
High Caloric Diet Induces Memory Impairment and Disrupts Synaptic Plasticity in Aged Rats

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
Diet
2.2. Behavior Assessment
2.2.1. Open Field (OF) Test
2.2.2. Novel Object Recognition (NOR) Test
2.3. Electrophysiological Recordings
2.3.1. Acute Hippocampal Slices
2.3.2. Field Excitatory Postsynaptic Potentials (fEPSPs) Quantification and LTP Induction
2.4. Western blot
2.4.1. Protein Extraction
2.4.2. Protein Electrophoresis, Transfer and Detection/Quantification
2.5. Immunohistochemistry
2.5.1. Tissue Processing
2.5.2. Antigen Detection
2.6. Statistical Analysis
3. Results
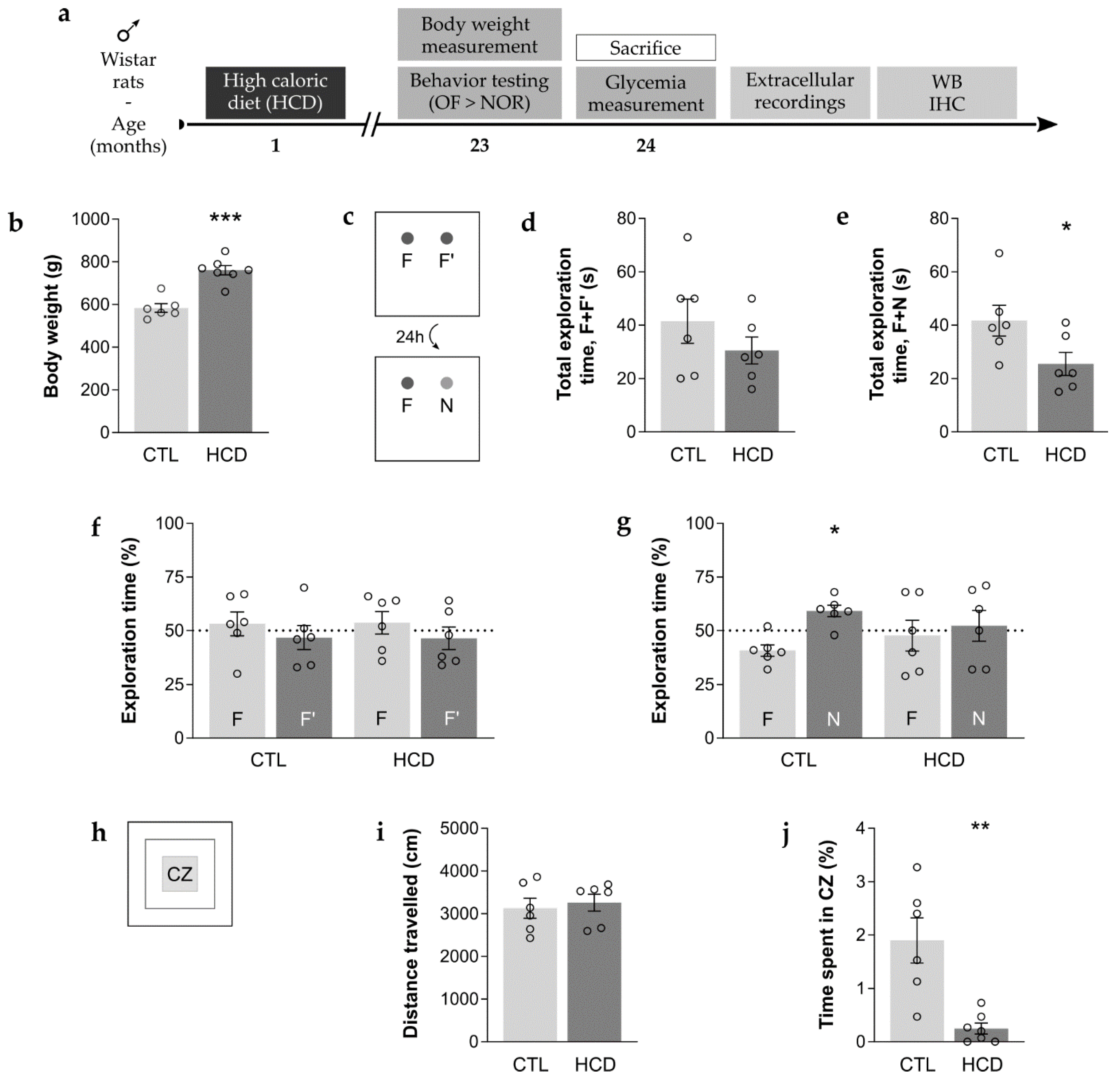
3.1. An Obesogenic HCD Promotes Long-Term Memory Impairment and Enhances Anxious-Related Behavior
3.2. HCD Induces Hippocampal Synaptic Plasticity Impairment along with Reduced Levels of Hippocampal TrkB-FL
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Frasca, D.; Blomberg, B.B.; Paganelli, R. Aging, obesity, and inflammatory age-related diseases. Front. Immunol. 2017, 8, 1745. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Healthy Diet. Available online: https://www.who.int/news-room/fact-sheets/detail/healthy-diet (accessed on 27 July 2021).
- Mujica-Parodi, L.R.; Amgalan, A.; Sultan, S.F.; Antal, B.; Sun, X.; Skiena, S.; Lithen, A.; Adra, N.; Ratai, E.-M.; Weistuch, C.; et al. Diet modulates brain network stability, a biomarker for brain aging, in young adults. Proc. Natl. Acad. Sci. USA 2020, 117, 6170–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beilharz, J.E.; Maniam, J.; Morris, M.J. Diet-induced cognitive deficits: The role of fat and sugar, potential mechanisms and nutritional interventions. Nutrients 2015, 7, 6719–6738. [Google Scholar] [CrossRef] [Green Version]
- Stranahan, A.M.; Norman, E.D.; Lee, K.; Cutler, R.G.; Telljohann, R.S.; Egan, J.M.; Mattson, M.P. Diet-induced insulin resistance impairs hippocampal synaptic plasticity and cognition in middle-aged rats. Hippocampus 2008, 18, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Carrera-Bastos, P.; Fontes-Villaba, M.; O’Keefe, J.; Lindeberg, S. The western diet and lifestyle and diseases of civilization. Res. Rep. Clin. Cardiol. 2011, 2, 15–35. [Google Scholar] [CrossRef] [Green Version]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Monllor, P.; Esteve, D.; Cervera-Ferri, A.; Lloret, M.A. Obesity as a risk factor for Alzheimer’s disease: Implication of leptin and glutamate. Front. Neurosci. 2019, 13, 508. [Google Scholar] [CrossRef]
- Ma, Y.; Ajnakina, O.; Steptoe, A.; Cadar, D. Higher risk of dementia in English older individuals who are overweight or obese. Int. J. Epidemiol. 2020, 49, 1353–1365. [Google Scholar] [CrossRef]
- Singh-Manoux, A.; Dugravot, A.; Shipley, M.; Brunner, E.J.; Elbaz, A.; Sabia, S.; Kivimaki, M. Obesity trajectories and risk of dementia: 28 years of follow-up in the Whitehall II Study. Alzheimers Dement. 2018, 14, 178–186. [Google Scholar] [CrossRef]
- Treviño, S.; Aguilar-Alonso, P.; Flores Hernandez, J.A.; Brambila, E.; Guevara, J.; Flores, G.; Lopez-Lopez, G.; Muñoz-Arenas, G.; Morales-Medina, J.C.; Toxqui, V.; et al. A high calorie diet causes memory loss, metabolic syndrome and oxidative stress into hippocampus and temporal cortex of rats. Synapse 2015, 69, 421–433. [Google Scholar] [CrossRef]
- Yeomans, M.R. Adverse effects of consuming high fat–sugar diets on cognition: Implications for understanding obesity. Proc. Nutr. Soc. 2017, 76, 455–465. [Google Scholar] [CrossRef]
- Binder, D.K.; Scharfman, H.E. Brain-derived neurotrophic factor. Growth Factors 2004, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Li, Y.; Parada, L.F.; Sisodia, S.S. Regulation of hippocampal progenitor cell survival, proliferation and dendritic development by BDNF. Mol. Neurodegener. 2009, 4, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. Neurotrophic Factors 2014, 220, 223–250. [Google Scholar] [CrossRef]
- Barnes, C.A. Long-term potentiation and the ageing brain. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivipelto, M.; Ngandu, T.; Fratiglioni, L.; Viitanen, M.; Kåreholt, I.; Winblad, B.; Helkala, E.L.; Tuomilehto, J.; Soininen, H.; Nissinen, A. Obesity and vascular risk factors at midlife and the risk of dementia and Alzheimer disease. Arch. Neurol. 2005, 62, 1556–1560. [Google Scholar] [CrossRef] [Green Version]
- Kivipelto, M.; Helkala, E.L.; Hänninen, T.; Laakso, M.P.; Hallikainen, M.; Alhainen, K.; Soininen, H.; Tuomilehto, J.; Nissinen, A. Midlife vascular risk factors and late-life mild cognitive impairment: A population-based study. Neurology 2001, 56, 1683–1689. [Google Scholar] [CrossRef]
- Vos, S.J.B.; Van Boxtel, M.P.J.; Schiepers, O.J.G.; Deckers, K.; De Vugt, M.; Carrière, I.; Dartigues, J.F.; Peres, K.; Artero, S.; Ritchie, K.; et al. Modifiable Risk Factors for Prevention of Dementia in Midlife, Late Life and the Oldest-Old: Validation of the LIBRA Index. J. Alzheimers Dis. 2017, 58, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Abbott, K.N.; Arnott, C.K.; Westbrook, R.F.; Tran, D.M.D. The effect of high fat, high sugar, and combined high fat-high sugar diets on spatial learning and memory in rodents: A meta-analysis. Neurosci. Biobehav. Rev. 2019, 107, 399–421. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.M.; Stevenson, R.J. Higher Reported Saturated Fat and Refined Sugar Intake Is Associated With Reduced Hippocampal-Dependent Memory and Sensitivity to Interoceptive Signals. Behav. Neurosci. 2011, 125, 943–955. [Google Scholar] [CrossRef]
- Abrous, D.N.; Wojtowicz, J.M. Interaction between neurogenesis and hippocampal memory system: New vistas. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulose, S.M.; Miller, M.G.; Scott, T.; Shukitt-Hale, B. Nutritional factors affecting adult neurogenesis and cognitive function. Adv. Nutr. 2017, 8, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Barnard, R.J.; Ying, Z.; Roberts, C.K.; Gómez-Pinilla, F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J. Neurochem. 2002, 82, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Mouro, F.M.; Batalha, V.L.; Ferreira, D.G.; Coelho, J.E.; Baqi, Y.; Müller, C.E.; Lopes, L.V.; Ribeiro, J.A.; Sebastião, A.M. Chronic and acute adenosine A2A receptor blockade prevents long-term episodic memory disruption caused by acute cannabinoid CB1 receptor activation. Neuropharmacology 2017, 117, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Taglialatela, G.; Hogan, D.; Zhang, W.-R.; Dineley, K.T. Intermediate- and long-term recognition memory deficits in Tg2576 mice are reversed with acute calcineurin inhibition. Behav. Brain Res. 2009, 200, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, W.W.; Collingridge, G.L. The LTP Program: A data acquisition program for on-line analysis of long-term potentiation and other synaptic events. J. Neurosci. Methods 2001, 108, 71–83. [Google Scholar] [CrossRef]
- Albensi, B.C.; Oliver, D.R.; Toupin, J.; Odero, G. Electrical stimulation protocols for hippocampal synaptic plasticity and neuronal hyper-excitability: Are they effective or relevant? Exp. Neurol. 2007, 204, 1–13. [Google Scholar] [CrossRef]
- Ribeiro, F.F.; Ferreira, F.; Rodrigues, R.S.; Soares, R.; Pedro, D.M.; Duarte-Samartinho, M.; Aroeira, R.I.; Ferreiro, E.; Valero, J.; Solá, S.; et al. Regulation of hippocampal postnatal and adult neurogenesis by adenosine A2A receptor: Interaction with brain-derived neurotrophic factor. Stem Cells 2021, 39, 1362–1381. [Google Scholar] [CrossRef]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.; Aigner, L. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 2005, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aguinis, H.; Gottfredson, R.K.; Joo, H. Best-Practice Recommendations for Defining, Identifying, and Handling Outliers. Organ. Res. Methods 2013, 16, 270–301. [Google Scholar] [CrossRef]
- Martin, M.A.; Roberts, S. Jackknife-after-bootstrap regression influence diagnostics. J. Nonparametr. Stat. 2010, 22, 257–269. [Google Scholar] [CrossRef]
- Gomes, J.R.; Costa, J.T.; Melo, C.V.; Felizzi, F.; Monteiro, P.; Pinto, M.J.; Inácio, A.R.; Wieloch, T.; Almeida, R.D.; Grãos, M.; et al. Excitotoxicity downregulates TrkB.FL signaling and upregulates the neuroprotective truncated TrkB receptors in cultured hippocampal and striatal neurons. J. Neurosci. 2012, 32, 4610–4622. [Google Scholar] [CrossRef] [Green Version]
- Ronan, L.; Alexander-Bloch, A.F.; Wagstyl, K.; Farooqi, S.; Brayne, C.; Tyler, L.K.; Fletcher, P.C. Obesity associated with increased brain age from midlife. Neurobiol. Aging 2016, 47, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Bischof, G.N.; Park, D.C. Obesity and Aging: Consequences for Cognition, Brain Structure, and Brain Function. Psychosom. Med. 2015, 77, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Salvestrini, V.; Sell, C.; Lorenzini, A. Obesity may accelerate the aging process. Front. Endocrinol. 2019, 10, 266. [Google Scholar] [CrossRef]
- Mei, Z.; Grummer-Strawn, L.M.; Pietrobelli, A.; Goulding, A.; Goran, M.I.; Dietz, W.H. Validity of body mass index compared with other body-composition screening indexes for the assessment of body fatness in children and adolescents. Am. J. Clin. Nutr. 2002, 75, 978–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Meireles, M.; Norberto, S.; Leite, J.; Freitas, J.; Pestana, D.; Faria, A.; Calhau, C. High-fat diet-induced obesity Rat model: A comparison between Wistar and Sprague-Dawley Rat. Adipocyte 2016, 5, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelli, E.L.B.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.H.; Cicogna, A.C.; Novelli Filho, J.L.V.B. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Rolls, B.J.; Rowe, E.A.; Turner, R.C. Persistent obesity in rats following a period of consumption of a mixed, high energy diet. J. Physiol. 1980, 298, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Nistiar, F.; Racz, O.; Lukacinova, A.; Hubkova, B.; Novakova, J.; Lovasova, E.; Sedlakova, E. Age dependency on some physiological and biochemical parameters of male Wistar rats in controlled environment. J. Environ. Sci. Health Part A 2012, 47, 1224–1233. [Google Scholar] [CrossRef]
- Dong, W.; Wang, R.; Ma, L.N.; Xu, B.L.; Zhang, J.S.; Zhao, Z.W.; Wang, Y.L.; Zhang, X. Influence of age-related learning and memory capacity of mice: Different effects of a high and low caloric diet. Aging Clin. Exp. Res. 2016, 28, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.J.; D’Angelo, H.; Soch, A.; Watkins, L.R.; Maier, S.F.; Barrientos, R.M. High-fat diet and aging interact to produce neuroinflammation and impair hippocampal- and amygdalar-dependent memory. Neurobiol. Aging 2017, 58, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Castro, J.P.; Andrade, J.P.; Dulce Madeira, M.; Cardoso, A. Cafeteria-diet effects on cognitive functions, anxiety, fear response and neurogenesis in the juvenile rat. Neurobiol. Learn. Mem. 2018, 155, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.D.; Reichelt, A.C. Impaired fear extinction retention and increased anxiety-like behaviours induced by limited daily access to a high-fat/high-sugar diet in male rats: Implications for diet-induced prefrontal cortex dysregulation. Neurobiol. Learn. Mem. 2016, 136, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Sivanathan, S.; Thavartnam, K.; Arif, S.; Elegino, T.; McGowan, P.O. Chronic high fat feeding increases anxiety-like behaviour and reduces transcript abundance of glucocorticoid signalling genes in the hippocampus of female rats. Behav. Brain Res. 2015, 286, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; Tovar, R.; Ramírez-López, M.T.; Navarro, J.A.; Vargas, A.; Suárez, J.; de Fonseca, F.R. Sex-specific anxiety and prefrontal cortex glutamatergic dysregulation are long-term consequences of pre-and postnatal exposure to hypercaloric diet in a rat model. Nutrients 2020, 12, 1829. [Google Scholar] [CrossRef]
- Simon, G.E.; Von Korff, M.; Saunders, K.; Miglioretti, D.L.; Crane, P.K.; Van Belle, G.; Kessler, R.C. Association between obesity and psychiatric disorders in the US adult population. Arch. Gen. Psychiatry 2006, 63, 824–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbaraly, T.N.; Brunner, E.J.; Ferrie, J.E.; Marmot, M.G.; Kivimaki, M.; Singh-Manoux, A. Dietary pattern and depressive symptoms in middle age. Br. J. Psychiatry 2009, 195, 408–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mccarthy, L.H.; Bigal, M.E.; Katz, M.; Derby, C.; Lipton, R.B. Chronic pain and obesity in elderly people: Results from the einstein aging study. J. Am. Geriatr. Soc. 2009, 57, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, G.J.; Pachana, N.A. Anxiety and depression in the elderly: Do we know any more? Curr. Opin. Psychiatry 2010, 23, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.A.; Durazo, E.M.; Slopen, N.; Zaslavsky, A.M.; Buring, J.E.; Silva, T.; Chasman, D.; Williams, D.R. Cumulative psychological stress and cardiovascular disease risk in middle aged and older women: Rationale, design, and baseline characteristics. Am. Heart J. 2017, 192, 1–12. [Google Scholar] [CrossRef]
- Baker, K.D.; Loughman, A.; Spencer, S.J.; Reichelt, A.C. The impact of obesity and hypercaloric diet consumption on anxiety and emotional behavior across the lifespan. Neurosci. Biobehav. Rev. 2017, 83, 173–182. [Google Scholar] [CrossRef]
- Bergado, J.A.; Almaguer, W. Aging and synaptic plasticity: A review. Neural Plast. 2002, 9, 217–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landfield, P.W.; Mcgaugh, J.L.; Tusa, R.J. Theta rhythm: A temporal correlate of memory storage processes in the rat. Science 1972, 175, 87–89. [Google Scholar] [CrossRef]
- Budni, J.; Bellettini-Santos, T.; Mina, F.; Garcez, M.L.; Zugno, A.I. The involvement of BDNF, NGF and GDNF in aging and Alzheimer’s disease. Aging Dis. 2015, 6, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Diógenes, M.; Costenla, A.; Lopes, L.; Jerónimo-Santos, A.; Sousa, V.; Fontinha, B.; Ribeiro, J.; Sebastião, A. Enhancement of LTP in aged rats is dependent on endogenous BDNF. Neuropsychopharmacology 2011, 36, 1823–1836. [Google Scholar] [CrossRef] [Green Version]
- Baranowski, B.J.; MacPherson, R.E.K. Acute exercise induced BDNF-TrkB signalling is intact in the prefrontal cortex of obese, glucose-intolerant male mice. Appl. Physiol. Nutr. Metab. 2018, 43, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Thirumangalakudi, L.; Prakasam, A.; Zhang, R.; Bimonte-Nelson, H.; Sambamurti, K.; Kindy, M.S.; Bhat, N.R. High cholesterol-induced neuroinflammation and amyloid precursor protein processing correlate with loss of working memory in mice. J. Neurochem. 2008, 106, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maesako, M.; Uemura, K.; Iwata, A.; Kubota, M.; Watanabe, K.; Uemura, M.; Noda, Y.; Asada-Utsugi, M.; Kihara, T.; Takahashi, R.; et al. Continuation of Exercise Is Necessary to Inhibit High Fat Diet-Induced β-Amyloid Deposition and Memory Deficit in Amyloid Precursor Protein Transgenic Mice. PLoS ONE 2013, 8, e72796. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Patil, I.Y.; Jiang, T.; Sancheti, H.; Walsh, J.P.; Stiles, B.L.; Yin, F.; Cadenas, E. High-fat diet induces hepatic insulin resistance and impairment of synaptic plasticity. PLoS ONE 2015, 10, e0128274. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.; Keralapurath, M.M.; Lin, Z.; Wagner, J.J.; de La Serre, C.B.; Harn, D.A.; Filipov, N.M. Neurochemical and electrophysiological deficits in the ventral hippocampus and selective behavioral alterations caused by high-fat diet in female C57BL/6 mice. Neuroscience 2015, 297, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Pinho, J.; Vale, R.; Batalha, V.L.; Costenla, A.R.; Dias, R.; Rombo, D.; Sebastião, A.M.; de Mendonça, A.; Diógenes, M.J. Enhanced LTP in aged rats: Detrimental or compensatory? Neuropharmacology 2017, 114, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Fontinha, B.M.; Diógenes, M.J.; Ribeiro, J.A.; Sebastião, A.M. Enhancement of long-term potentiation by brain-derived neurotrophic factor requires adenosine A2A receptor activation by endogenous adenosine. Neuropharmacology 2008, 54, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Minichiello, L. TrkB signalling pathways in LTP and learning. Nat. Rev. Neurosci. 2009, 10, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Silhol, M.; Bonnichon, V.; Rage, F.; Tapia-Arancibia, L. Age-related changes in brain-derived neurotrophic factor and tyrosine kinase receptor isoforms in the hippocampus and hypothalamus in male rats. Neuroscience 2005, 132, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Rage, F.; Silhol, M.; Binamé, F.; Arancibia, S.; Tapia-Arancibia, L. Effect of aging on the expression of BDNF and TrkB isoforms in rat pituitary. Neurobiol. Aging 2007, 28, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.Y.; Kinney, C.E.; An, J.J.; Xu, B. TrkB-expressing neurons in the dorsomedial hypothalamus are necessary and sufficient to suppress homeostatic feeding. Proc. Natl. Acad. Sci. USA 2019, 116, 3256–3261. [Google Scholar] [CrossRef] [Green Version]
- Tsao, D.; Thomsen, H.K.; Chou, J.; Stratton, J.; Hagen, M.; Loo, C.; Garcia, C.; Sloane, D.L.; Rosenthal, A.; Lin, J.C. TrkB agonists ameliorate obesity and associated metabolic conditions in mice. Endocrinology 2008, 149, 1038–1048. [Google Scholar] [CrossRef] [Green Version]
- Perreault, M.; Feng, G.; Will, S.; Gareski, T.; Kubasiak, D.; Marquette, K.; Vugmeyster, Y.; Unger, T.J.; Jones, J.; Qadri, A.; et al. Activation of TrkB with TAM-163 Results in Opposite Effects on Body Weight in Rodents and Non-Human Primates. PLoS ONE 2013, 8, e62616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanqueiro, S.R.; Ramalho, R.M.; Rodrigues, T.M.; Lopes, L.V.; Sebastião, A.M.; Diógenes, M.J. Inhibition of NMDA receptors prevents the loss of BDNF function induced by amyloid β. Front. Pharmacol. 2018, 9, 237. [Google Scholar] [CrossRef]
- Jerónimo-Santos, A.; Vaz, S.H.; Parreira, S.; Rapaz-Lérias, S.; Caetano, A.P.; Scherrer, V.B.; Castrén, E.; Valente, C.A.; Blum, D.; Sebastião, A.M.; et al. Dysregulation of TrkB Receptors and BDNF function by amyloid-β peptide is mediated by calpain. Cereb. Cortex 2015, 25, 3107–3121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca-Gomes, J.; Jerónimo-Santos, A.; Lesnikova, A.; Casarotto, P.; Castrén, E.; Sebastião, A.M.; Diógenes, M.J. TrkB-ICD fragment, originating from BDNF receptor cleavage, is translocated to cell nucleus and phosphorylates nuclear and axonal proteins. Front. Mol. Neurosci. 2019, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.; Moon, H.Y.; Van Praag, H. On the run for hippocampal plasticity. Cold Spring Harb. Perspect. Med. 2018, 8, a029736. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Narisawa, Y.; Shimono, R.; Ohmori, H.; Mori, M.; Ohe, K.; Mine, K.; Enjoji, M. A high fat diet-induced decrease in hippocampal newly-born neurons of male mice is exacerbated by mild psychological stress using a Communication Box. J. Affect. Disord. 2017, 209, 209–216. [Google Scholar] [CrossRef]
- Rivera, P.; Romero-Zerbo, Y.; Pavón, F.J.; Serrano, A.; López-Ávalos, M.D.; Cifuentes, M.; Grondona, J.M.; Bermúdez-Silva, F.J.; Fernández-Llebrez, P.; de Fonseca, F.R.; et al. Obesity-dependent cannabinoid modulation of proliferation in adult neurogenic regions. Eur. J. Neurosci. 2011, 33, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Robison, L.S.; Albert, N.M.; Camargo, L.A.; Anderson, B.M.; Salinero, A.E.; Riccio, D.A.; Abi-Ghanem, C.; Gannon, O.J.; Zuloaga, K.L. High-fat diet-induced obesity causes sex-specific deficits in adult hippocampal neurogenesis in mice. eNeuro 2020, 7, 391–410. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.K.; White, C.W.; Villeda, S.A. The systemic environment: At the interface of aging and adult neurogenesis. Cell Tissue Res. 2018, 371, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Van Praag, H.; Shubert, T.; Zhao, C.; Gage, F.H. Exercise enhances learning and hippocampal neurogenesis in aged mice. J. Neurosci. 2005, 25, 8680–8685. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antigen | Host | Supplier | Catalog Number | Dilution |
|---|---|---|---|---|
| Primary antibodies | ||||
| β-Actin (C4) | Mouse | Santa Cruz Biotechnology, Dallas, TX, USA | sc-47778 | 1:5000 |
| DCX (C-18) | Goat | sc-8066 | 1:500 | |
| GAPDH (6C5) | Mouse | Thermo Fisher Scientific, Waltham, MA, USA | AM4300 | 1:5000 |
| Trk (C-14) | Rabbit | Santa Cruz Biotechnology, Dallas, TX, USA | sc-11 | 1:1000 |
| Secondary antibodies | ||||
| Anti-Goat Alexa Fluor® 488 | Donkey | Thermo Fisher Scientific, Waltham, MA, USA | A-11055 | 1:500 |
| Anti-Mouse IgG-HRP | Goat | Santa Cruz Biotechnology, Dallas, TX, USA | sc-2005 | 1:10,000 |
| Anti-Rabbit IgG-HRP | Bio-Rad, Hercules, CA, USA | 1706515 | 1:10,000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulo, S.L.; Miranda-Lourenço, C.; Belo, R.F.; Rodrigues, R.S.; Fonseca-Gomes, J.; Tanqueiro, S.R.; Geraldes, V.; Rocha, I.; Sebastião, A.M.; Xapelli, S.; et al. High Caloric Diet Induces Memory Impairment and Disrupts Synaptic Plasticity in Aged Rats. Curr. Issues Mol. Biol. 2021, 43, 2305-2319. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030162
Paulo SL, Miranda-Lourenço C, Belo RF, Rodrigues RS, Fonseca-Gomes J, Tanqueiro SR, Geraldes V, Rocha I, Sebastião AM, Xapelli S, et al. High Caloric Diet Induces Memory Impairment and Disrupts Synaptic Plasticity in Aged Rats. Current Issues in Molecular Biology. 2021; 43(3):2305-2319. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030162
Chicago/Turabian StylePaulo, Sara L., Catarina Miranda-Lourenço, Rita F. Belo, Rui S. Rodrigues, João Fonseca-Gomes, Sara R. Tanqueiro, Vera Geraldes, Isabel Rocha, Ana M. Sebastião, Sara Xapelli, and et al. 2021. "High Caloric Diet Induces Memory Impairment and Disrupts Synaptic Plasticity in Aged Rats" Current Issues in Molecular Biology 43, no. 3: 2305-2319. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030162