
Essential Oil of Carvone Chemotype Lippia alba (Verbenaceae) Regulates Lipid Mobilization and Adipogenesis in Adipocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization
2.1.1. Sample Preparation
2.1.2. GC/MS Analysis
- LRI = Linear retention index for the compound of interest.
- n, N = Number of carbon atoms of the n-alkane that elutes before (n), or after (N) the compound of interest (x).
- tRX = Retention time of the compound of interest (min).
- tRN, tRn = Retention times of the n-alkanes that elute before (n) or after (N) the compound of interest (x) (min).
2.2. Cell Culture
2.3. High Glucose—High Insulin Induction Model (HGHI)
2.4. Evaluation of Cytotoxicity by MTT
2.5. Cell Size
2.6. Lipogenesis (Lipid Synthesis) and Lipolysis (Lipid Breakdown)
2.7. Adipogenesis
2.8. Statistical Analysis
3. Results
3.1. Characterization of the Essential Oil of Carvone Chemotype L. alba EO
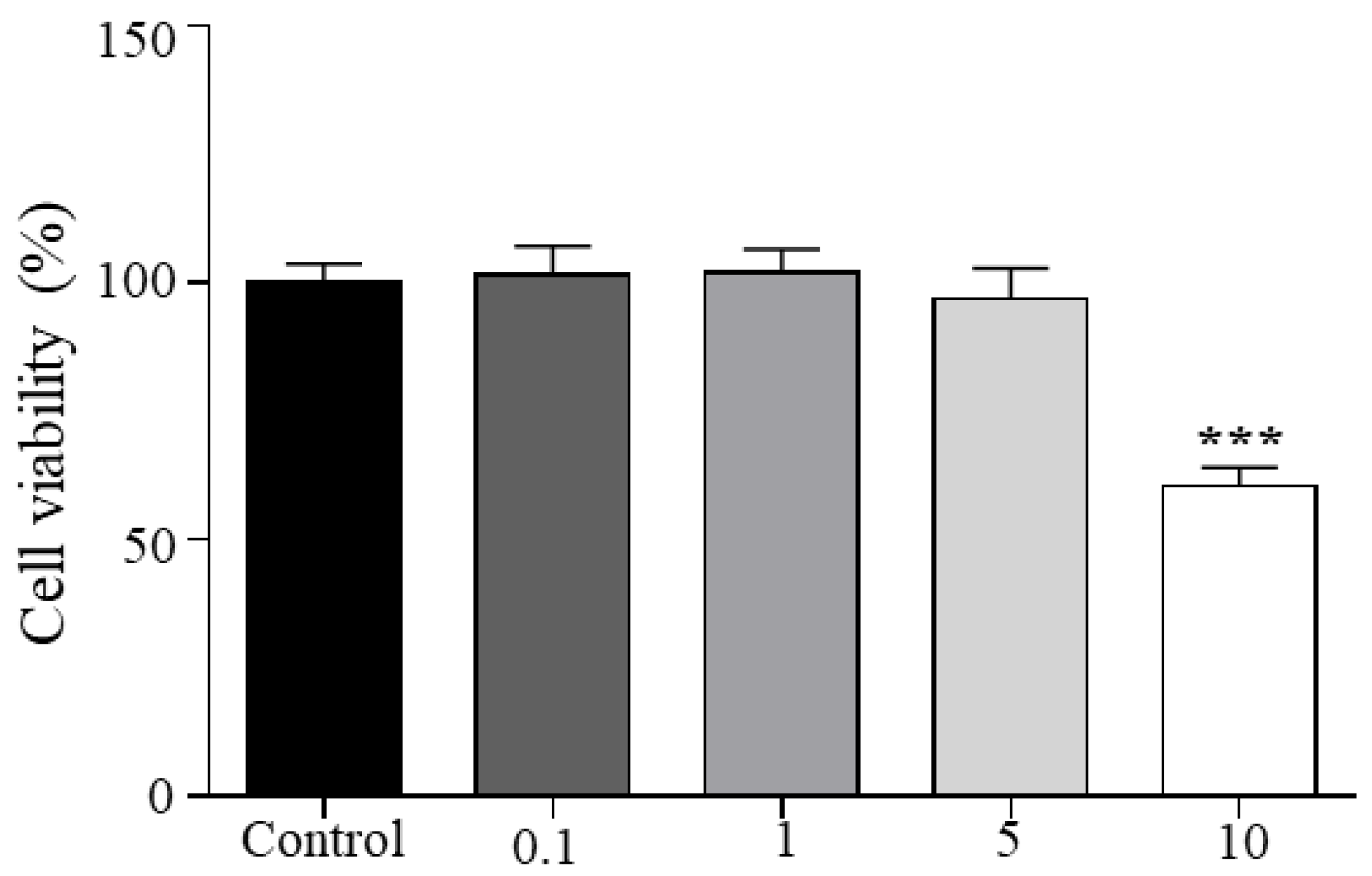
3.2. Viability
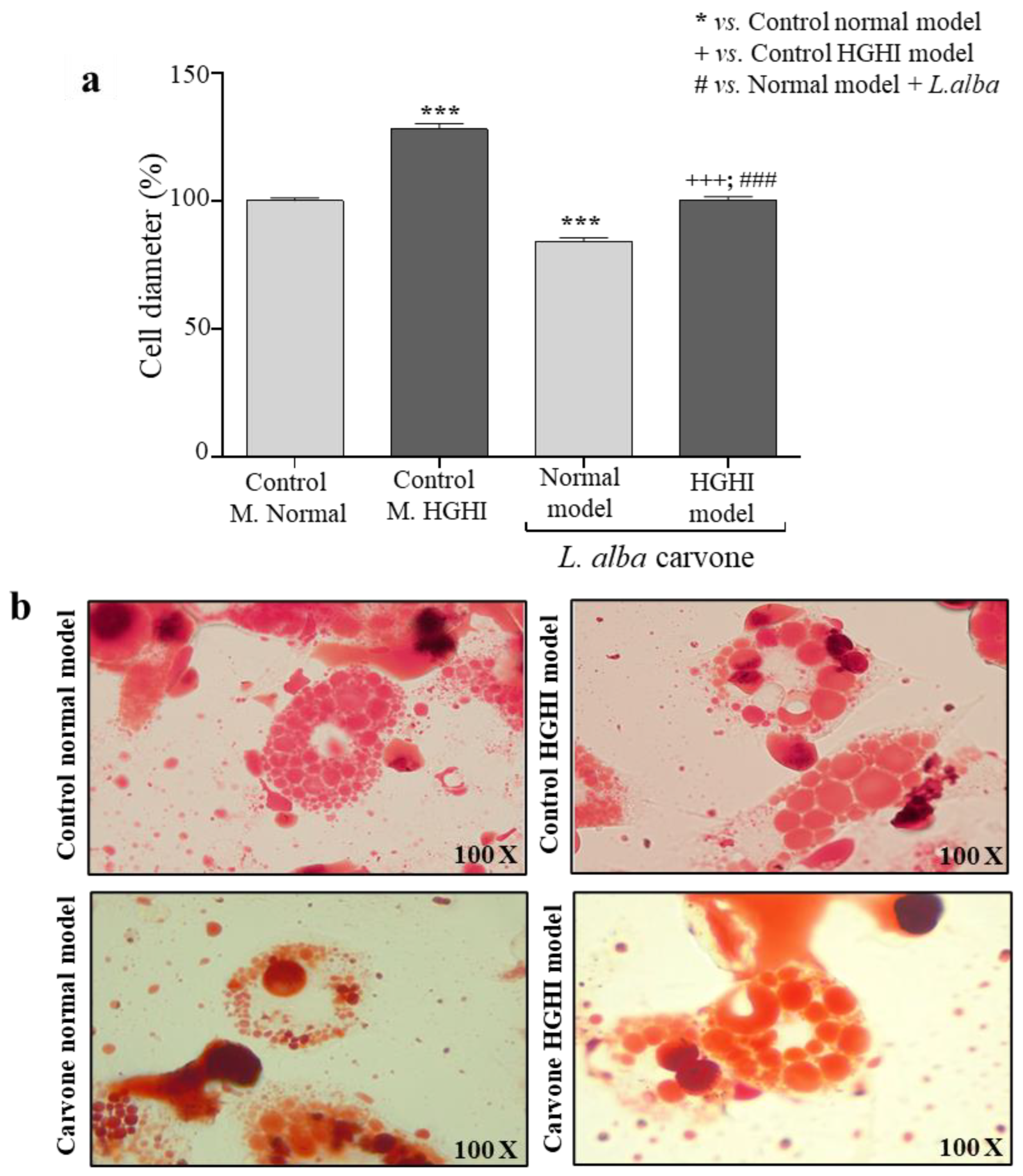
3.3. Cell Size
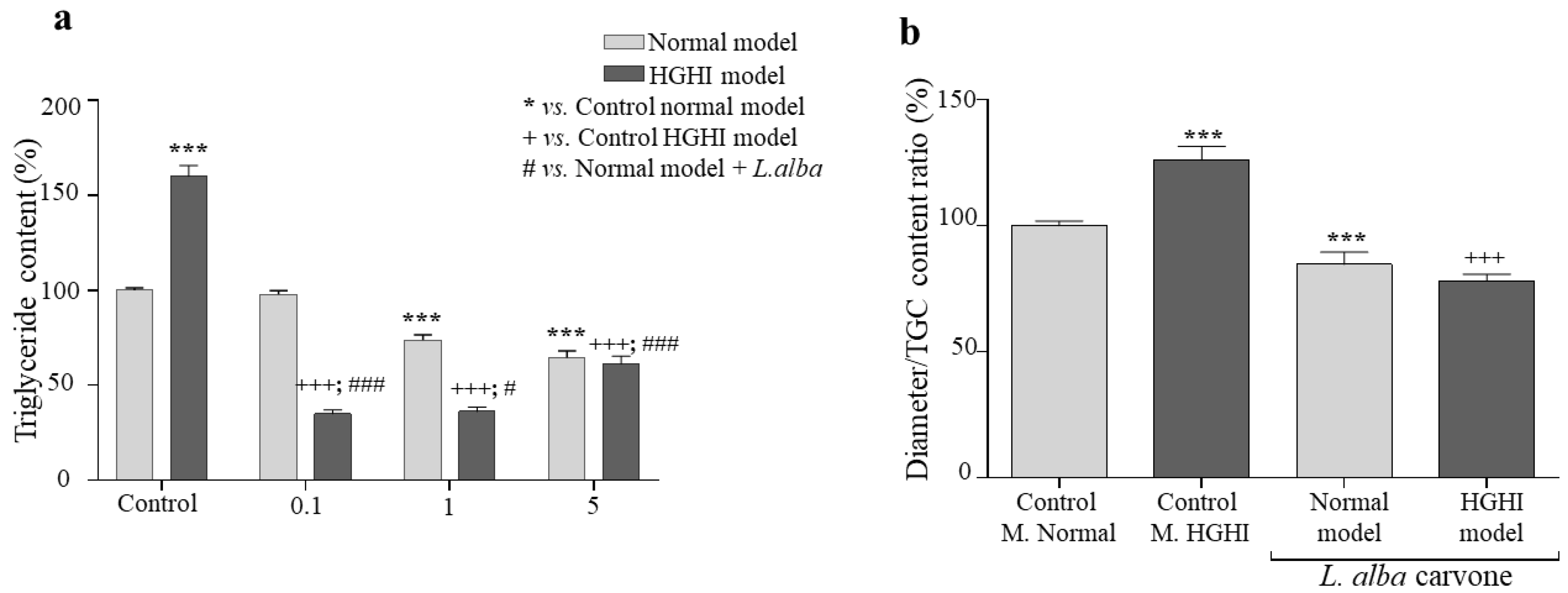
3.4. Lipogenesis
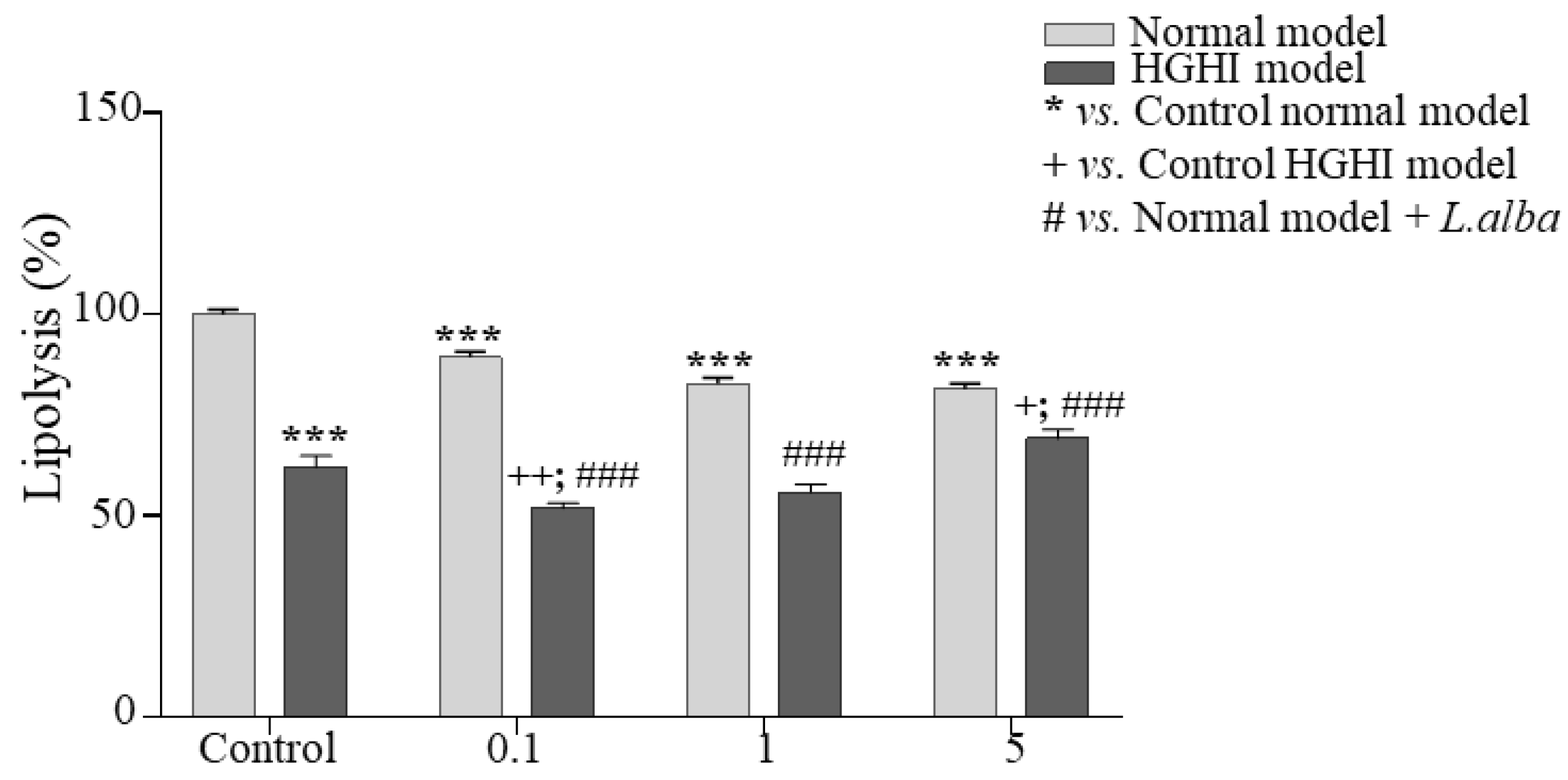
3.5. Lipolysis
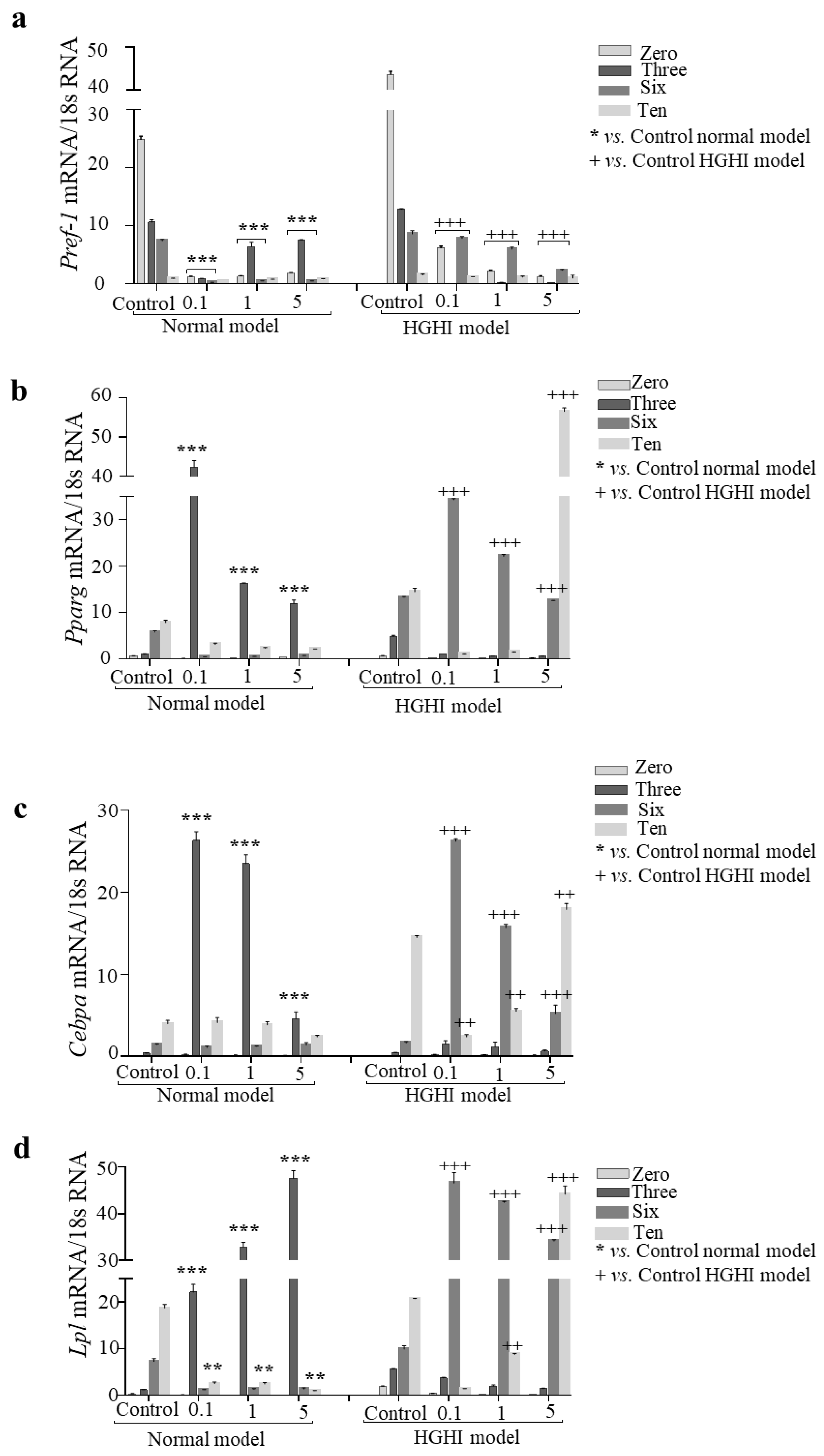
3.6. Adipogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Maria, M.M.; Alberto, D.R.; Rocio, G.R.; Yolanda, J.G.; Natalia, R.M.; Socorro, G.N.; Vázquez-Martínez, R.; Peinado, J.R. Adipobiology for Novel Therapeutic Approaches in Metabolic Syndrome. Curr. Vasc. Pharmacol. 2013, 11, 954–967. [Google Scholar]
- Beg, M.; Chauhan, P.; Varshney, S.; Shankar, K.; Rajan, S.; Saini, D.; Srivastava, M.N.; Yadav, P.P.; Gaikwad, A.N. A withanolide coagulin-L inhibits adipogenesis modulating Wnt/beta-catenin pathway and cell cycle in mitotic clonal expansion. Phytomedicine 2014, 21, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Jack, B.U.; Malherbe, C.J.; Mamushi, M.; Muller, C.J.; Joubert, E.; Louw, J.; Pheiffer, C. Adipose tissue as a possible therapeutic target for polyphenols: A case for Cyclopia extracts as anti-obesity nutraceuticals. Biomed. Pharmacother. 2019, 120, 109439. [Google Scholar] [CrossRef] [PubMed]
- Bolsoni-Lopes, A.; Alonso-Vale, M.I.C. Lipolysis and lipases in white adipose tissue—An update. Arch. Endocrinol. Metab. 2015, 59, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Z.; Ngandiri, D.A.; Llerins Perez, M.; Wolf, A.; Wang, Y. The Molecular Brakes of Adipose Tissue Lipolysis. Front. Physiol. 2022, 13, 826314. [Google Scholar] [CrossRef]
- Daval, M.; Foufelle, F.; Ferré, P. Functions of AMP-activated protein kinase in adipose tissue. J. Physiol. 2006, 574 Pt 1, 55–62. [Google Scholar] [CrossRef]
- Gaidhu, M.P.; Ceddia, R.B. The Role of Adenosine Monophosphate Kinase in Remodeling White Adipose Tissue Metabolism. Exerc. Sport Sci. Rev. 2011, 39, 102–108. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Pêgo-Fernandes, P.M.; Bibas, B.J.; Deboni, M. Obesity: The greatest epidemic of the 21st century? Sao Paulo Med. J. 2011, 129, 283–284. [Google Scholar] [CrossRef] [Green Version]
- Aranaz, P.; Romo-Hualde, A.; Navarro-Herrera, D.; Zabala, M.; López-Yoldi, M.; González-Ferrero, C.; Gil, A.G.; Martínez, J.A.; Vizmanos, J.L.; Milagro, F.I.; et al. Low doses of cocoa extract supplementation ameliorate diet-induced obesity and insulin resistance in rats. Food Funct. 2019, 10, 4811–4822. [Google Scholar] [CrossRef]
- Hussain, K.; Abbas, R.; Abbas, A.; Samiullah, K.; Ahmed, T.; Siddique, F.; Mohsin, M.; Rehman, A.; Rahman, A.U.L.; Waqas, M.U. Anticoccidial potential of Ageratum conyzoides and its effect on Blood parameters of experimentally infected Broiler Chickens. J. Hell. Vet. Med. Soc. 2021, 72, 3085–3090. [Google Scholar] [CrossRef]
- Jamil, M.; Aleem, M.T.; Shaukat, A.; Khan, A.; Mohsin, M.; Rehman, T.U.; Abbas, R.Z.; Saleemi, M.K.; Khatoon, A.; Babar, W.; et al. Medicinal Plants as an Alternative to Control Poultry Parasitic Diseases. Life 2022, 12, 449. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Han, J.H.; Hong, M.; Lee, S.Y.; Lee, S.U.; Kwon, T.H. Antioxidant and lipid-reducing effects of Rosa rugosa root extract in 3T3-L1 cell. Food Sci. Biotechnol. 2022, 31, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Cho, B.O.; Jang, S.I. Anti-obesity effects of Diospyros lotus leaf extract in mice with high-fat diet-induced obesity. Int. J. Mol. Med. 2019, 43, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Phytochemicals and regulation of the adipocyte life cycle. J. Nutr. Biochem. 2008, 19, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Herranz-Lopez, M.; Fernández-Arroyo, S.; Pérez-Sanchez, A.; Barrajón-Catalán, E.; Beltrán-Debón, R.; Menendez, J.; Alonso-Villaverde, C.; Segura-Carretero, A.; Joven, J.; Micol, V. Synergism of plant-derived polyphenols in adipogenesis: Perspectives and implications. Phytomedicine 2012, 19, 253–261. [Google Scholar] [CrossRef]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): A review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Silva-Vazquez, R.; Duran-Melendez, L.A.; Estrada, E.S.; Xie, M.; Dunford, T.; Goad, C. Antioxidant Activity of Essential Oils from Various Mexican Oregano Ecotypes and Oil Fractions Obtained by Distillation. 2017. Available online: https://www.jscimedcentral.com/Chemistry/chemistry-5-1046.pdf (accessed on 3 August 2022).
- Gómez-Sequeda, N.; Cáceres, M.; Stashenko, E.E.; Hidalgo, W.; Ortiz, C. Antimicrobial and Antibiofilm Activities of Essential Oils against Escherichia coli O157:H7 and Methicillin-Resistant Staphylococcus aureus (MRSA). Antibiotics 2020, 9, 730. [Google Scholar] [CrossRef]
- Alsanea, S.; Liu, D. BITC and S-Carvone Restrain High-fat Diet-induced Obesity and Ameliorate Hepatic Steatosis and Insulin Resistance. Pharm. Res. 2017, 34, 2241–2249. [Google Scholar] [CrossRef]
- Lone, J.; Yun, J.W. Monoterpene limonene induces brown fat-like phenotype in 3T3-L1 white adipocytes. Life Sci. 2016, 153, 198–206. [Google Scholar] [CrossRef]
- Soundharrajan, I.; Kim, D.H.; Srisesharam, S.; Kuppusamy, P.; Choi, K.C. R-Limonene Enhances Differentiation and 2-Deoxy-D-Glucose Uptake in 3T3-L1 Preadipocytes by Activating the Akt Signaling Pathway. Evid. Based Complement Altern. Med. 2018, 2018, 4573254. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Huang, Y.-W.; Hou, C.-Y.; Chen, Y.-T.; Dong, C.-D.; Chen, C.-W.; Singhania, R.-R.; Leang, J.-Y.; Hsieh, S.-L. The Anti-Obesity Effects of Lemon Fermented Products in 3T3-L1 Preadipocytes and in a Rat Model with High-Calorie Diet-Induced Obesity. Nutrients 2021, 13, 2809. [Google Scholar] [CrossRef] [PubMed]
- Ngamdokmai, N.; Paracha, T.U.; Waranuch, N.; Chootip, K.; Wisuitiprot, W.; Suphrom, N.; Insumrong, K.; Ingkaninan, K. Effects of Essential Oils and Some Constituents from Ingredients of Anti-Cellulite Herbal Compress on 3T3-L1 Adipocytes and Rat Aortae. Pharmaceuticals 2021, 14, 253. [Google Scholar] [CrossRef]
- Hengpratom, T.; Lowe, G.M.; Thumanu, K.; Suknasang, S.; Tiamyom, K.; Eumkeb, G. Oroxylum indicum (L.) Kurz extract inhibits adipogenesis and lipase activity in vitro. BMC Complement Altern. Med. 2018, 18, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Yang, A.J.; Ekambaranellore, P.; Huang, K.C.; Lin, W.W. Anti-obesity action of INDUS810, a natural compound from Trigonella foenum-graecum: AMPK-dependent lipolysis effect in adipocytes. Obes. Res. Clin. Pract. 2018, 12, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Gómez, C.I.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Moreno-Jiménez, M.R.; Vázquez-Cabral, B.D.; González-Laredo, R.F. Plants with potential use on obesity and its complications. EXCLI J. 2015, 14, 809–831. [Google Scholar]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- González-Castejón, M.; Rodriguez-Casado, A. Dietary phytochemicals and their potential effects on obesity: A review. Pharmacol. Res. 2011, 64, 438–455. [Google Scholar] [CrossRef]
- Linde, G.A.; Colauto, N.B.; Albertó, E.; Gazim, Z.C. Quimiotipos, Extracción, Composición y Aplicaciones del Aceite Esencial de Lippia alba. Rev. Bras. Plantas Med. 2016, 18, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Howitz, K.T.; Sinclair, D.A. Xenohormesis: Sensing the Chemical Cues of Other Species. Cell 2008, 133, 387–391. [Google Scholar] [CrossRef] [Green Version]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herranz-Lopez, M.; Barrajon-Catalan, E.; Segura-Carretero, A.; Menendez, J.A.; Joven, J.; Micol, V. Lemon verbena (Lippia citriodora) polyphenols alleviate obesity-related disturbances in hypertrophic adipocytes through AMPK-dependent mechanisms. Phytomedicine. 2015, 22, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Moreno–Castellanos, N.R.; Angel–Martin, A.; Mantilla–Mora, G. Effects of oleic (18: 1n-9) and palmitic (16:0) fatty acids on the metabolic state of adipocytes. Salud UIS 2021, 53. Available online: https://revistas.uis.edu.co/index.php/revistasaluduis/article/view/11597 (accessed on 3 August 2022). [CrossRef]
- Bonilla-Carvajal, K.; Ángel-Martín, A.; Moreno-Castellanos, N. Hipertrofia y resistencia a la insulina en un modelo in vitro de obesidad y DMT2 inducida por alta glucosa e insulina. Salud UIS 2022, 54. Available online: https://revistas.uis.edu.co/index.php/revistasaluduis/article/view/12576 (accessed on 3 August 2022). [CrossRef]
- Sánchez-Cardona, Y.; Echeverri-Cuartas, C.E.; López, M.E.L.; Moreno-Castellanos, N. Chitosan/Gelatin/PVA Scaffolds for Beta Pancreatic Cell Culture. Polymers 2021, 13, 2372. [Google Scholar] [CrossRef] [PubMed]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Wheaton, IL, USA, 2007; ISBN 1932633219. Available online: https://www.amazon.com.br/Identification-Essential-Components-Chromatography-Spectrometry/dp/1932633219 (accessed on 19 September 2022).
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for frequently reported compounds of plant essential oils. J. Phys. Chem. Ref. Data 2011, 40, 043101. [Google Scholar] [CrossRef] [Green Version]
- “NIST Database”. Version 2.3, Database, Agilent Format. 2017. Available online: https://www.sisweb.com/software/ms/nist.htm (accessed on 19 September 2022).
- International Organization for Standardization (ISO). Biological Evaluation of Medical Devices. Part 5: Tests for In Vitro Cytotoxicity; ISO: Geneva, Switzerland, 2009. [Google Scholar]
- De Cássia da Silveira e Sá, R.; Andrade, L.N.; De Sousa, D.P. A Review on Anti-Inflammatory Activity of Monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef]
- Jing, L.; Zhang, Y.; Fan, S.; Gu, M.; Guan, Y.; Lu, X.; Huang, C.; Zhou, Z. Preventive and ameliorating effects of citrus D-limonene on dyslipidemia and hyperglycemia in mice with high-fat diet-induced obesity. Eur. J. Pharmacol. 2013, 715, 46–55. [Google Scholar] [CrossRef]
- Gomes, D.S.; Negrão-Corrêa, D.A.; Miranda, G.S.; Rodrigues, J.G.M.; Guedes, T.J.F.L.; Junior, W.D.L.; Filho, J.C.F.D.S.; Nizio, D.A.D.C.; Blank, A.F.; Feitosa, V.L.C.; et al. Lippia alba and Lippia gracilis essential oils affect the viability and oviposition of Schistosoma mansoni. Acta Trop. 2022, 231, 106434. [Google Scholar] [CrossRef]
- Ocazionez, R.E.; Meneses, R.; Torres, F.Á.; Stashenko, E. Virucidal activity of Colombian Lippia essential oils on dengue virus replication in vitro. Memórias Do Inst. Oswaldo Cruz. 2010, 105, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab. Syndr. 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Sul, H.S. Minireview: Pref-1: Role in Adipogenesis and Mesenchymal Cell Fate. Mol. Endocrinol. 2009, 23, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Doumit, M.E.; Hill, R.A. Regulation of Adipogenesis and Key Adipogenic Gene Expression by 1, 25-Dihydroxyvitamin D in 3T3-L1 Cells. PLoS ONE 2015, 10, e0126142. [Google Scholar]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Zhang, Y.; Gong, Z.; Huang, C.; Zang, Y.Q. Improved Insulin Resistance and Lipid Metabolism by Cinnamon Extract through Activation of Peroxisome Proliferator-Activated Receptors. PPAR Res. 2008, 2008, e581348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zechner, R.; Strauss, J.; Frank, S.; Wagner, E.; Hofmann, W.; Kratky, D.; Hiden, M. The role of lipoprotein lipase in adipose tissue development and metabolism. Int. J. Obes. 2000, 24, S53–S56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Linear Rentention Indices | GC Relative Area, % DB-5MS | Identifi-Cation Criteria | |||
|---|---|---|---|---|---|---|---|
| DB-5MS | DB-WAX | ||||||
| Exp.* | Lit.** | Exp.* | Lit.** | ||||
| 1 | cis-Hex-3-en-1-ol | 854 | 850 [37] | 1386 | 1380 [38] | 0.3 | a, b |
| 2 | α-Pinene | 934 | 932 [37] | 1022 | 1025 [38] | 0.1 | a, b, c |
| 3 | Camphene | 952 | 954 [37] | 1066 | 1069 [38] | 0.2 | a, b, c |
| 4 | Oct-1-en-3-ol | 979 | 980 [38] | 1451 | 1444 [38] | 0.3 | a, b, c |
| 5 | β-Myrcene | 989 | 990 [37] | 1163 | 1161 [38] | 0.7 | a, b, c |
| 6 | Limonene | 1037 | 1029 [37] | 1206 | 1198 [38] | 31.5 | a, b, c |
| 7 | trans-β-Ocimene | 1047 | 1050 [37] | 1252 | 1250 [38] | 0.6 | a, b |
| 8 | Linalool | 1100 | 1096 [37] | 1548 | 1543 [38] | 1.0 | a, b, c |
| 9 | trans-p-Mentha-2,8-dien-1-ol | 1125 | 1122 [37] | 1632 | 1639 [38] | 0.3 | a, b |
| 10 | cis-Limonene oxide | 1140 | 1136 [37] | 1465 | 1451 [38] | 0.3 | a, b, c |
| 11 | Borneol | 1178 | 1169 [37] | 1708 | 1700 [38] | 0.8 | a, b |
| 12 | trans-Dihydrocarvone | 1209 | 1200 [37] | - | - | 0.1 | a, b |
| 13 | cis-Carveol | 1229 | 1229 [37] | 1838 | 1854 [38] | 0.4 | a, b |
| 14 | Carvone | 1259 | 1258 [39] | 1747 | 1734 [38] | 37.3 | a, b, c |
| 15 | Piperitone oxide | 1262 | 1256 [39] | 1725 | 1711 [39] | 0.5 | a, b |
| 16 | Piperitone | 1264 | 1264 [39] | 1737 | 1730 [38] | 1.8 | a, b |
| 17 | Geranial | 1269 | 1270 [38] | 1747 | 1725 [38] | 0.4 | a, b, c |
| 18 | Carvone oxide | 1281 | 1273 [37] | 1840 | - | 0.1 | a, b |
| 19 | Thymol | 1290 | 1290 [37] | 2180 | 2164 [38] | 0.5 | a, b, c |
| 20 | trans-Carvyl acetate | 1331 | 1342 [37] | - | 1727 [38] | 0.1 | a, b |
| 21 | Piperitenone | 1343 | 1343 [37] | 1929 | 1909 [38] | 1.7 | a, b |
| 22 | α-Copaene | 1382 | 1376 [37] | 1501 | 1491 [38] | 0.1 | a, b |
| 23 | β-Bourbonene | 1392 | 1376 [37] | 1530 | 1523 [38] | 2.6 | a, b |
| 24 | β-Elemene | 1394 | 1390 [37] | 1598 | 1591 [38] | 1.6 | a, b |
| 25 | β-Ylangene | 1427 | 1421 [39] | 1585 | 1589 [39] | 0.5 | a, b |
| 26 | trans-β-Caryophyllene | 1430 | 1427 [39] | 1609 | 1599 [38] | 0.4 | a, b, c |
| 27 | β-Copaene | 1439 | 1432 [39] | 1585 | 1580 [38] | 0.5 | a, b |
| 28 | trans-β-Farnesene | 1454 | 1456 [37] | 1669 | 1664 [38] | 1.4 | a, b |
| 29 | Alloaromadendrene | 1471 | 1460 [37] | 1658 | 1650 [39] | 0.7 | a, b |
| 30 | Germacrene D | 1493 | 1481 [38] | 1725 | 1708 [38] | 11.8 | a, b, c |
| 31 | Bicyclogermacrene | 1505 | 1500 [37] | 1748 | 1735 [38] | 0.7 | a, b |
| 32 | δ-Cadinene | 1524 | 1523 [37] | 1762 | 1756 [38] | 0.4 | a, b |
| 33 | trans-α-Bisabolene | 1544 | 1544 [39] | 1775 | 1777 [39] | 0.2 | a, b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonilla-Carvajal, K.; Stashenko, E.E.; Moreno-Castellanos, N. Essential Oil of Carvone Chemotype Lippia alba (Verbenaceae) Regulates Lipid Mobilization and Adipogenesis in Adipocytes. Curr. Issues Mol. Biol. 2022, 44, 5741-5755. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44110389
Bonilla-Carvajal K, Stashenko EE, Moreno-Castellanos N. Essential Oil of Carvone Chemotype Lippia alba (Verbenaceae) Regulates Lipid Mobilization and Adipogenesis in Adipocytes. Current Issues in Molecular Biology. 2022; 44(11):5741-5755. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44110389
Chicago/Turabian StyleBonilla-Carvajal, Katherin, Elena E. Stashenko, and Natalia Moreno-Castellanos. 2022. "Essential Oil of Carvone Chemotype Lippia alba (Verbenaceae) Regulates Lipid Mobilization and Adipogenesis in Adipocytes" Current Issues in Molecular Biology 44, no. 11: 5741-5755. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44110389