Poly(ADP-Ribose) Polymerase Inhibitor PJ34 Reduces Brain Damage after Stroke in the Neonatal Mouse Brain

 , , and
, , and 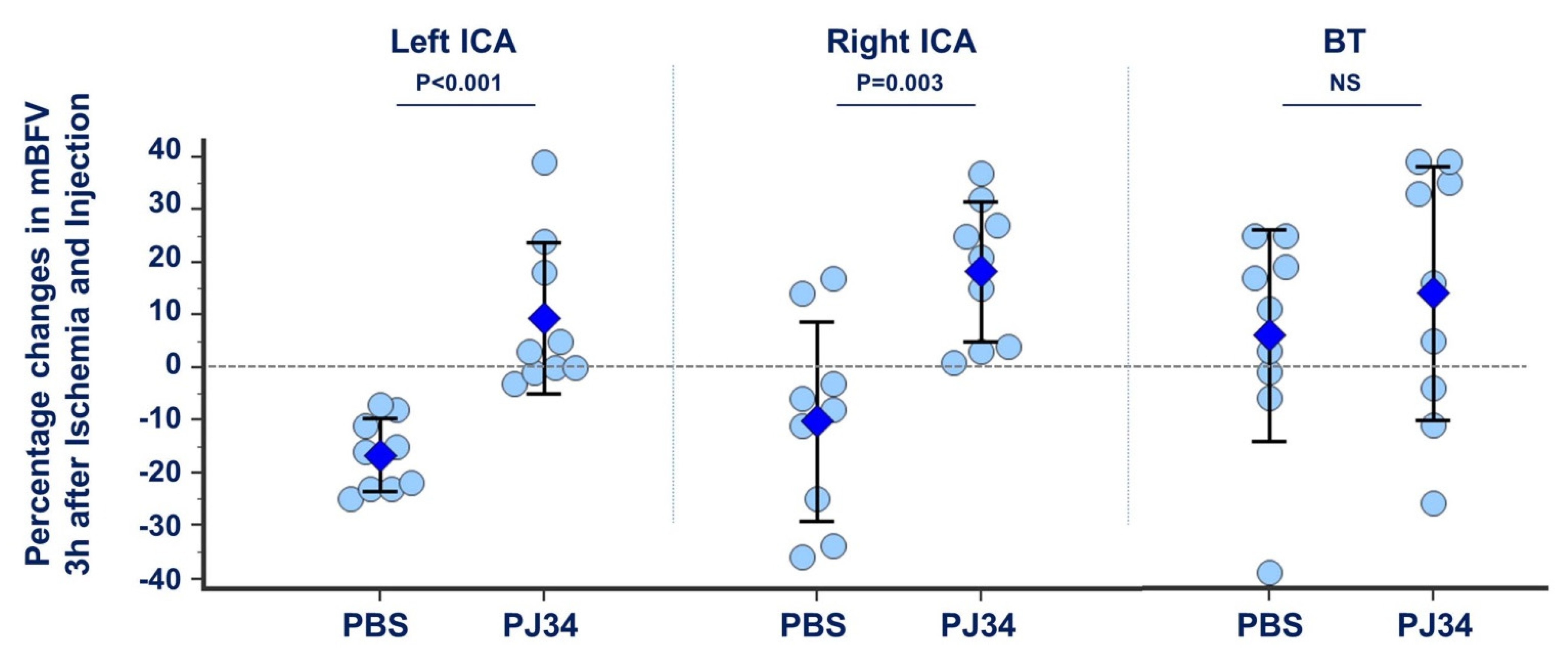{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
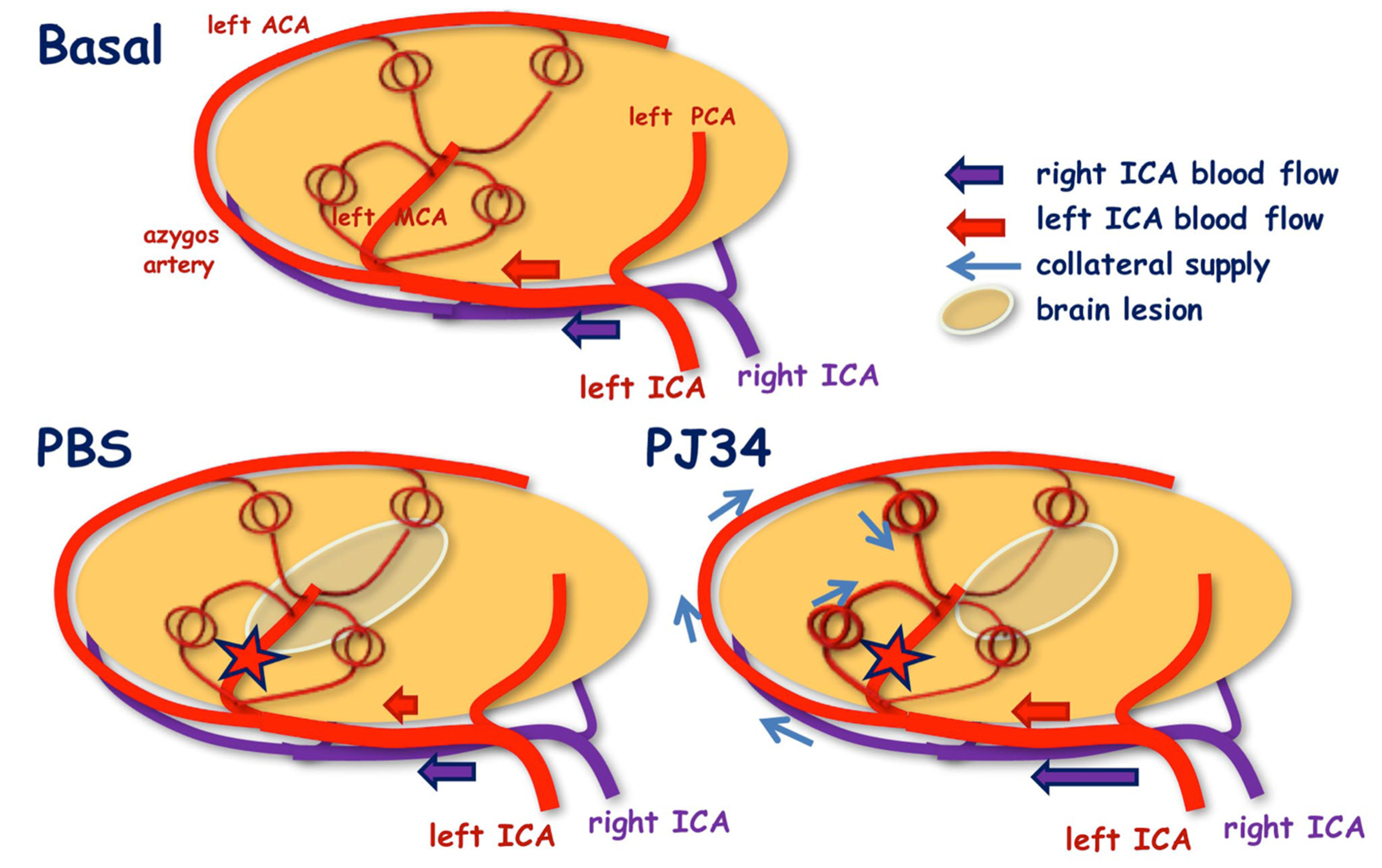
2.1. Collateral Recruitment by PJ34 after Neonatal Ischemia
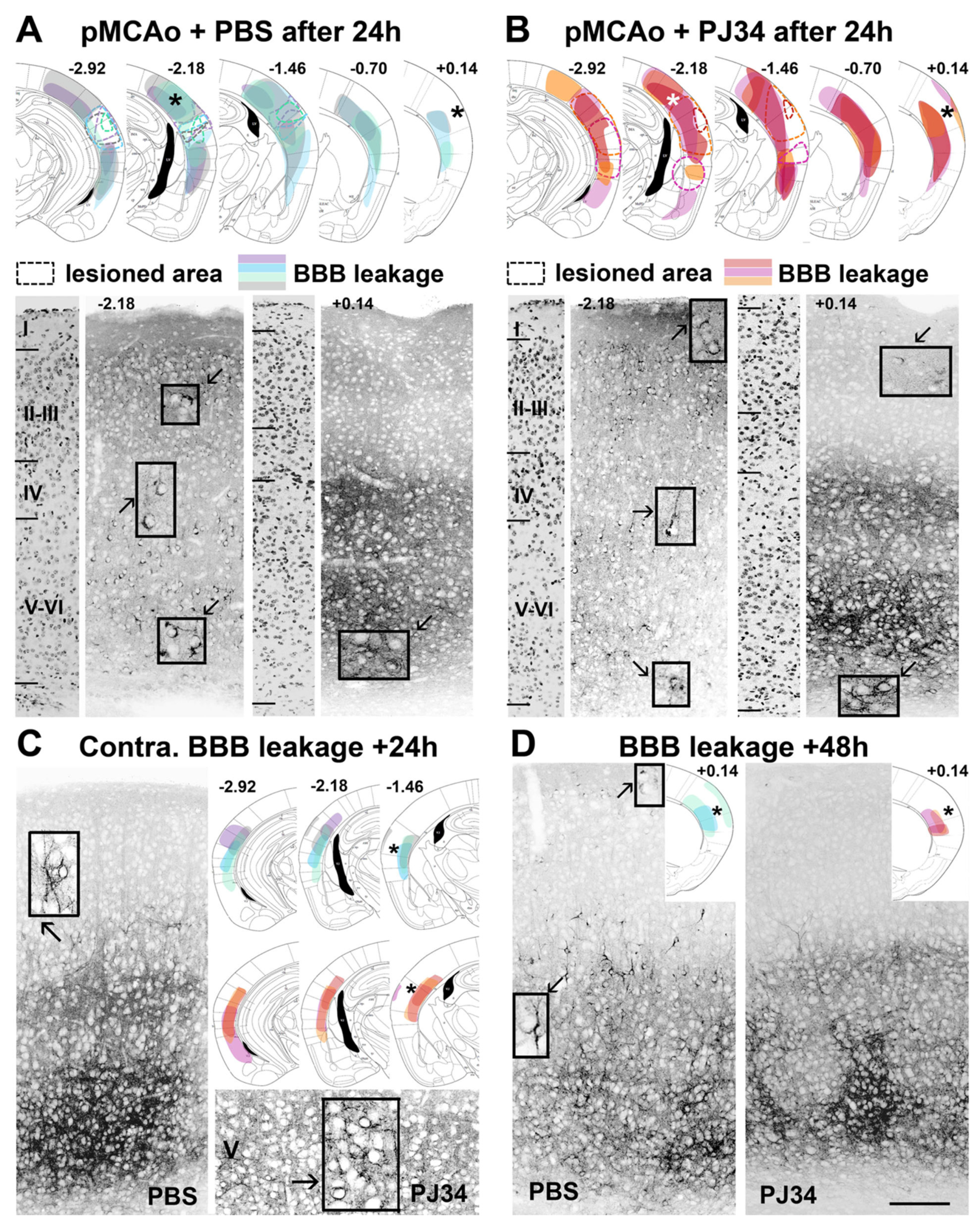
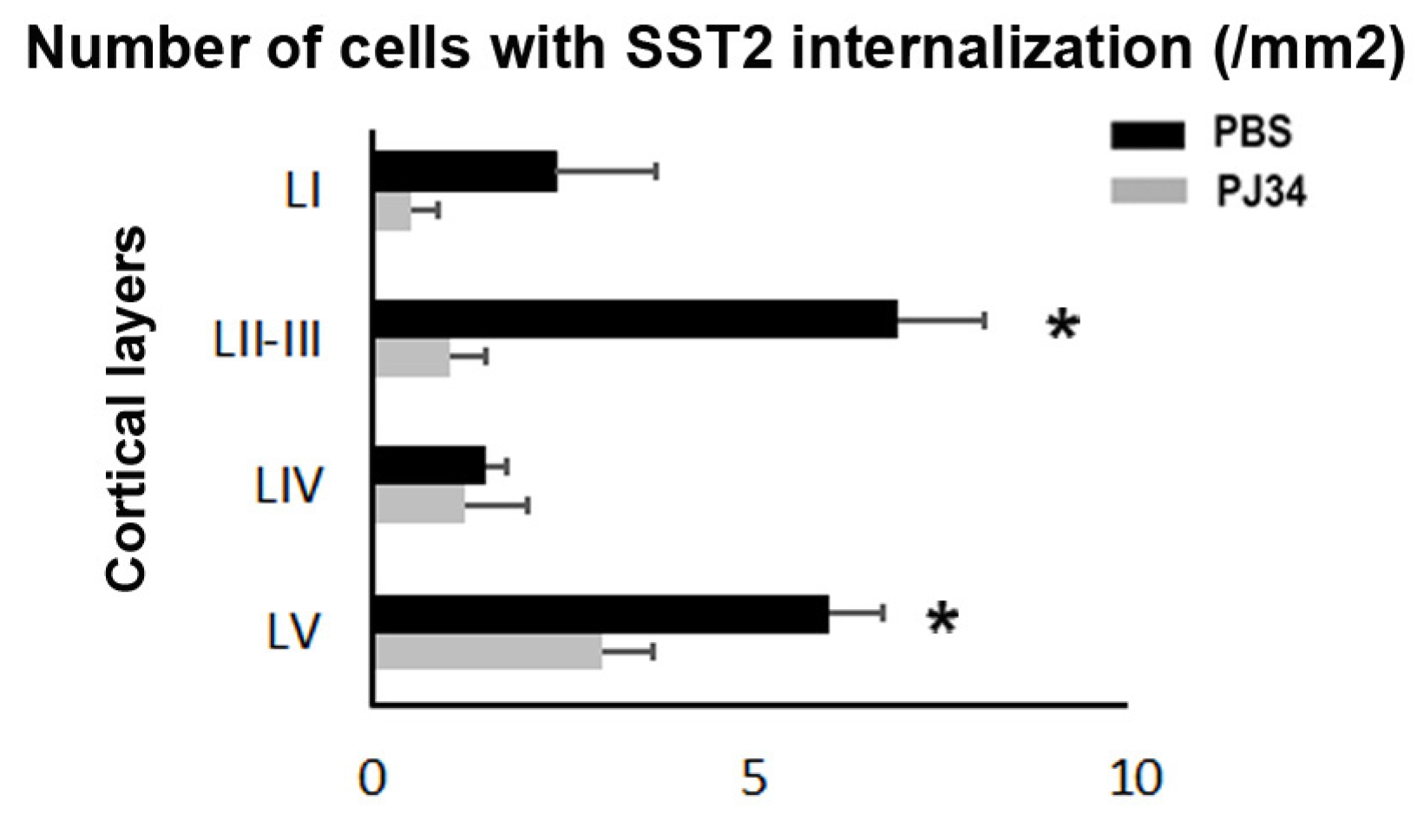
2.2. PJ34 Reduces BBB Permeability in the Anterior Brain
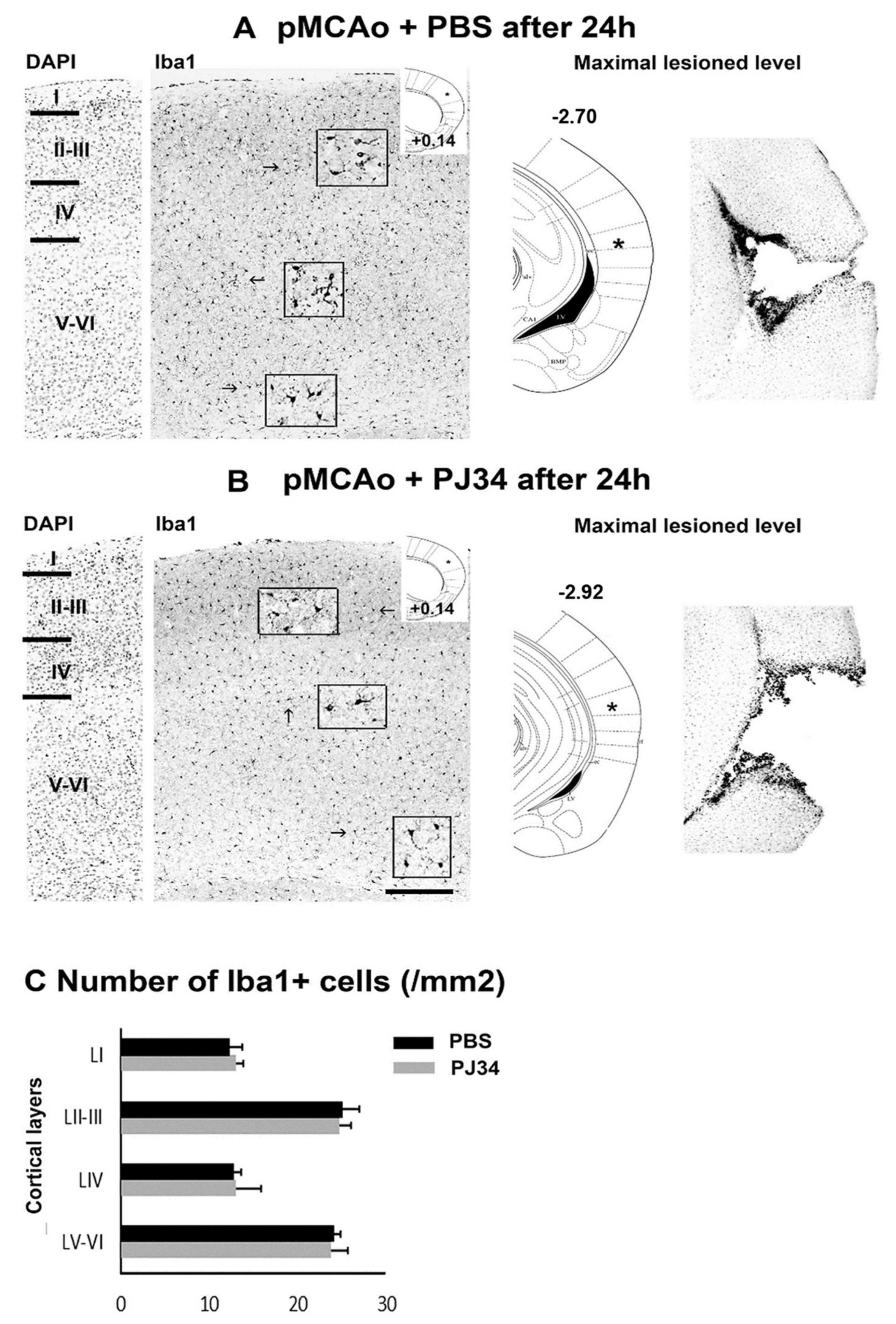
2.3. PJ34 and Microglia in the Anterior and Posterior Brain
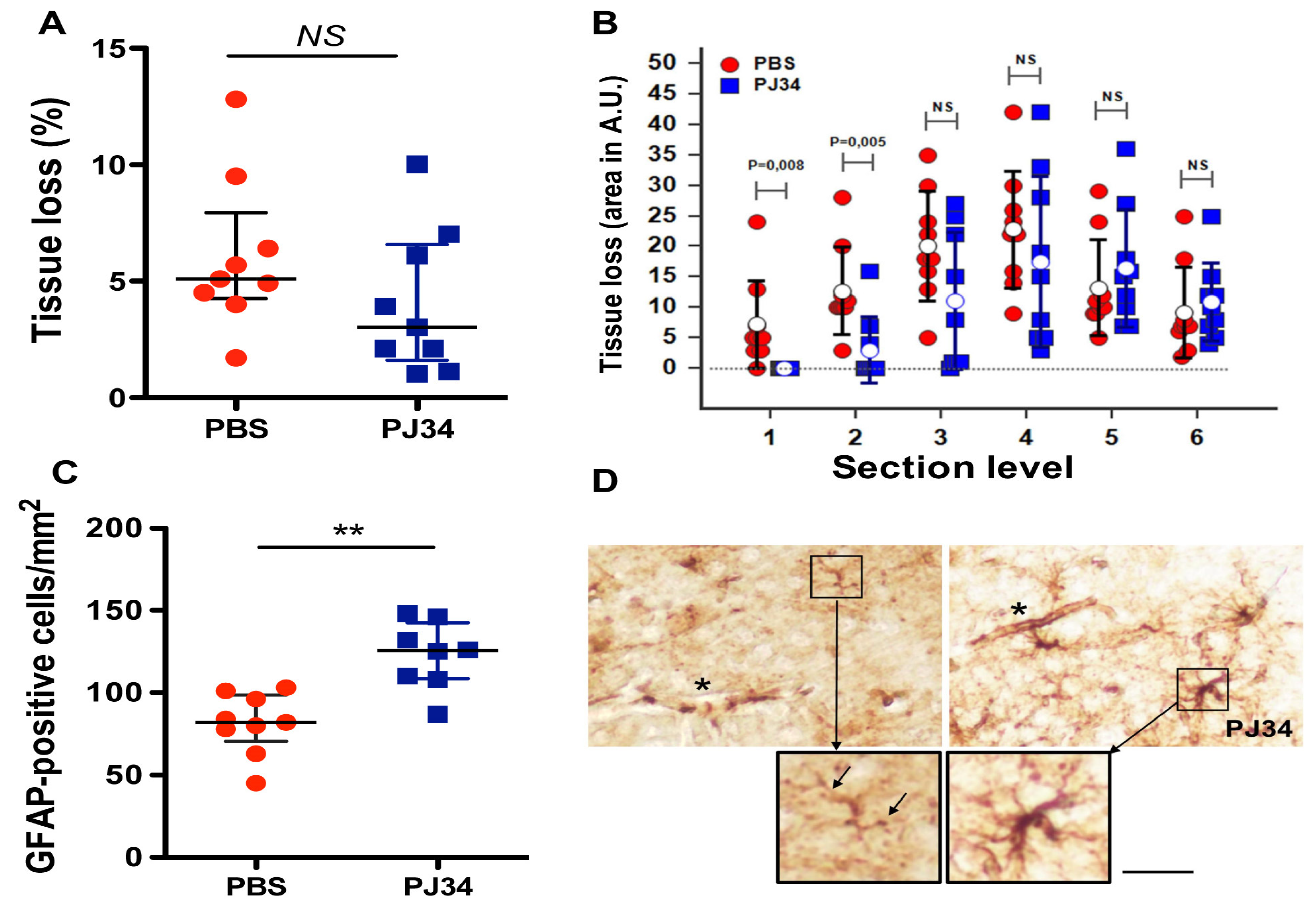
2.4. PJ34 Reduces Tissue Loss and Astrocyte Demise in the Anterior Brain
3. Discussion
4. Materials and Methods
4.1. Ethic Statement
4.2. Neonatal Cerebral Ischemia
4.3. Ultrasound Imaging
4.4. Tissue Preparation and Blood-brain Barrier Disruption
4.5. Measurement of Infarct Lesion
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roach, G.D. Perinatal arterial ischemic stroke. Neoreviews 2020, 21, e741–e748. [Google Scholar] [CrossRef]
- Ducrocq, S.; Benjelloun, N.; Plotkine, M.; Ben-Ari, Y.; Charriaut-Marlangue, C. Poly(ADP-ribose) synthase inhibition reduces ischemic injury and inflammation in neonatal rat brain. J. Neurochem. 2000, 74, 2504–2511. [Google Scholar] [CrossRef] [Green Version]
- El Amki, M.; Lerouet, D.; Garraud, M.; Teng, F.; Beray-Berthat, V.; Coqueran, B.; Barsacq, B.; Abbou, C.; Palmier, B.; Marchand-Leroux, C.; et al. Improved reperfusion and vasculoprotection by the Poly(ADP-ribose) polymerase inhibitor PJ34 after stroke and thrombolysis in mice. Mol. Neurobiol. 2018, 55, 9156–9168. [Google Scholar] [CrossRef]
- Soriano, F.G.; Pacher, P.; Mabley, J.; Liaudet, L.; Szabo, C. Rapid reversal of the diabetic endothelial dysfunction by pharmacological inhibition of poly(ADP-ribose) polymerase. Circ. Res. 2001, 89, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Mabley, J.; Soriano, F.G.; Liaudet, L.; Szabo, C. Activation of poly(ADP-ribose) polymerase contributes to the endothelial dysfunction associated with hypertension and aging. Int. J. Mol. Med. 2002, 9, 659–664. [Google Scholar] [CrossRef]
- Yilmaz, B.; Sahin, P.; Ordueri, E.; Celik-Ozenci, C.; Tasatargil, A. Poly(ADP-ribose) polymerase inhibition improves endothelin-1-induced endothelial dysfunction in rat thoracic aorta. Upsala J. Med. Sci. 2014, 119, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, P.; Charriaut-Marlangue, C.; Pansiot, J.; Boutigny, A.; Launay, J.-M.; Besson, V.C. Cerebral vasodilatory property of poly(ADP-ribose) polymerase inhibitor (PJ34) in the neonatal and adult mouse is mediated by the nitric oxide pathway. Int. J. Mol. Sci. 2020, 21, 6569. [Google Scholar] [CrossRef]
- Kratzer, I.; Chip, S.; Vexler, Z.S. Barrier mechanisms in neonatal stroke. Front. Neurosci. 2014, 8, 359. [Google Scholar] [CrossRef] [Green Version]
- Charriaut-Marlangue, C.; Bonnin, P.; Leger, P.-L.; Renolleau, S. Brief updates on hemodynamic responses in animal models of neonatal ischemia and hypoxia-ischemia. Exp. Neurol. 2013, 248, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Csaba, Z.; Vitalis, T.; Charriaut-Marlangue, C.; Margaill, I.; Coqueran, B.; Leger, P.-L.; Parente, I.; Jacquens, A.; Titomanlio, L.; Constans, C.; et al. A simple novel aaproach for detecting blood-brain barrier permeability using GPCR internalization. Neuropathol. Appl. Neurobiol. 2021, 47, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Kettemann, H.; Hanish, U.K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef]
- Nayak, D.; Roth, T.L.; McGavern, D.B. Microglia development and function. ANN Rev. Immunol. 2014, 32, 367–402. [Google Scholar] [CrossRef] [Green Version]
- Moretti, R.; Leger, P.L.; Besson, V.; Csaba, Z.; Pansiot, J.; Di Crisio, L.; Gentili, A.; Titomanlio, L.; Bonnin, P.; Baud, O.; et al. Sildenafil, a cyclic GMP phosphodiesterase inhibitor, induces microglial modulation after focal ischemia in the neonatal mouse brain. J. Neuroinflamm. 2016, 13, 95. [Google Scholar] [CrossRef]
- Charriaut-Marlangue, C.; Leconte, C.; Csaba, Z.; Chafa, L.; Pansiot, J.; Talatizi, M.; Simon, K.; Moretti, R.; Marchand-Leroux, C.; Baud, O.; et al. Sex differences in the effects of PARP inhibition on microglia phenotypes following neonatal ischemia. Brain Behav. Immun. 2018, 73, 375–389. [Google Scholar] [CrossRef]
- Gelot, A.; Villapol, S.; Billette de Villemeur, T.; Renolleau, S.; Charriaut-Marlangue, C. Astrocytic demise in the developing rat and human brain after hypoxic-ischemic damage. Dev. Neurosci. 2009, 31, 459–470. [Google Scholar] [CrossRef]
- Bonnin, P.; Mazighi, M.; Charriaut-Marlangue, C.; Kubis, N. Early collateral recruitment after stroke in infants and adults. Stroke 2019, 50, 2604–2611. [Google Scholar] [CrossRef]
- Nitzsche, A.; Poittevin, M.; Benarab, A.; Bonnin, P.; Faraco, G.; Uchida, H.; Favre, J.; Garcia-Bonilla, L.; Leger, P.L.; Thérond, P.; et al. Endothelial S1P1 signaling counteracts infarct expansion in ischemic stroke. Circ. Res. 2021, 128, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lopez, D.; Faustino, J.; Daneman, R.; Zhou, L.; Lee, S.Y.; Derugin, N.; Wendland, M.F.; Vexler, Z.S. Blood-brain barrier permeability is increased after acute adult stroke but not neonatal stroke in the rat. J. Neurosci. 2012, 32, 9588–9600. [Google Scholar] [CrossRef] [PubMed]
- Ek, C.J.; D’Angelo, B.D.; Baburamani, A.; Lehner, C.; Leverin, A.-L.; Smith, P.; Nilsson, H.; Svedin, P.; Hagberg, H.; Mallard, C. Brain-barrier properties and cerebral blood flow in neonatal mice subjected to cerebral hypoxia-ischemia. J. Cereb. Blood Flow Metab. 2015, 35, 818–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villapol, S.; Bonnin, P.; Fau, S.; Baud, O.; Renolleau, S.; Charriaut-Marlangue, C. Unilateral blood flow decrease induces bilateral and symmetric responses in the immature brain. Am. J. Pathol. 2009, 175, 2111–2120. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.A. The multifaceted profile of activated microglia. Mol. Neurobiol. 2009, 40, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Hamby, A.M.; Suh, S.W.; Kanpinnen, T.M.; Swanson, R.A. Use of poly(ADP-ribose) polymerase inhibitor to suppress inflammation and neuronal death after cerebral ischemia. Stroke 2007, 38, 632–636. [Google Scholar] [CrossRef] [Green Version]
- Benjelloun, N.; Renolleau, S.; Represa, A.; Ben-Ari, Y.; Charriaut-Marlangue, C. Inflammatory responses in the cerebral cortex after ischemia in the P7 neonatal rat. Stroke 1999, 30, 1916–1923; discussion 1923–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Zhang, R.; Huang, Q.Y.; Cao, X.; Zhou, L.Y.; Chu, X.F. Intervention timing and effect of PJ34 on astrocytes during oxygen-glucose deprivation/reperfusion and cell death pathways. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2015, 35, 397–404. [Google Scholar] [CrossRef]
- Phulwani, N.; Kielan, T. Poly(ADP-ribose) polymerases (PARPs) regulate 1-3 astrocyte activation. J. Neurochem. 2008, 106, 578–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, M.; Beray-Berthat, V.; Coqueran, B.; Palmier, B.; Szabo, C.; Plotkine, M.; Margaill, I. Reduction of hemorrhagic transformation by PJ34, a poly(ADP-ribose) polymerase inhibitor, after permanent focal cerebral ischemia. Eur. J. Pharmacol. 2008, 588, 52–57. [Google Scholar] [CrossRef]
- Haddad, M.; Beray-Berthat, V.; Coqueran, B.; Plotkine, M.; Marchand-Leroux, C.; Margaill, I. combined therapy with PJ34, a poly(ADP-ribose) polymerase inhibitor, reduces tissue plasminogen activator-induced hemorrhagic transformations in cerebral ischemia in mice. Fund. Clin. Pharmacol. 2013, 27, 393–401. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnin, P.; Vitalis, T.; Schwendimann, L.; Boutigny, A.; Mohamedi, N.; Besson, V.C.; Charriaut-Marlangue, C. Poly(ADP-Ribose) Polymerase Inhibitor PJ34 Reduces Brain Damage after Stroke in the Neonatal Mouse Brain. Curr. Issues Mol. Biol. 2021, 43, 301-312. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010025
Bonnin P, Vitalis T, Schwendimann L, Boutigny A, Mohamedi N, Besson VC, Charriaut-Marlangue C. Poly(ADP-Ribose) Polymerase Inhibitor PJ34 Reduces Brain Damage after Stroke in the Neonatal Mouse Brain. Current Issues in Molecular Biology. 2021; 43(1):301-312. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010025
Chicago/Turabian StyleBonnin, Philippe, Tania Vitalis, Leslie Schwendimann, Alexandre Boutigny, Nassim Mohamedi, Valérie C Besson, and Christiane Charriaut-Marlangue. 2021. "Poly(ADP-Ribose) Polymerase Inhibitor PJ34 Reduces Brain Damage after Stroke in the Neonatal Mouse Brain" Current Issues in Molecular Biology 43, no. 1: 301-312. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010025