–2134. [Google Scholar] [CrossRef]
- Lim, H.J.; Lee, J.H.; Choi, J.S.; Lee, S.K.; Kim, Y.S.; Kim, H.P. Inhibition of airway inflammation by the roots of Angelica decursiva and its constituent, columbianadin. J. Ethnopharmacol. 2014, 155, 1353–1361. [Google Scholar] [CrossRef]
- Li, R.; Zhao, C.; Yao, M.; Song, Y.; Wu, Y.; Wen, A. Analgesic effect of coumarins from Radix Angelicae pubescentis is mediated by inflammatory factors and TRPV1 in a spared nerve injury model of neuropathic pain. J. Ethnopharmacol. 2017, 195, 81–88. [Google Scholar] [CrossRef]
- Zhang, C.; Hsu, A.C.Y.; Pan, H.; Gu, Y.N.; Zuo, X.; Dong, B.; Wang, Z.Y.; Zheng, J.T.; Lu, J.Y.; Zheng, R.P.; et al. Columbianadin Suppresses Lipopolysaccharide (LPS)-Induced Inflammation and Apoptosis through the NOD1 Pathway. Molecules 2019, 24, 549. [Google Scholar] [CrossRef]
- Chen, S.J.; Bai, Y.; Wang, Y.; Liang, C.X.; Du, K.Z.; Wang, S.Q.; Li, J.; Chang, Y.X. Immunosuppressive effect of Columbianadin on maturation, migration, allogenic T cell stimulation and phagocytosis capacity of TNF-alpha induced dendritic cells. J. Ethnopharmacol. 2022, 285, 114818. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Li, W.; Yang, X.W. Biotransformation of columbianadin by rat hepatic microsomes and inhibition of biotransformation products on NO production in RAW 264.7 cells in vitro. Phytochemistry 2012, 81, 109–116. [Google Scholar] [CrossRef]
- Lu, J.; Fang, K.; Wang, S.; Xiong, L.; Zhang, C.; Liu, Z.; Guan, X.; Zheng, R.; Wang, G.; Zheng, J.; et al. Anti-inflammatory effect of columbianetin on lipopolysaccharide-stimulated human peripheral blood mononuclear cells. Mediat. Inflamm. 2018, 2018, 9191743. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-w.; Guo, Q.-m.; Wang, Y. Absorption and transport of 6 coumarins isolated from the roots of Angelica pubescens f. biserrata in human Caco-2 cell monolayer model. Zhong Xi Yi Jie He Xue Bao = J. Chin. Integr. Med. 2008, 6, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-Q.; Xie, Y.-L.; Gui, S.-H.; Zhang, X.; Mo, Z.-Z.; Sun, C.-Y.; Li, C.-L.; Luo, D.-D.; Zhang, Z.-B.; Su, Z.-R.; et al. Polydatin attenuates D-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 7, 4545–4555. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, Y.; Pi, F.W.; Guo, Y.H.; Cheng, Y.L.; Qian, H. Torularhodin ameliorates oxidative activity in vitro and D-Galactose-induced liver injury via the Nrf2/HO-1 signaling pathway in vivo. J. Agric. Food Chem. 2019, 67, 10059–10068. [Google Scholar] [CrossRef]
- Zhao, N.; Guo, F.-F.; Xie, K.-Q.; Zeng, T. Targeting Nrf-2 is a promising intervention approach for the prevention of ethanol-induced liver disease. Cell. Mol. Life Sci. 2018, 75, 3143–3157. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.Q.; Zhou, Y.D.; Hou, J.G.; Liu, Y.; Wang, Y.P.; Gong, X.J.; Lin, X.H.; Jiang, S.; Wang, Z. Rare Ginsenoside 20(R)-Rg3 Inhibits D-Galactose-Induced Liver and Kidney Injury by Regulating Oxidative Stress-Induced Apoptosis. Am. J. Chin. Med. 2020, 48, 1141–1157. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, C.Y.; Zhang, M.; Lu, X.M.; Cao, W.S.; Xie, C.F.; Li, X.T.; Wu, J.S.; Zhong, C.Y.; Geng, S.S. Protective effects of ginseng stem-leaf saponins on D-galactose-induced reproductive injury in male mice. Aging 2021, 13, 8916–8928. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Kuang, Z.P.; Zhang, J.; Zhao, X.; Pu, P.; Yan, J.F. The preventive effect of Apocynum venetum polyphenols on D-galactose-induced oxidative stress in mice. Exp. Ther. Med. 2020, 19, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.X.; Wang, T.T.; Li, Z.M.; Guo, Y.X.; Granato, D. Green Tea Polyphenols Upregulate the Nrf2 Signaling Pathway and Suppress Oxidative Stress and Inflammation Markers in D-Galactose-Induced Liver Aging in Mice. Front. Nutr. 2022, 9, 836112. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, T.; Li, M.L.; Shi, D.X.; Tan, X.T.; Qiu, F.B. Lycopene attenuates d-galactose-induced insulin signaling impairment by enhancing mitochondrial function and suppressing the oxidative stress/inflammatory response in mouse kidneys and livers. Food Funct. 2022, 13, 7720–7729. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, Y.P.; Lu, M.T.; Yu, L.; Tan, S.M.; Ren, T.Y. Effects of Rosa roxburghii Tratt glycosides and quercetin on D-galactose-induced aging mice model. J. Food Biochem. 2022, 46, e14425. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.F.; Wu, M.M.; Fang, H.; Su, Y.; Zhang, L.L.; Zhou, H. Puerarin Prevents Acute Liver Injury via Inhibiting Inflammatory Responses and ZEB2 Expression. Front. Pharmacol. 2021, 12, 727916. [Google Scholar] [CrossRef] [PubMed]
- Li, M.L.; Zhang, X.H.; Wang, B.; Xu, X.B.; Wu, X.W.; Guo, M.X.; Wang, F.Y. Effect of JAK2/STAT3 signaling pathway on liver injury associated with severe acute pancreatitis in rats. Exp. Ther. Med. 2018, 16, 2013–2021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Y.; Wang, L.K.; Wei, N. Pyrrolidine dithiocarbamate alleviates the anti-tuberculosis drug-induced liver injury through JAK2/STAT3 signaling pathway: An experimental study. Asian Pac. J. Trop. Med. 2017, 10, 493–496. [Google Scholar] [CrossRef]
- Perner, F.; Perner, C.; Ernst, T.; Heidel, F.H. Roles of JAK2 in Aging, Inflammation, Hematopoiesis and Malignant Transformation. Cells 2019, 8, 854. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Z.M.; Cui, H.P.; Wang, T.; Xu, Y.H.; Zhao, J. Urantide prevents CCl4-induced acute liver injury in rats by regulating the MAPK signalling pathway. Mol. Med. Rep. 2021, 24, 688. [Google Scholar] [CrossRef]
- Bataille, A.M.; Manautou, J.E. Nrf2: A Potential Target for New Therapeutics in Liver Disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef]
- Nowak, A.J.; Relja, B. The Impact of Acute or Chronic Alcohol Intake on the NF-κB Signaling Pathway in Alcohol-Related Liver Disease. Int. J. Mol. Sci. 2020, 21, 9407. [Google Scholar] [CrossRef]
- Yan, J.Y.; Shen, S.; He, Y.T.; Li, Z.Q. TLR5 silencing reduced hyperammonaemia-induced liver injury by inhibiting oxidative stress and inflammation responses via inactivating NF-kappa B and MAPK signals. Chem.-Biol. Interact. 2019, 299, 102–110. [Google Scholar] [CrossRef]
- Huang, Q.F.; Bai, F.C.; Nie, J.L.; Lu, S.J.; Lu, C.Y.; Zhu, X.S.; Zhuo, L.; Lin, X. Didymin ameliorates hepatic injury through inhibition of MAPK and NF-kappa B pathways by up-regulating RKIP expression. Int. Immunopharmacol. 2017, 42, 130–138. [Google Scholar] [CrossRef]
- Ma, Z.; Peng, L.; Chu, W.H.; Wang, P.; Fu, Y.Q. Osthole Alleviates D-Galactose-Induced Liver Injury In Vivo via the TLR4/MAPK/NF-kappa B Pathways. Molecules 2023, 28, 443. [Google Scholar] [CrossRef]
- Tao, W.L.; Yue, X.; Ye, R.L.; Nabi, F.; Shang, Y.F.; Zhu, Z.R.; Ahmed, B.Z.; Liu, J. Hepatoprotective Effect of the Penthorum Chinense Pursh Extract against the CCl4-Induced Acute Liver Injury via NF-kappa B and p38-MAPK PATHWAYS in Dogs. Animals 2022, 12, 569. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Weight (g) | Liver Indices (mg/g) | Kidney Indices (mg/g) | |
|---|---|---|---|---|
| Initial | Final | |||
| Control | 31.07 ± 0.68 | 47.17 ± 3.79 d | 17.01 ± 2.34 d | 5.77 ± 0.67 |
| Model | 29.98 ± 0.91 | 33.75 ± 1.19 a | 10.61 ± 0.48 a | 4.94 ± 0.98 |
| CBN (200 mg/kg) | 30.62 ± 1.24 | 35.88 ± 2.93 b | 11.21 ± 1.77 b | 4.74 ± 0.68 |
| CBN (400 mg/kg) | 30.11 ± 1.62 | 35.50 ± 1.29 b | 11.73 ± 0.98 b | 4.48 ± 0.43 |
| CBN (800 mg/kg) | 29.54 ± 0.53 | 35.38 ± 2.75 b | 12.37 ± 0.82 c | 5.01 ± 0.38 |
| Vitamin E (150 mg/kg) | 30.67 ± 0.78 | 37.33 ± 3.40 c | 12.63 ± 0.30 c | 5.11 ± 0.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Peng, L.; Sheng, Y.; Chu, W.; Fu, Y. Anti-Inflammatory Effect of Columbianadin against D-Galactose-Induced Liver Injury In Vivo via the JAK2/STAT3 and JAK2/p38/NF-κB Pathways. Pharmaceuticals 2024, 17, 378. https://0-doi-org.brum.beds.ac.uk/10.3390/ph17030378
Ma Z, Peng L, Sheng Y, Chu W, Fu Y. Anti-Inflammatory Effect of Columbianadin against D-Galactose-Induced Liver Injury In Vivo via the JAK2/STAT3 and JAK2/p38/NF-κB Pathways. Pharmaceuticals. 2024; 17(3):378. https://0-doi-org.brum.beds.ac.uk/10.3390/ph17030378
Chicago/Turabian StyleMa, Zhe, Lin Peng, Yaoyao Sheng, Wenhui Chu, and Yongqian Fu. 2024. "Anti-Inflammatory Effect of Columbianadin against D-Galactose-Induced Liver Injury In Vivo via the JAK2/STAT3 and JAK2/p38/NF-κB Pathways" Pharmaceuticals 17, no. 3: 378. https://0-doi-org.brum.beds.ac.uk/10.3390/ph17030378