How Different Dietary Methionine Sources Could Modulate the Hepatic Metabolism in Rainbow Trout?

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Feeding Trial and Growth Performances
- Mortality (%) (M%) = [(number of dead fish/number of fish at the beginning) × 100];
- Individual weight gain (iWG, g) = iFBW (individual final body weight, g) − iIBW (individual initial body weight, g);
- Feed conversion ratio (FCR) = total feed supplied [g, dry matter (DM)/WG (g);
- Protein efficiency ratio (PER) = WG (g)/total protein fed (g, DM);
- Specific growth rate (SGR, % day-1) = [(lnFBW − lnIBW)/number of feeding days] × 100;
- Feeding rate (FR, %/d) = [(total feed supplied (g, DM) × 100/number of feeding day)]/[e(lnFBW+lnIBW)×0.5];
- Thermal growth coefficient = (FBW1/3 − IBW1/3)/Ʃ(T × d) × 100 (where d, is the number of days of trial).
2.3. Chemical Analyses of Diets and Fish Whole-Body Composition
2.4. Gene Expression Analysis
2.4.1. Total RNA Extraction and cDNA Synthesis
2.4.2. Generation of In Vitro-Transcribed mRNAs for Each Gene Standard Curves
2.4.3. Transcript Quantification by One-Step TaqMan® Real-Time RT-PCR
2.5. SAM/SAH HPLC Analysis
2.5.1. Standards and Sample Processing
2.5.2. HPLC System and Chromatographic Conditions
2.5.3. Validation
2.6. Statistical Analyses
3. Results
3.1. Fish Performance
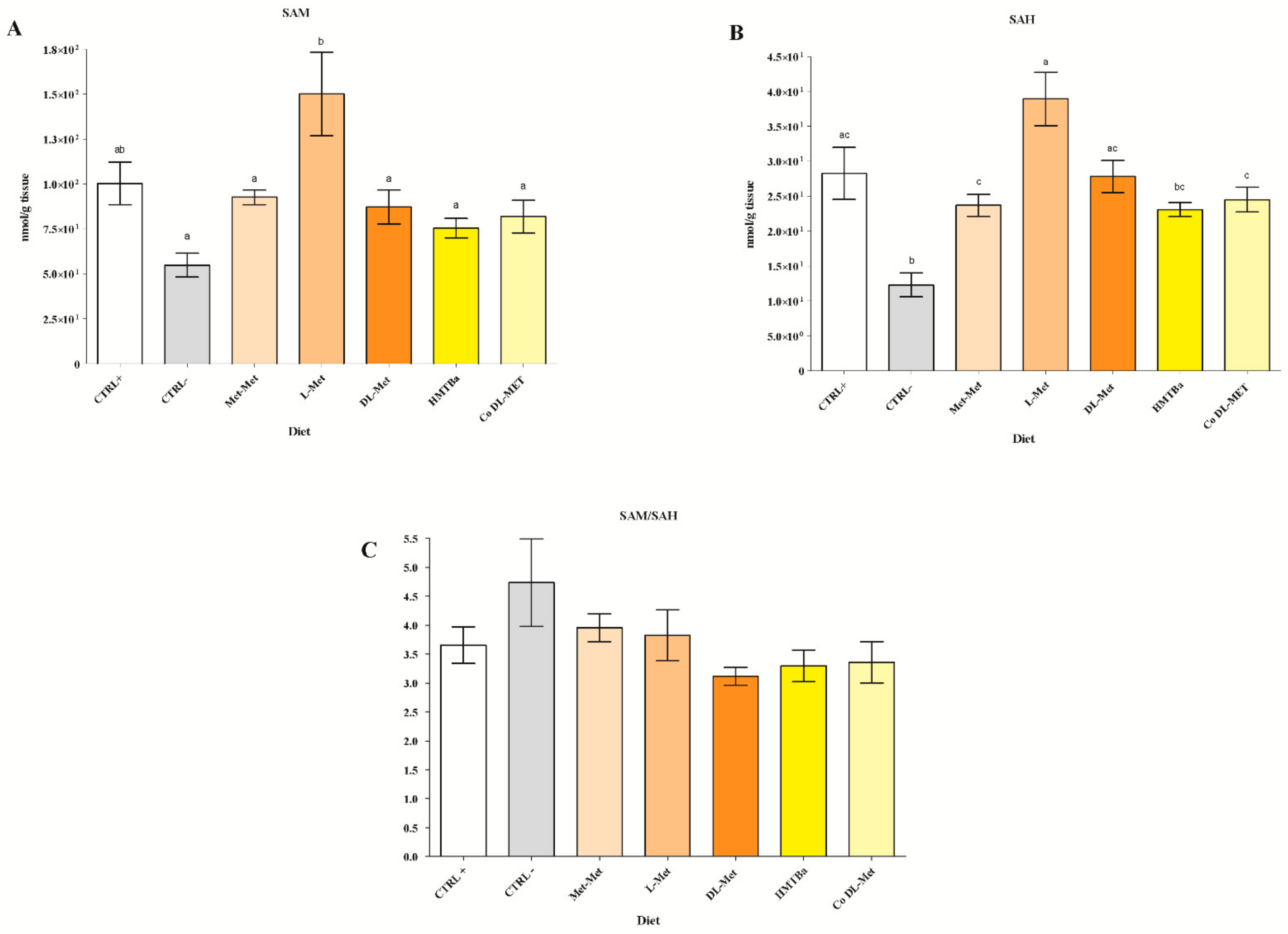
3.2. Hepatic SAM and SAH Levels
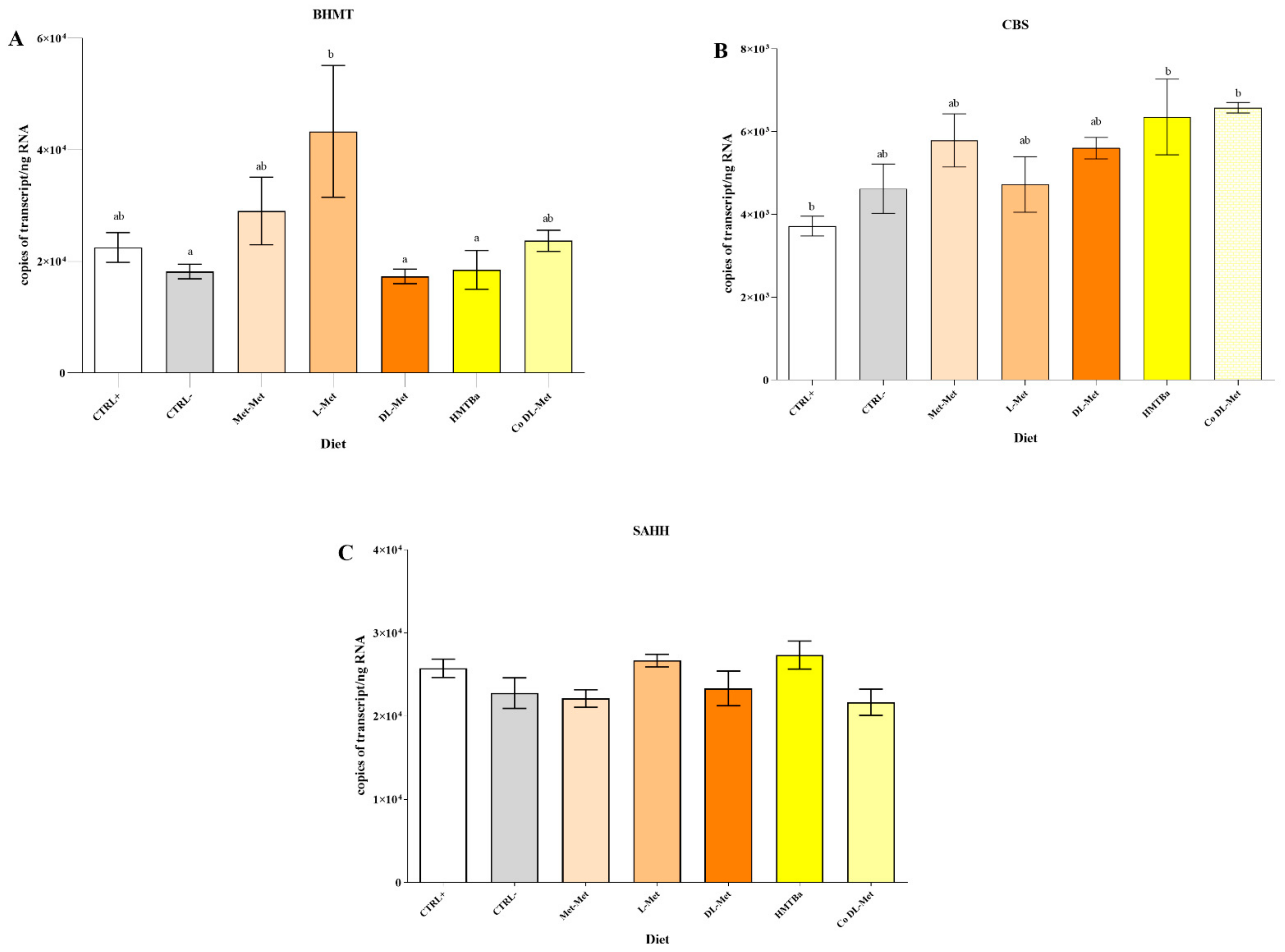
3.3. Expression of Genes Involved into Met Metabolism
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Hardy, R.W. Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Chi, S.; Tan, B.; Dong, X.; Yang, Q.; Liu, H. Effects of supplemental coated or crystalline methionine in low-fishmeal diet on the growth performance and body composition of juvenile cobia Rachycentron canadum (Linnaeus). Chin. J. Oceanol. Limnol. 2014, 32, 1297–1306. [Google Scholar] [CrossRef]
- Rolland, M.; Skov, P.V.; Larsen, B.K.; Holm, J.; Gómez-Requeni, P.; Dalsgaard, J. Increasing levels of dietary crystalline methionine affect plasma methionine profiles, ammonia excretion, and the expression of genes related to the hepatic intermediary metabolism in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 198, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.C.; Gong, S.Y.; Yang, H.J.; Lin, Y.C.; Yu, D.H.; Luo, Z. Effects of supplementation of crystalline or coated lysine and/or methionine on growth performance and feed utilization of the Chinese sucker, Myxocyprinus asiaticus. Aquaculture 2011, 316, 31–36. [Google Scholar] [CrossRef]
- Powell, C.D.; Chowdhury, M.A.K.; Bureau, D.P. Assessing the bioavailability of L-methionine and a methionine hydroxy analogue (MHA-Ca) compared to DL-methionine in rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2017, 48, 332–346. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, W.B.; Bai, N.; Mai, K.S.; Xu, W. Effects of dietary crystalline methionine or oligo-methionine on growth performance and feed utilization of white shrimp (Litopenaeus vannamei) fed plant protein-enriched diets. Aquac. Nutr. 2013, 19, 39–46. [Google Scholar] [CrossRef]
- Guo, T.Y.; Zhao, W.; Liao, S.Y.; Xie, J.J.; Xie, S.W.; Masagounder, K.; Tian, L.X.; Niu, J.; He, J.Y.; Liu, Y.J. Dietary dl-methionyl-dl-methionine supplementation increased growth performance, antioxidant ability, the content of essential amino acids and improved the diversity of intestinal microbiota in Nile tilapia (Oreochromis niloticus). Br. J. Nutr. 2020, 123, 72–83. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, Y.; Liu, W.; Ju, T.; Zhan, X. Effects of different methionine sources on production and reproduction performance, egg quality and serum biochemical indices of broiler breeders. Asian-Australas. J. Anim. Sci. 2017, 30, 828–833. [Google Scholar] [CrossRef]
- Dibner, J.J.; Durley, R.C.; Kostelc, J.G.; Ivey, F.J. 2-Hydroxy-4-(methylthio) butanoic acid is a naturally occurring methionine precursor in the chick. J. Nutr. 1990, 120, 553–560. [Google Scholar] [CrossRef]
- Vázquez-Añón, M.; Bertin, G.; Mercier, Y.; Reznik, G.; Roberton, J.L. Review of the chemistry, metabolism, and dose response of two supplemental methionine sources and the implications in their relative bioefficacy. World’s Poult. Sci. J. 2017, 73, 725–736. [Google Scholar] [CrossRef]
- Zhang, S.; Wong, E.A.; Gilbert, E.R. Bioavailability of different dietary supplemental methionine sources in animals. Front. Biosci. 2015, 7, 478–490. [Google Scholar]
- Teodósio, R.; Engrola, S.; Cabano, M.; Colen, R.; Masagounder, K.; Aragão, C. Metabolic and nutritional responses of Nile tilapia juveniles to dietary methionine sources. Br. J. Nutr. 2022, 127, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Dibner, J.J. Review of the metabolism of 2-hydroxy-4-(methylthio) butanoic acid. World’s Poult. Sci. J. 2003, 59, 99–110. [Google Scholar] [CrossRef]
- To, V.P.H.T.; Subramaniam, M.; Masagounder, K.; Loewen, M.E. Characterization of the segmental transport mechanisms of DL-methionine hydroxy analogue along the intestinal tract of rainbow trout with an additional comparison to DL-methionine. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 249, 110776. [Google Scholar] [CrossRef]
- Zhou, Y.; He, J.; Su, N.; Masagounder, K.; Xu, M.; Chen, L. Effects of DL-methionine and a methionine hydroxy analogue (MHA-Ca) on growth, amino acid profiles and the expression of genes related to taurine and protein synthesis in common carp (Cyprinus carpio). Aquaculture 2021, 532, 735962. [Google Scholar] [CrossRef]
- He, Y.; Chi, S.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S. Effects of three methionine sources in diets on temporal B0AT1 and ASCT2 mRNA expression in different intestinal segments of cobia, Rachycentron canadum. J. World Aquac. Soc. 2020, 52, 171–183. [Google Scholar] [CrossRef]
- Ma, R.; Hou, H.; Mai, K.; Bharadwaj, A.S.; Cao, H.; Ji, F.; Zhang, W. Comparative study on the effects of L-methionine or 2-hydroxy-4-(methylthio) butanoic acid as dietary methionine source on growth performance and anti-oxidative responses of turbot (Psetta maxima). Aquaculture 2013, 412–413, 136–143. [Google Scholar] [CrossRef]
- Zhao, J.X.; Li, X.Q.; Leng, X.J.; Peng, S.; Hu, J.; Zhao, X.X. Comparative study on the utilization of different methionine sources by channel catfish, Ictalurus punctatus (Rafinesque, 1818). Aquac. Res. 2017, 48, 3618–3630. [Google Scholar] [CrossRef]
- Guo, J.; Bao, Y.; Davis, R.; Abebe, A.; Wilson, A.E.; Davis, D.A. Application of meta-analysis towards understanding the effect of adding a methionine hydroxy analogue in the diet on growth performance and feed utilization of fish and shrimp. Rev. Aquac. 2020, 12, 2316–2332. [Google Scholar] [CrossRef]
- Yang, Z.; Htoo, J.K.; Liao, S.F. Methionine nutrition in swine and related monogastric animals: Beyond protein biosynthesis. Anim. Feed Sci. Technol. 2020, 268, 114608. [Google Scholar] [CrossRef]
- Su, Y.; Wu, P.; Feng, L.; Jiang, W.; Jiang, J.; Zhang, Y. Fish and Shell fi sh Immunology The improved growth performance and enhanced immune function by DL-methionyl-DL-methionine are associated with NF-κ B and TOR signalling in intestine of juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2018, 74, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Espe, M.; Andersen, S.M.; Holen, E.; Rønnestad, I.; Veiseth-Kent, E.; Zerrahn, J.E.; Aksnes, A. Methionine deficiency does not increase polyamine turnover through depletion of hepatic S-adenosylmethionine in juvenile Atlantic salmon. Br. J. Nutr. 2014, 112, 1274–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terova, G.; Ceccotti, C.; Ascione, C.; Gasco, L.; Rimoldi, S. Effects of partially defatted hermetia illucens meal in rainbow trout diet on hepatic methionine metabolism. Animals 2020, 10, 1059. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.A. Amino Acid Metabolism; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Espe, M.; Veiseth-Kent, E.; Zerrahn, J.E.; Rønnestad, I.; Aksnes, A. Juvenile Atlantic salmon decrease white trunk muscle IGF-1 expression and reduce muscle and plasma free sulphur amino acids when methionine availability is low while liver sulphur metabolites mostly is unaffected by treatment. Aquac. Nutr. 2016, 22, 801–812. [Google Scholar] [CrossRef]
- Goff, J.B.; Gatlin, D.M. Evaluation of different sulfur amino acid compounds in the diet of red drum, Sciaenops ocellatus, and sparing value of cystine for methionine. Aquaculture 2004, 241, 465–477. [Google Scholar] [CrossRef]
- Boonyoung, S.; Haga, Y.; Satoh, S. Preliminary study on effects of methionine hydroxy analog and taurine supplementation in a soy protein concentrate-based diet on the biological performance and amino acid composition of rainbow trout [Oncorhynchus mykiss (Walbaum)]. Aquac. Res. 2013, 44, 1339–1347. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Is dietary taurine supplementation beneficial for farmed fish and shrimp? A comprehensive review. Rev. Aquac. 2014, 6, 241–255. [Google Scholar] [CrossRef]
- Biasato, I.; Chemello, G.; Caimi, C.; Bellezza Oddon, S.; Capucchio, M.T.; Colombino, E.; Schiavone, A.; Ceccotti, C.; Terova, G.; Gasco, L. Taurine supplementation in plant-based diets for juvenile rainbow trout (Oncorhynchus mykiss): Effects on growth performance, whole body composition, and histomorphological features. Anim. Feed Sci. Technol. 2022, 289, 115314. [Google Scholar] [CrossRef]
- Ceccotti, C.; Al-Sulaivany, B.S.A.; Al-Habbib, O.A.M.; Saroglia, M.; Rimoldi, S.; Terova, G. Protective effect of dietary taurine from ROS production in European seabass under conditions of forced swimming. Animals 2019, 9, 607. [Google Scholar] [CrossRef] [Green Version]
- Rønnestad, I.; Conceição, L.E.C.; Aragão, C.; Dinis, M.T. Free amino acids are absorbed faster and assimilated more efficiently than protein in postlarval Senegal sole (Solea senegalensis). J. Nutr. 2000, 130, 2809–2812. [Google Scholar] [CrossRef] [Green Version]
- Mato, J.M.; Alvarez, L.; Ortiz, P.; Pajares, M.A. S-adenosylmethionine synthesis: Molecular mechanisms and clinical implications. Pharmacol. Ther. 1997, 73, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Kwasek, K.; Terova, G.; Lee, B.J.; Bossi, E.; Saroglia, M.; Dabrowski, K. Dietary methionine supplementation alters the expression of genes involved in methionine metabolism in salmonids. Aquaculture 2014, 433, 223–228. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Alami-Durante, H.; Aragão, C.; Kaushik, S.J.; Geurden, I. Parental and early-feeding effects of dietary methionine in rainbow trout (Oncorhynchus mykiss). Aquaculture 2017, 469, 16–27. [Google Scholar] [CrossRef]
- Zuo, F.; Gu, Q.; Li, S.; Wei, H.; Peng, J. Effects of different methionine sources on methionine metabolism in the IPEC-J2 cells. Biomed. Res. Int. 2019, 2019, 5464906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Venegas, R.; Geraert, P.A.; Ferrer, R. Conversion of the methionine hydroxy analogue DL-2-hydroxy-(4-methylthio) butanoic acid to sulfur-containing amino acids in the chicken small intestine. Poult. Sci. 2006, 85, 1932–1938. [Google Scholar] [CrossRef]
- Rowling, M.J.; McMullen, M.H.; Chipman, D.C.; Schalinske, K.L. Hepatic glycine N-methyltransferase is up-regulated by excess dietary methionine in rats. J. Nutr. 2002, 132, 2545–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espe, M.; Hevrøy, E.M.; Liaset, B.; Lemme, A.; El-Mowafi, A. Methionine intake affect hepatic sulphur metabolism in Atlantic salmon, Salmo salar. Aquaculture 2008, 274, 132–141. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| BAS+ | BAS− | |
|---|---|---|
| Fish meal A a | 0.00 | 13.56 |
| Fish meal B b | 45.00 | 0 |
| Fish protein hydrolysate | 5.00 | 0 |
| Fish oil | 12.94 | 14.30 |
| Rapeseed meal | 6.70 | 0.00 |
| Soybean meal | 0.00 | 19.33 |
| Guar germ meal | 0.00 | 20.00 |
| Wheat milling | 22.52 | 8.00 |
| Corn gluten meal | 0.00 | 8.00 |
| Wheat gluten | 4.92 | 1.00 |
| Soy protein concentrate | 0.00 | 12.00 |
| Emulsifier (E484) | 0.20 | 0.20 |
| MAP c | 1.20 | 1.90 |
| Lysine hydrochloride | 0.30 | 1.40 |
| Vitamin and mineral premix d | 0.65 | 0.65 |
| Stay C 35% | 0.07 | 0.06 |
| Taurine | 0.50 | 0.60 |
| Amino Acids (% as Is) | BAS+ | BAS− |
|---|---|---|
| Ala | 2.68 | 2.08 |
| Arg | 2.50 | 2.93 |
| Asp | 3.76 | 3.72 |
| Glu | 7.45 | 6.79 |
| Gly | 2.75 | 2.30 |
| His | 0.97 | 1.05 |
| Iso | 1.97 | 1.56 |
| Leu | 3.32 | 3.11 |
| Lys | 3.06 | 3.13 |
| Met | 1.00 | 0.70 |
| Phe | 1.78 | 2.07 |
| Pro | 2.37 | 2.21 |
| Ser | 2.01 | 1.85 |
| Thr | 1.79 | 1.56 |
| Tyr | 1.38 | 1.34 |
| Trp | 0.50 | 0.39 |
| Val | 2.35 | 1.81 |
| Tau | 0.66 | 0.65 |
| CTRL+ | CTRL− | Met-Met | L-Met | DL-Met | HMTBa | Co DL-Met | |
|---|---|---|---|---|---|---|---|
| DM | 95.79 | 95.64 | 95.91 | 95.63 | 95.42 | 95.67 | 95.21 |
| CP | 44.43 | 44.19 | 44.23 | 44.83 | 44.23 | 44.73 | 44.41 |
| EE | 21.70 | 21.20 | 21.54 | 21.19 | 21.28 | 21.24 | 21.49 |
| Ash | 7.05 | 7.12 | 7.03 | 7.05 | 7.03 | 7.14 | 7.11 |
| Gene | Nucleotide Sequence (5′→3′) | Purpose |
|---|---|---|
| Cloning | ||
| BHMT FW | TGCAGAGTACTTTGAGCACGT | |
| BHMT RV | CCGTGACTACTGGGAGAAGC | |
| SAHHB FW | CCCTTCAAAGTTGCTGACATCA | |
| SAHHB RV | ATGTGTGGTGCATTGAGCAGA | |
| CBS FW | AAACCCTGGTGGTGGAAC | |
| CBS RV | GTGCTCTACAAACAATTCAAACAGGT | |
| Standard Curve | ||
| T7 BHMT sense | gtaatacgactcactatagggTGAAAGAGGGAGTGGAGAGG | |
| BHMT antisense | CCGTGACTACTGGGAGAAGC | |
| T7 SAHHB sense | gtaatacgactcactatagggAGATGAGGGAGCTGTATGGC | |
| SAHHB antisense | ATGTGTGGTGCATTGAGCAGA | |
| T7 CBS sense | gtaatacgactcactatagggAAACCCTGGTGGTGGAAC | |
| CBS antisense | GTGCTCTACAAACAATTCAAACAGGT | |
| Real-time RT-PCR | ||
| BHMT FW | TGCCAGGGATTCATCGATCTG | |
| BHMT RV | ATGACCAGGTGGGACATGCAC | Amplicon size: 75 bp; E = 91%; R2 value = 0.99 |
| BHMT probe | AGAATTCCCCTTCGGTCTGGAGCCCA | |
| SAHHB FW | CCGCCGTGCTCATTGAGA | Amplicon size: 65 bp; E = 93%; R2 value = 0.99 |
| SAHHB RV | GTTCAATGGTCCAGCTGCAATATC | |
| SAHHB probe | CTGCCCTTGGAGCCGA | |
| CBS FW | AGACCATCAAGATCCTCAAGGAGAA | Amplicon size: 62 bp; E = 94%; R2 value = 0.99 |
| CBS RV | TCGTTGACGAGTCCGGC | |
| CBS probe | GGCTTTTGACCAGG |
| CTRL+ | CTRL− | Met-Met | L-Met | DL-Met | HMTBa | Co DL-Met | SEM | |
|---|---|---|---|---|---|---|---|---|
| M (%) | 3.70 ± 3.70 | 9.88 ± 2.14 | 6.17 ± 2.14 | 7.41 ± 5.24 | 8.64 ± 4.28 | 7.41 ± 3.07 | 6.17 ± 5.66 | 2.09 |
| iIBW (g) | 3.40 ± 0.03 | 3.41 ± 0.03 | 3.39 ± 0.02 | 3.40 ± 0.05 | 3.40 ± 0.02 | 3.41 ± 0.04 | 3.40 ± 0.03 | 0.01 |
| iFBW (g) | 21.40 ± 1.73 | 20.18 ± 1.38 | 20.05 ± 1.42 | 19.31 ± 1.72 | 20.66 ± 1.29 | 20.57 ± 2.23 | 20.99 ± 1.75 | 0.34 |
| WG (g) | 18.00 ± 1.76 | 16.77 ± 1.35 | 16.67 ± 1.42 | 15.91 ± 1.67 | 17.26 ± 1.23 | 17.17 ± 2.19 | 17.59 ± 1.72 | 0.33 |
| FCR | 0.82 ± 0.07 | 0.87 ± 0.11 | 0.82 ± 0.01 | 0.82 ± 0.03 | 0.81 ± 0.03 | 0.82 ± 0.090 | 0.78 ± 0.01 | 0.01 |
| PER | 2.87 ± 0.25 | 2.75 ± 0.34 | 2.83 ± 0.03 | 2.83 ± 0.11 | 2.93 ± 0.12 | 2.86 ± 0.30 | 3.02 ± 0.02 | 0.04 |
| SGR (%/d) | 3.13 ± 0.312 | 2.61 ± 0.59 | 2.97 ± 0.12 | 2.84 ± 0.06 | 2.98 ± 0.15 | 2.88 ± 0.20 | 3.04 ± 0.06 | 0.06 |
| FR (%/d) | 2.94 ± 0.16 | 2.47 ± 0.39 | 2.76 ± 0.12 | 2.62 ± 0.03 | 2.74 ± 0.06 | 2.65 ± 0.13 | 2.71 ± 0.07 | 0.04 |
| TGC | 0.16 ± 0.01 | 0.15 ± 0.01 | 0.15 ± 0.01 | 0.15 ± 0.011 | 0.16 ± 0.01 | 0.15 ± 0.01 | 0.16 ± 0.01 | 0.01 |
| CTRL+ | CTRL− | Met-Met | L-Met | DL-Met | HMTBa | Co DL-Met | SEM | p | |
|---|---|---|---|---|---|---|---|---|---|
| DM | 25.75 ± 0.52 ab | 26.62 ± 0.79 ab | 25.66 ± 0.40 b | 26.61 ± 0.56 ab | 26.28 ± 0.41 ab | 26.59 ± 0.19 ab | 26.71 ± 0.88 a | 0.10 | 0.009 |
| CP | 13.97 ± 0.31 c | 14.74 ± 0.61 abc | 14.18 ± 0.54 bc | 15.07 ± 0.70 ab | 14.79 ± 0.54 abc | 15.35 ± 0.35 a | 15.28 ± 0.57 a | 0.11 | 0.000 |
| EE | 8.47 ± 0.43 abc | 9.15 ± 060 a | 7.93 ± 0.27 c | 8.36 ± 0.36 bc | 8.54 ± 0.32 abc | 8.83 ± 0.71 ab | 8.66 ± 0.44 abc | 0.08 | 0.001 |
| Ash PG EG | 2.99 ± 0.12 b 2.49 ± 0.07 bcd 1.74 ± 0.09 a | 2.26 ± 0.09 c 2.48 ± 0.10 bcd 1.77 ± 0.13 a | 3.25 ± 0.061 a 2.35 ± 0.11 d 1.51 ± 0.05 a | 2.21 ± 0.11 c 2.41 ± 0.13 cd 1.54 ± 0.07 bc | 2.22 ± 0.16 c 2.56 ± 0.11 abc 1.69 ± 0.07 ab | 2.16 ± 0.06 c 2.66 ± 0.07 ab 1.74 ± 0.09 a | 2.35 ± 0.12 c 2.71 ± 0.13 a 1.74 ± 0.06 a | 0.07 | 0.000 <0.001 <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceccotti, C.; Biasato, I.; Gasco, L.; Caimi, C.; Bellezza Oddon, S.; Rimoldi, S.; Brambilla, F.; Terova, G. How Different Dietary Methionine Sources Could Modulate the Hepatic Metabolism in Rainbow Trout? Curr. Issues Mol. Biol. 2022, 44, 3238-3252. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44070223
Ceccotti C, Biasato I, Gasco L, Caimi C, Bellezza Oddon S, Rimoldi S, Brambilla F, Terova G. How Different Dietary Methionine Sources Could Modulate the Hepatic Metabolism in Rainbow Trout? Current Issues in Molecular Biology. 2022; 44(7):3238-3252. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44070223
Chicago/Turabian StyleCeccotti, Chiara, Ilaria Biasato, Laura Gasco, Christian Caimi, Sara Bellezza Oddon, Simona Rimoldi, Fabio Brambilla, and Genciana Terova. 2022. "How Different Dietary Methionine Sources Could Modulate the Hepatic Metabolism in Rainbow Trout?" Current Issues in Molecular Biology 44, no. 7: 3238-3252. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb44070223