Pancreatic Islets Exhibit Dysregulated Adaptation of Insulin Secretion after Chronic Epinephrine Exposure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Preparations
2.2. Pancreatic Islet Isolation and Incubation
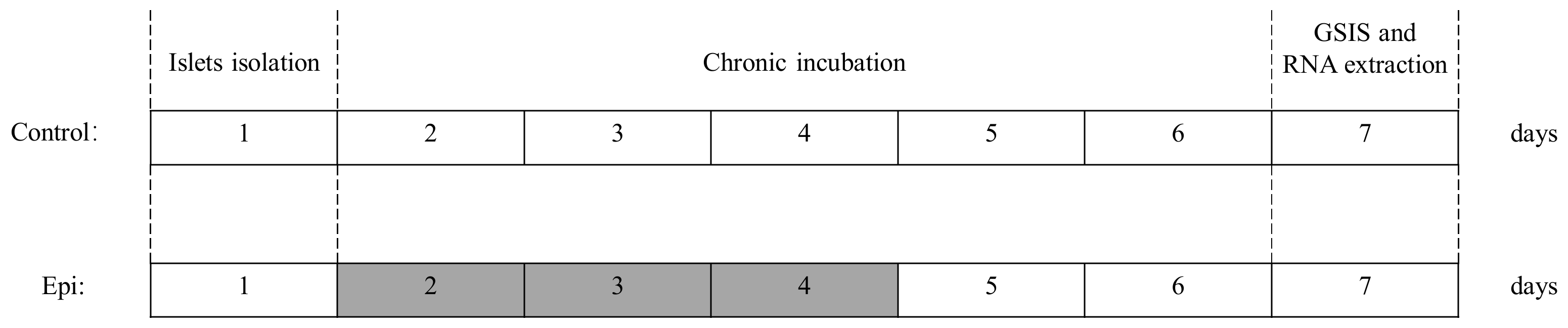
2.3. Chronic Epinephrine Exposure Experimental Design
2.4. Islets Insulin Responsiveness Assessment
2.5. Quantitative Real Time PCR
2.6. Statistical Analysis
3. Results
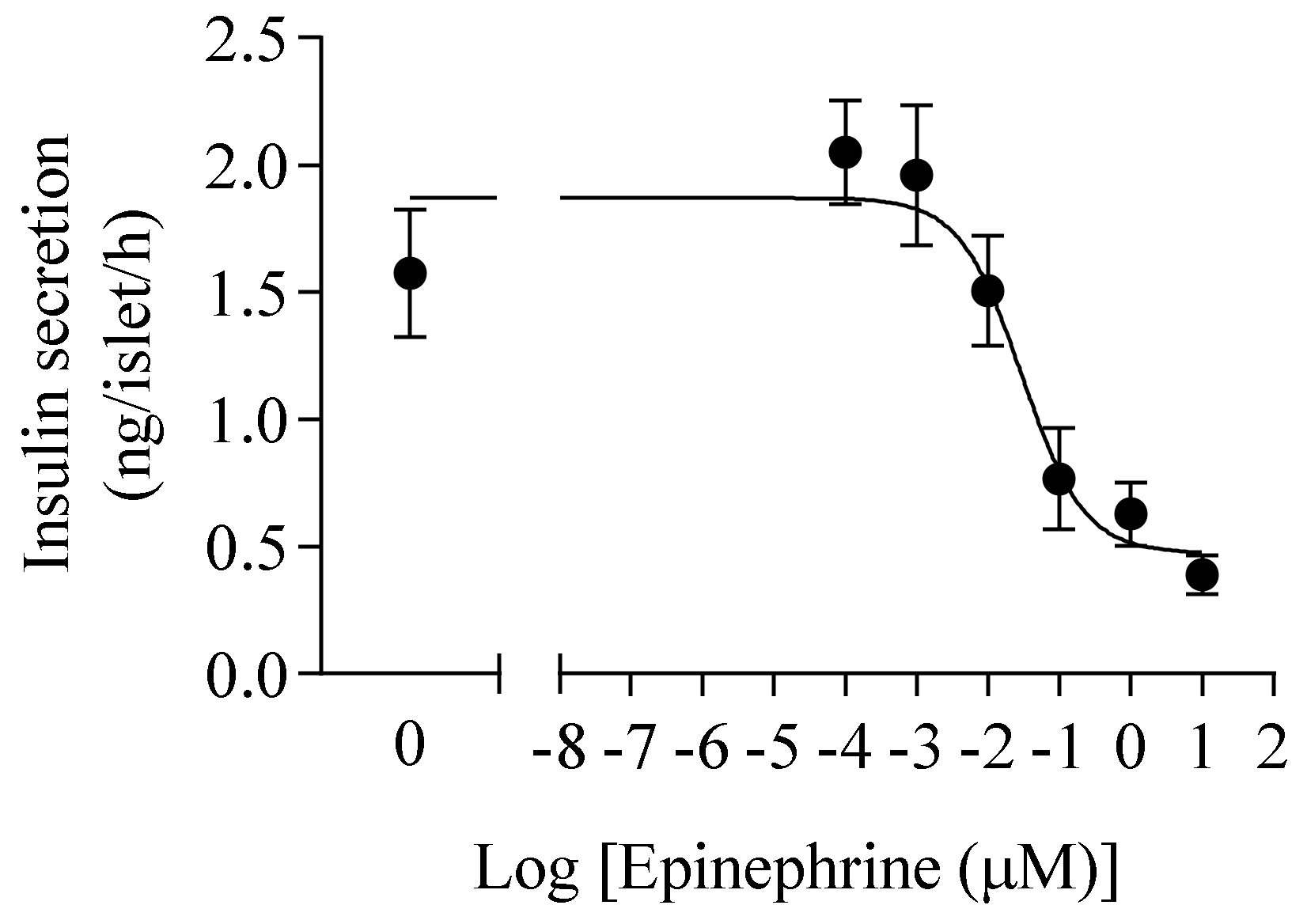
3.1. Determination Epinephrine Half Maximal Inhibitory Concentration on Insulin Secretion
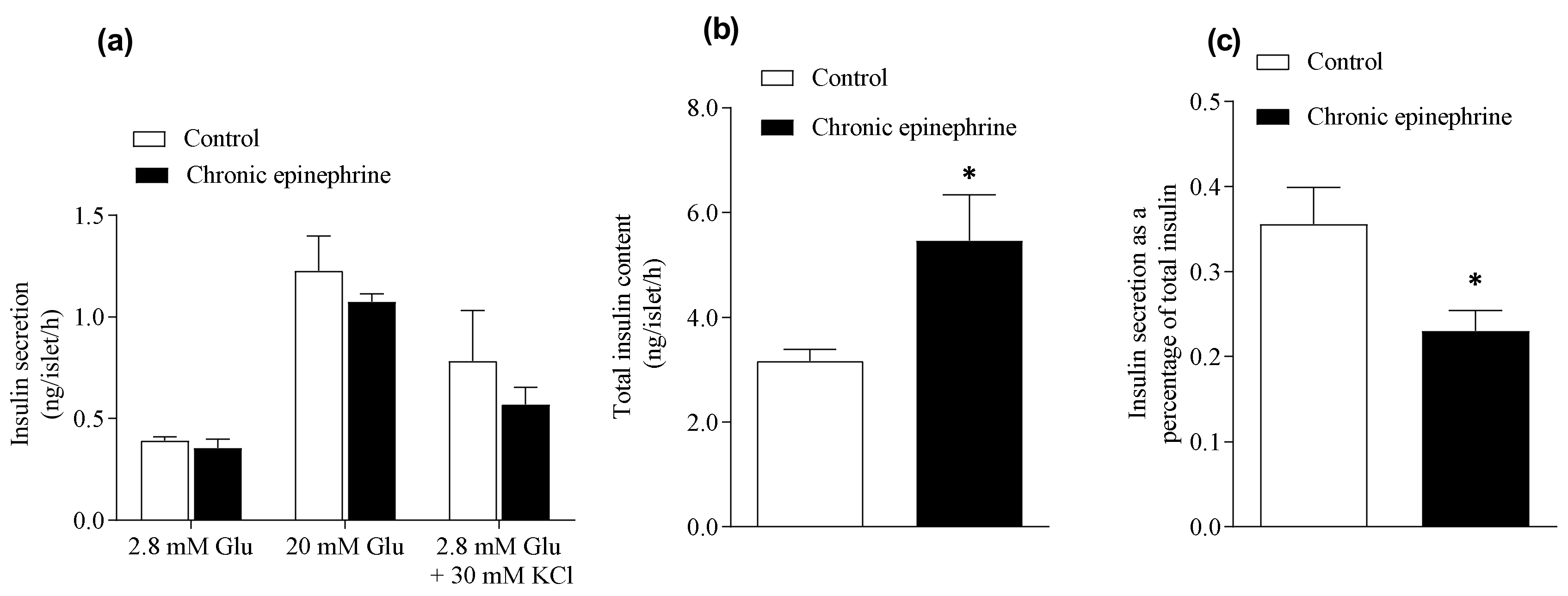
3.2. Total Insulin Content Was Higher and Fractional Insulin Release Was Lower after Chronic Epinephrine Incubation
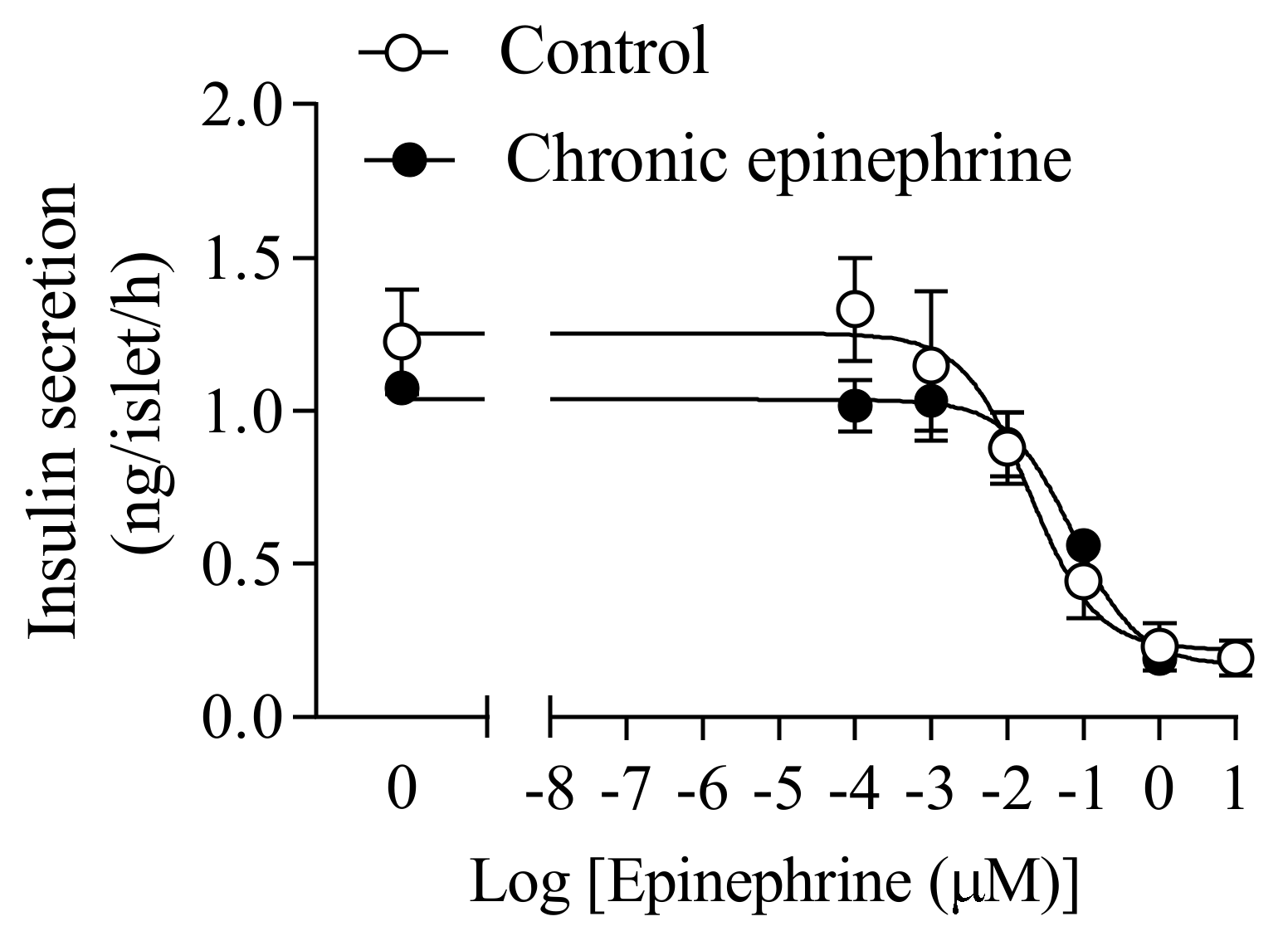
3.3. Half Maximal Inhibitory Concentration of Epinephrine Was Higher after Chronic Exposure
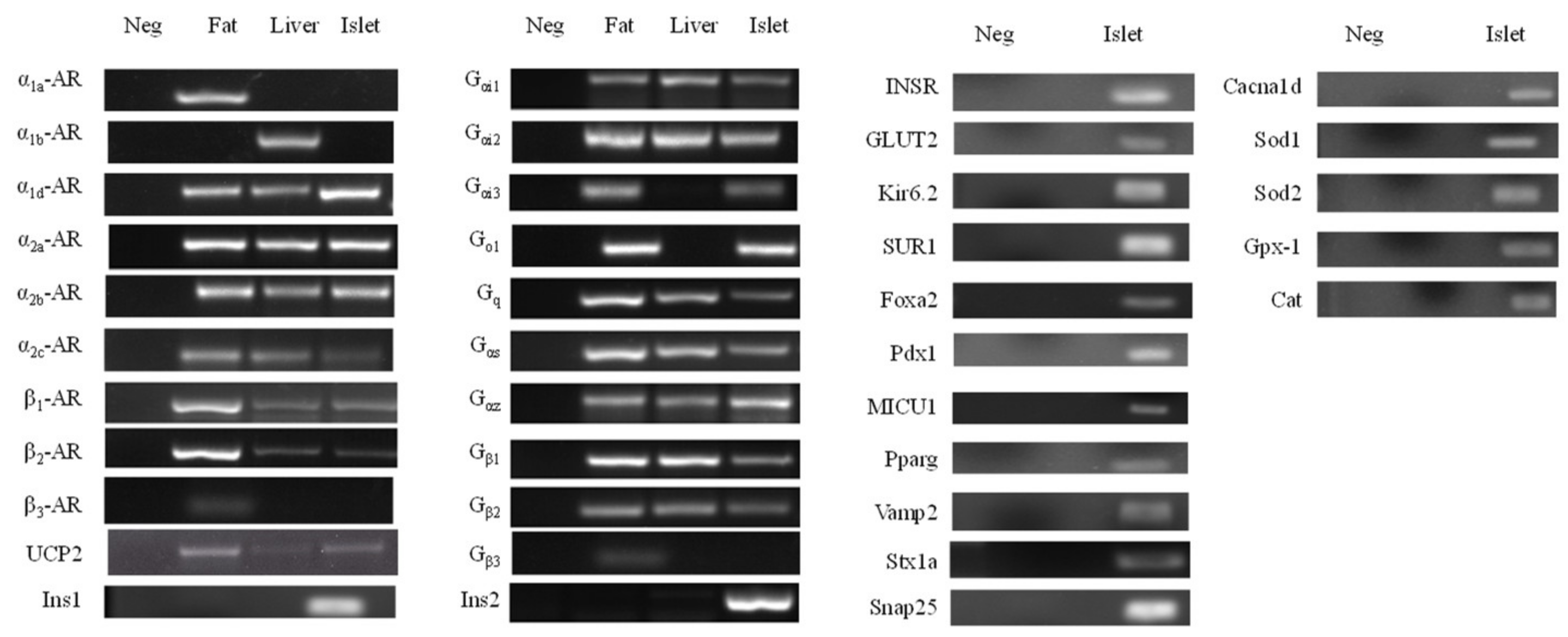
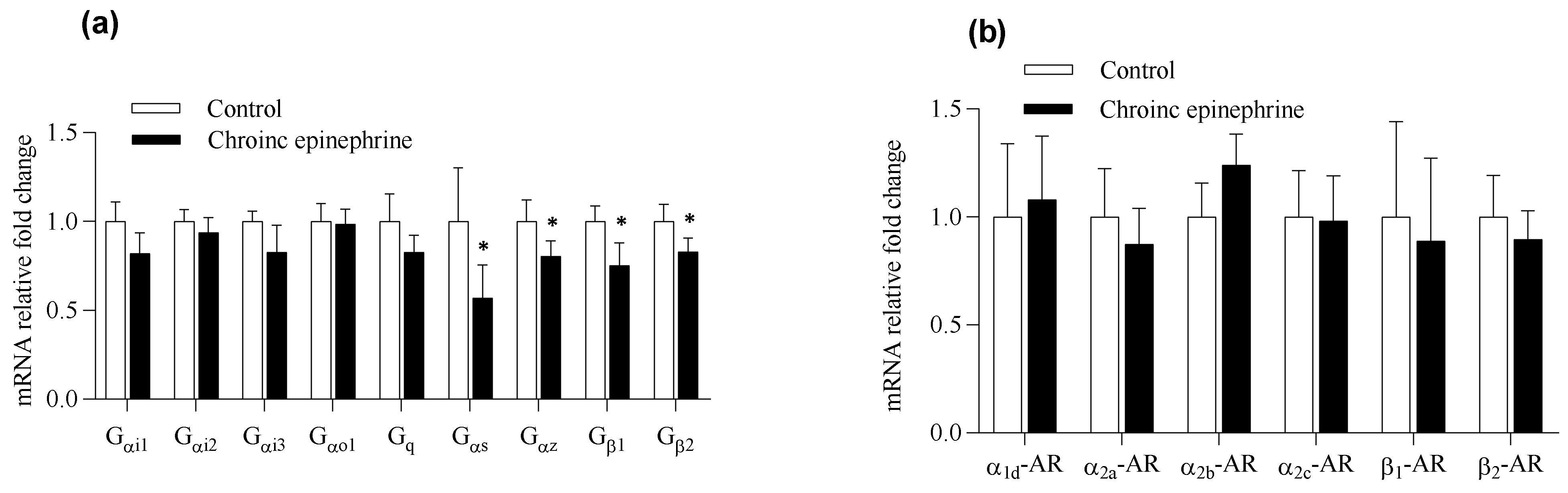
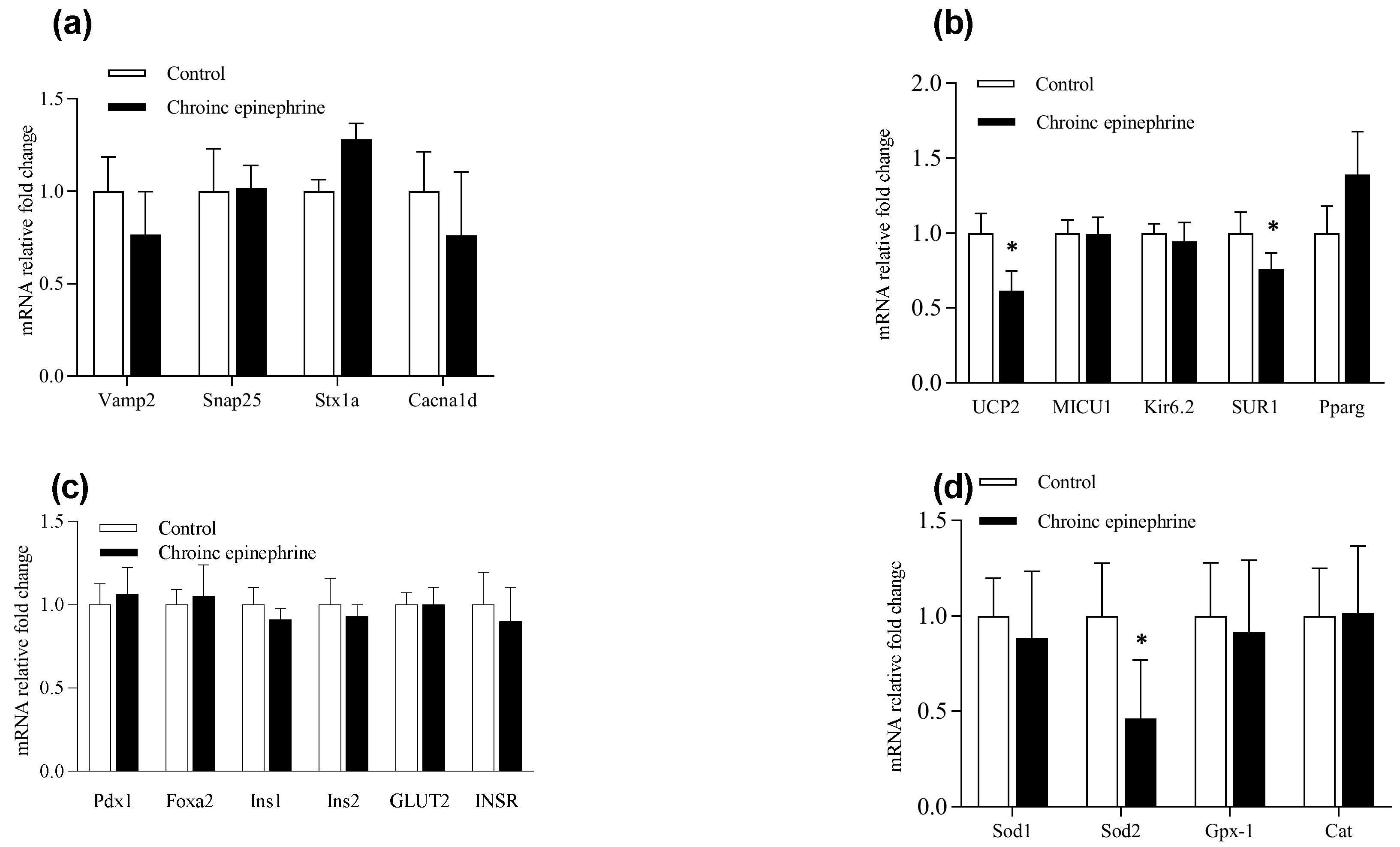
3.4. Expression Profile of mRNA in Pancreatic Islets
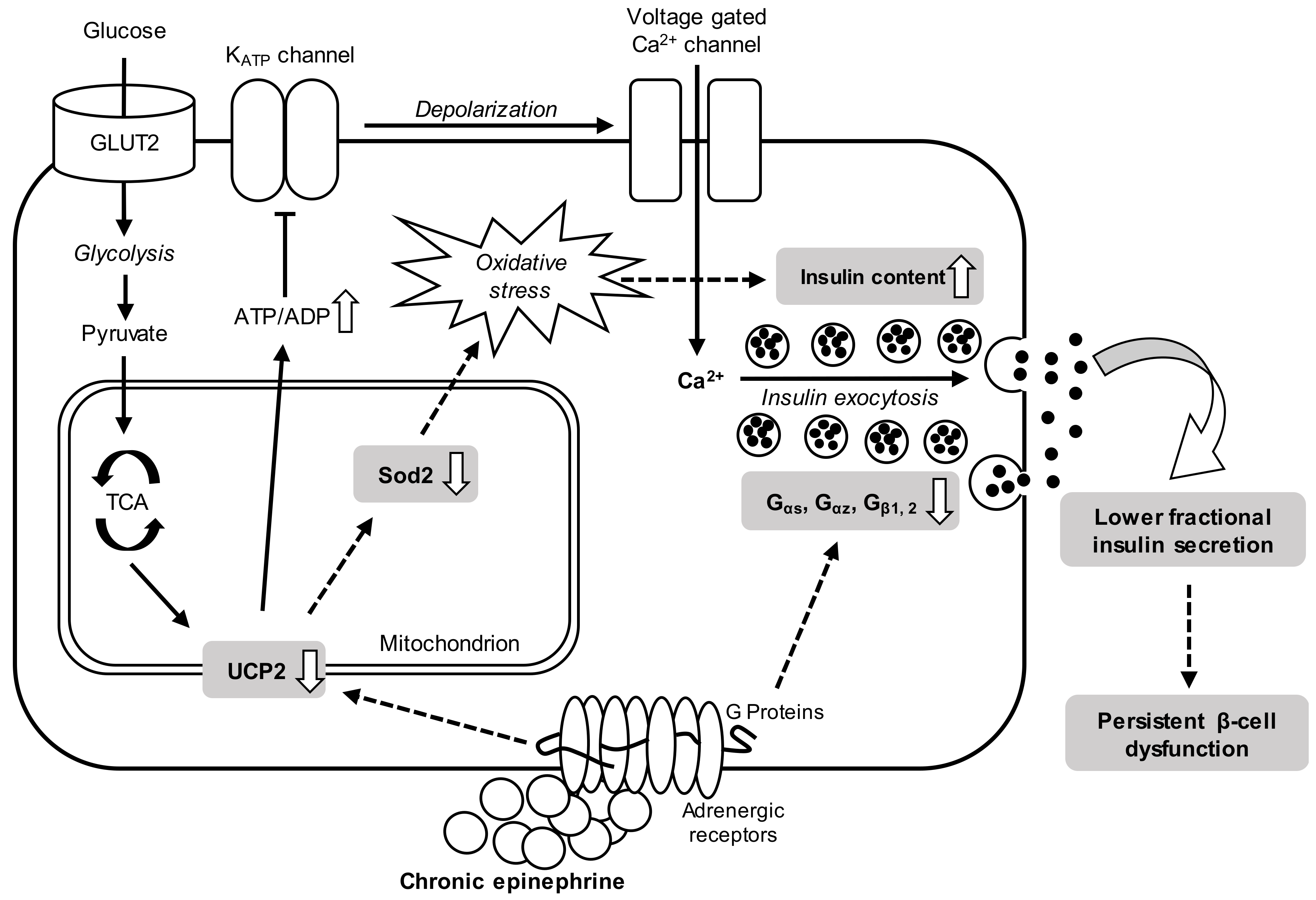
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Limesand, S.W.; Rozance, P.J. Fetal adaptations in insulin secretion result from high catecholamines during placental insufficiency. J. Physiol. 2017, 595, 5103–5113. [Google Scholar] [CrossRef]
- Macko, A.R.; Yates, D.T.; Chen, X.; Green, A.S.; Kelly, A.C.; Brown, L.D.; Limesand, S.W. Elevated Plasma Norepinephrine Inhibits Insulin Secretion, but Adrenergic Blockade Reveals Enhanced Beta-Cell Responsiveness in an Ovine Model of Placental Insufficiency at 0.7 of Gestation. J. Dev. Orig. Health Dis. 2013, 4, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Leos, R.A.; Anderson, M.J.; Chen, X.; Pugmire, J.; Anderson, K.A.; Limesand, S.W. Chronic exposure to elevated norepinephrine suppresses insulin secretion in fetal sheep with placental insufficiency and intrauterine growth restriction. Am. J. Physiol. Metab. 2010, 298, E770–E778. [Google Scholar] [CrossRef]
- Chen, X.; Kelly, A.C.; Yates, D.T.; Macko, A.R.; Lynch, R.M.; Limesand, S.W. Islet adaptations in fetal sheep persist following chronic exposure to high norepinephrine. J. Endocrinol. 2017, 232, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Strathmann, M.; Wilkie, T.M.; Simon, M.I. Diversity of the G-protein family: Sequences from five additional alpha subunits in the mouse. Proc. Natl. Acad. Sci. USA 1989, 86, 7407–7409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinoff, P.B. Alpha- and Beta-Adrenergic Receptor Subtypes Properties, Distribution and Regulation. Drugs 1984, 28 (Suppl. S2), 1–15. [Google Scholar] [CrossRef] [PubMed]
- Straub, S.G.; Sharp, G.W.G. Evolving insights regarding mechanisms for the inhibition of insulin release by norepinephrine and heterotrimeric G proteins. Am. J. Physiol. Physiol. 2012, 302, C1687–C1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, M.; McDermott, A.M.; Gillison, S.L.; Sharp, G.W. Time Course of Action of Pertussis Toxin to Block the Inhibition of Stimulated Insulin Release by Norepinephrine. Endocrinology 1995, 136, 1857–1863. [Google Scholar] [CrossRef]
- Sassmann, A.; Gier, B.; Gröne, H.-J.; Drews, G.; Offermanns, S.; Wettschureck, N. The Gq/G11-mediated signaling pathway is critical for autocrine potentiation of insulin secretion in mice. J. Clin. Investig. 2010, 120, 2184–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, A.C.; Camacho, L.E.; Pendarvis, K.; Davenport, H.M.; Steffens, N.R.; Smith, K.E.; Weber, C.S.; Lynch, R.M.; Papas, K.K.; Limesand, S.W. Adrenergic receptor stimulation suppresses oxidative metabolism in isolated rat islets and Min6 cells. Mol. Cell. Endocrinol. 2018, 473, 136–145. [Google Scholar] [CrossRef]
- Stefan, K.; Zhang, C.; Scorrano, L.; Dalgaard, L.T.; St-Pierre, J.; Grey, S.T.; Lowell, B.B. Superoxide-Mediated Activation of Uncoupling Protein 2 Causes Pancreatic Β Cell Dysfunction. J. Clin. Investig. 2003, 112, 1831–1842. [Google Scholar]
- Kelly, A.C.; Bidwell, C.A.; Chen, X.; Macko, A.R.; Anderson, M.J.; Limesand, S.W. Chronic Adrenergic Signaling Causes Abnormal RNA Expression of Proliferative Genes in Fetal Sheep Islets. Endocrinology 2018, 159, 3565–3578. [Google Scholar] [CrossRef] [Green Version]
- O’Driscoll, L.; Gammell, P.; McKiernan, E.; Ryan, E.; Jeppesen, P.B.; Rani, S.; Clynes, M. Phenotypic and global gene expression profile changes between low passage and high passage MIN-6 cells. J. Endocrinol. 2006, 191, 665–676. [Google Scholar] [CrossRef]
- Chen, X.; Fahy, A.L.; Green, A.S.; Anderson, M.J.; Rhoads, R.P.; Limesand, S.W. Beta2-Adrenergic Receptor Desensitization in Perirenal Adipose Tissue in Fetuses and Lambs with Placental Insufficiency-Induced Intrauterine Growth Restriction. J. Physiol. 2010, 588 Pt 18, 3539–3549. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, X.; Green, A.S.; Macko, A.R.; Yates, D.T.; Kelly, A.C.; Limesand, S.W. Enhanced insulin secretion responsiveness and islet adrenergic desensitization after chronic norepinephrine suppression is discontinued in fetal sheep. Am. J. Physiol. Metab. 2014, 306, E58–E64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bawa, T.; Altememi, G.F.; Eikenburg, D.C.; Standifer, K.M. Desensitization of Alpha 2a-Adrenoceptor Signalling by Modest Levels of Adrenaline Is Facilitated by Beta 2-Adrenoceptor-Dependent Grk3 up-Regulation. Br. J. Pharmacol. 2003, 138, 921–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Chen, M.; Zhang, Q.H.; Ma, Z.; Weinstein, L.S. Beta Cell-Specific Deficiency of the Stimulatory G Protein Alpha-Subunit Gsalpha Leads to Reduced Beta Cell Mass and Insulin-Deficient Diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 19601–19606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimple, M.E.; Joseph, J.W.; Bailey, C.L.; Fueger, P.T.; Hendry, I.A.; Newgard, C.B.; Casey, P.J. Galphaz Negatively Regulates Insulin Secretion and Glucose Clearance. J. Biol. Chem. 2008, 283, 4560–4567. [Google Scholar] [CrossRef] [Green Version]
- Kimple, M.E.; Nixon, A.B.; Kelly, P.; Bailey, C.L.; Young, K.H.; Fields, T.A.; Casey, P.J. A Role for G(Z) in Pancreatic Islet Beta-Cell Biology. J. Biol. Chem. 2005, 280, 31708–31713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Fang, Q.; Straub, S.G.; Lindau, M.; Sharp, G.W. Noradrenaline Inhibits Exocytosis Via the G Protein Betagamma Subunit and Refilling of the Readily Releasable Granule Pool Via the Alpha(I1/2) Subunit. J. Physiol. 2010, 588 Pt 18, 3485–3498. [Google Scholar] [CrossRef]
- Parnaud, G.; Bosco, D.; Berney, T.; Pattou, F.; Kerr-Conte, J.; Donath, M.Y.; Bruun, C.; Mandrup-Poulsen, T.; Billestrup, N.; Halban, P.A. Proliferation of sorted human and rat beta cells. Diabetologia 2007, 51, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliasson, L.; Abdulkader, F.; Braun, M.; Galvanovskis, J.; Hoppa, M.B.; Rorsman, P. Novel aspects of the molecular mechanisms controlling insulin secretion. J. Physiol. 2008, 586, 3313–3324. [Google Scholar] [CrossRef]
- Supale, S.; Li, N.; Brun, T.; Maechler, P. Mitochondrial Dysfunction in Pancreatic Beta Cells. Trends Endocrinol. Metab. 2012, 23, 477–487. [Google Scholar] [CrossRef]
- Kelly, A.C.; Bidwell, C.A.; McCarthy, F.M.; Taska, D.J.; Anderson, M.J.; Camacho, L.E.; Limesand, S.W. RNA Sequencing Exposes Adaptive and Immune Responses to Intrauterine Growth Restriction in Fetal Sheep Islets. Endocrinology 2017, 158, 743–755. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are Oxidative Stress-Activated Signaling Pathways Mediators of Insulin Resistance and Beta-Cell Dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echtay, K.S.; Roussel, D.; St-Pierre, J.; Jekabsons, M.B.; Cadenas, S.; Stuart, J.A.; Harper, J.A.; Roebuck, S.J.; Morrison, A.; Pickering, S.; et al. Superoxide activates mitochondrial uncoupling proteins. Nat. Cell Biol. 2002, 415, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Collins, S. Reactive Oxygen Species and Uncoupling Protein 2 in Pancreatic Beta-Cell Function. Diabetes Obes. Metab. 2010, 12 (Suppl. S2), 141–148. [Google Scholar] [CrossRef]
- Produit-Zengaffinen, N.; Davis-Lameloise, N.; Perreten, H.; Bécard, D.; Gjinovci, A.; Keller, P.A.; Wollheim, C.B.; Herrera, P.; Muzzin, P.; Assimacopoulos-Jeannet, F. Increasing uncoupling protein-2 in pancreatic beta cells does not alter glucose-induced insulin secretion but decreases production of reactive oxygen species. Diabetologia 2006, 50, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Affourtit, C.; Brand, M.D. On the role of uncoupling protein-2 in pancreatic beta cells. Biochim. Biophys. Acta 2008, 1777, 973–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedge, M.; Lortz, S.; Drinkgern, J.; Lenzen, S. Relation between Antioxidant Enzyme Gene Expression and Antioxidative Defense Status of Insulin-Producing Cells. Diabetes 1997, 46, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Dai, C.; Lustig, M.E.; Bonner, J.S.; Mayes, W.H.; Mokshagundam, S.; James, F.D.; Thompson, C.S.; Lin, C.-T.; Perry, C.G.; et al. Heterozygous SOD2 Deletion Impairs Glucose-Stimulated Insulin Secretion, but Not Insulin Action, in High-Fat–Fed Mice. Diabetes 2014, 63, 3699–3710. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Product Size (bp) | Accession Number |
|---|---|---|---|---|
| β-actin | GTGGGTATGGGTCAGAAGGAC | TGTGGTGCCAAATCTTCTCCA | 133 | NM_031144.3 |
| G protein subunits | ||||
| Gαi1 | AACCCAGCAGGATGTTCTCAG | CCTCAGCAAGAACCAGGTCATA | 203 | NM_013145.1 |
| Gαi2 | GCTGTTCGCACTGTCCTGT | GACGATGCCTGTGGTCTTCA | 241 | NM_031035.3 |
| Gαi3 | GATGATGCCCGACAGTTATTTG | CTTGAAGGTGAAGTGGGTCTCC | 273 | NM_013106.1 |
| Gαo1 | TGTCGCACTCAGCGGCTAT | GAAAGCAGATGGTCAAGGGTG | 191 | NM_017327.1 |
| Gαz | TCAAGATGGTGGATGTGGGA | TTCAGGAAGAGGATGAGGGAG | 220 | NM_013189.2 |
| Gαs | TGCCCAGGAAGACCGTTG | GCCGATTTGTGGCGTGAC | 241 | NM_001024823.4 |
| Gq | TGAAGACAAGAGGGGCTTTACC | CTCGCCGTCTATCGTAGCATT | 242 | NM_031036.1 |
| Gβ1 | TCCGAGAAGGGATGTGCC | TTGAAGTCGTCATAGCCAGCG | 241 | NM_030987.2 |
| Gβ2 | GTGCCGACAGACATTCATAGGT | TCAGCCCGCAGGTCAAAC | 123 | NM_031037.2 |
| Gβ3 | GCTCTGGGATGTGAGGGAAG | ACTGAGTGAGAAGGCTACGGAC | 217 | NM_021858.3 |
| Adrenergic receptors | ||||
| α1d-AR | GACCAGCGCCAAAGGATA | TGAAGTAGCCCAGCCAGAA | 245 | NM_024483.1 |
| α2a-AR | GGTGTGTTGGTTCCCGTTCT | CGGAAGTCGTGGTTGAAAATG | 150 | NM_012739.3 |
| α2b- AR | CAGCCTCAGACTTCTCGGGTA | TAGATGACAGGGTTCAAAGAG | 293 | NM_138505.2 |
| α2c-AR | TGCTCTTCTGCACCTCGTCC | GATGACAGCCGAGATGAGCC | 152 | NM_138506.1 |
| β1-AR | CCGATCTGGTCATGGGACT | GCAGGCTCTGGTAGCGAAA | 121 | NM_012701.1 |
| β2-AR | GCCACGACATCACTCAGGAA | CCAGAACTCGCACCAGAAAT | 266 | NM_012492.2 |
| α1a-AR | CGTGGTGGGTTGCTTCGT | AGACACTGGATTCGCAGGACA | 210 | NM_017191.2 |
| α1b-AR | CCTTGGGCATTGTAGTCGGA | GCACGGGTAGATGATGGGAT | 167 | NM_016991.2 |
| Signals related to mitochondria and ATP synthesis | ||||
| MICU1 | AGCCTACTCCACACCAGACAA | CGTTCCTGGGCAATTTTCTTTC | 198 | NM_199412.1 |
| Pparg | GAGGGCGATCTTGACAGGAA | ACAGCTTCCACGGATCGAAA | 180 | NM_013124.3 |
| Kir6.2 | ACCACGTCATCGACTCCAAC | GAATAGCGGCCATCCTCCTC | 208 | NM_031358.3 |
| SUR1 | TCTTCACCTGGACCCCTGAT | TTCTCCCTCGCTGTCTGGAA | 194 | AF039595.1 |
| UCP2 | CTGGGCACCATCCTAACC | GGAAGCGGACCTTTACCA | 247 | NM_019354.3 |
| Signals related to insulinogenesis | ||||
| Ins1 | CCAAGTCCCGTCGTGAAGT | CTCCAGTTGGTAGAGGGAGC | 164 | NM_019129.3 |
| Ins2 | ACAGCACCTTTGTGGTTCTCA | CAGTGCCAAGGTCTGAAGGT | 166 | NM_019130.2 |
| Foxa2 | GACTGAGGTGGGTAGCCAGAA | CACGGCTCCCAGCATACTTTA | 162 | NM_012743.1 |
| Pdx1 | GAACGCTGGAACAGGGAAGT | CCAGTCTCGGTTCCATTCG | 164 | NM_022852.3 |
| INSR | CAGTTTGTGGAACGGTGCTG | TGGTAGGGTCATCGGGTTCT | 142 | NM_017071.2 |
| GLUT2 | TTGGCTCAGCAGTTCTCTGG | CGGCACAGAAAAACATGCCA | 208 | NM_012879.2 |
| Signals related to endocytosis and exocytosis | ||||
| Vamp2 | TGAGGGTGAATGTGGACAAGG | GGGAGGGGGCTGAAAGATATG | 306 | NM_012663.2 |
| Stx1a | GAGGAAGGTCTGAACCGCTC | GTGCCTGGTCTCGATCTCAC | 291 | NM_053788.2 |
| Snap25 | ATTGAGGAAGGGATGGACCAAA | AGCTTGTTACAGGGACACACA | 107 | NM_030991.3 |
| Cacna1d | GAGGAGGGCAAACGAAACAC | CAAGTGGGCTGAGAACCTAGA | 285 | NM_017298.1 |
| Signals related to oxidative damage | ||||
| Sod1 | GCGTCATTCACTTCGAGCAG | CCTCTCTTCATCCGCTGGAC | 191 | NM_017050.1 |
| Sod2 | ACGCGACCTACGTGAACAAT | GCCTCCAGCAACTCTCCTTT | 196 | NM_017051.2 |
| Gpx-1 | AGTGCGAGGTGAATGGTGAG | TCGATGTCGATGGTGCGAAA | 226 | NM_030826.4 |
| Cat | GAGGAAACGCCTGTGTGAGA | TTGGCAGCTATGTGAGAGCC | 201 | NM_012520.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Huang, H.; Limesand, S.W.; Chen, X. Pancreatic Islets Exhibit Dysregulated Adaptation of Insulin Secretion after Chronic Epinephrine Exposure. Curr. Issues Mol. Biol. 2021, 43, 240-250. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010020
Li R, Huang H, Limesand SW, Chen X. Pancreatic Islets Exhibit Dysregulated Adaptation of Insulin Secretion after Chronic Epinephrine Exposure. Current Issues in Molecular Biology. 2021; 43(1):240-250. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010020
Chicago/Turabian StyleLi, Rui, Huichai Huang, Sean W. Limesand, and Xiaochuan Chen. 2021. "Pancreatic Islets Exhibit Dysregulated Adaptation of Insulin Secretion after Chronic Epinephrine Exposure" Current Issues in Molecular Biology 43, no. 1: 240-250. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010020