
Urinary Proteome Differences in Patients with Type 2 Diabetes Pre and Post Liraglutide Treatment

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations and Informed Consent
2.2. Study Subjects
2.3. Urine Collection and Protein Extraction
2.4. Protein Labeling with Cyanine Dyes
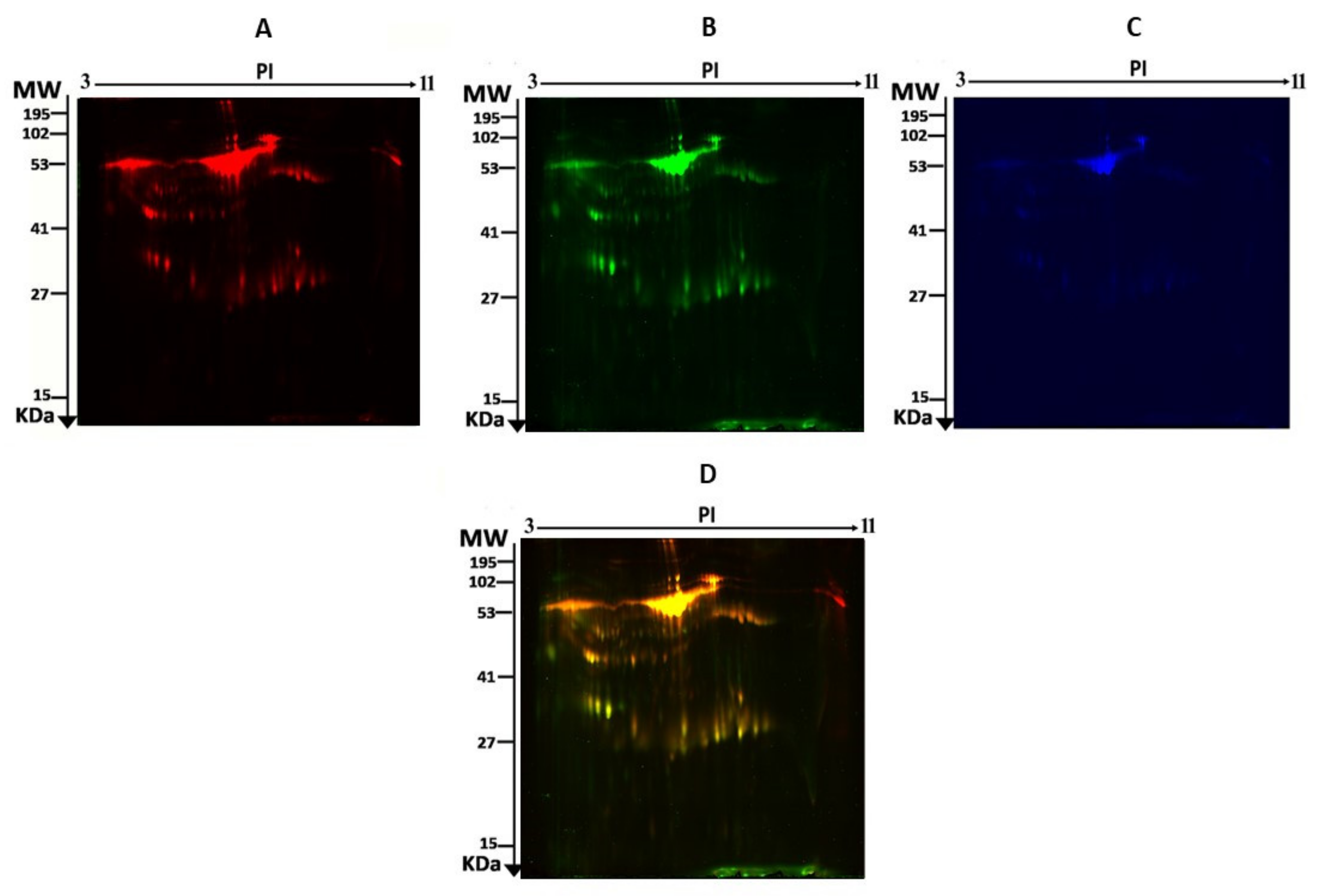
2.5. 2D-DIGE and Image Scanning
2.6. Colloidal Coomassie Blue Staining of the Preparative Gel
2.7. Statistical Analysis
2.8. Protein Digestion and MALDI Analysis
2.9. Bioinformatics Analysis
3. Results
3.1. Clinical and Biochemical Data
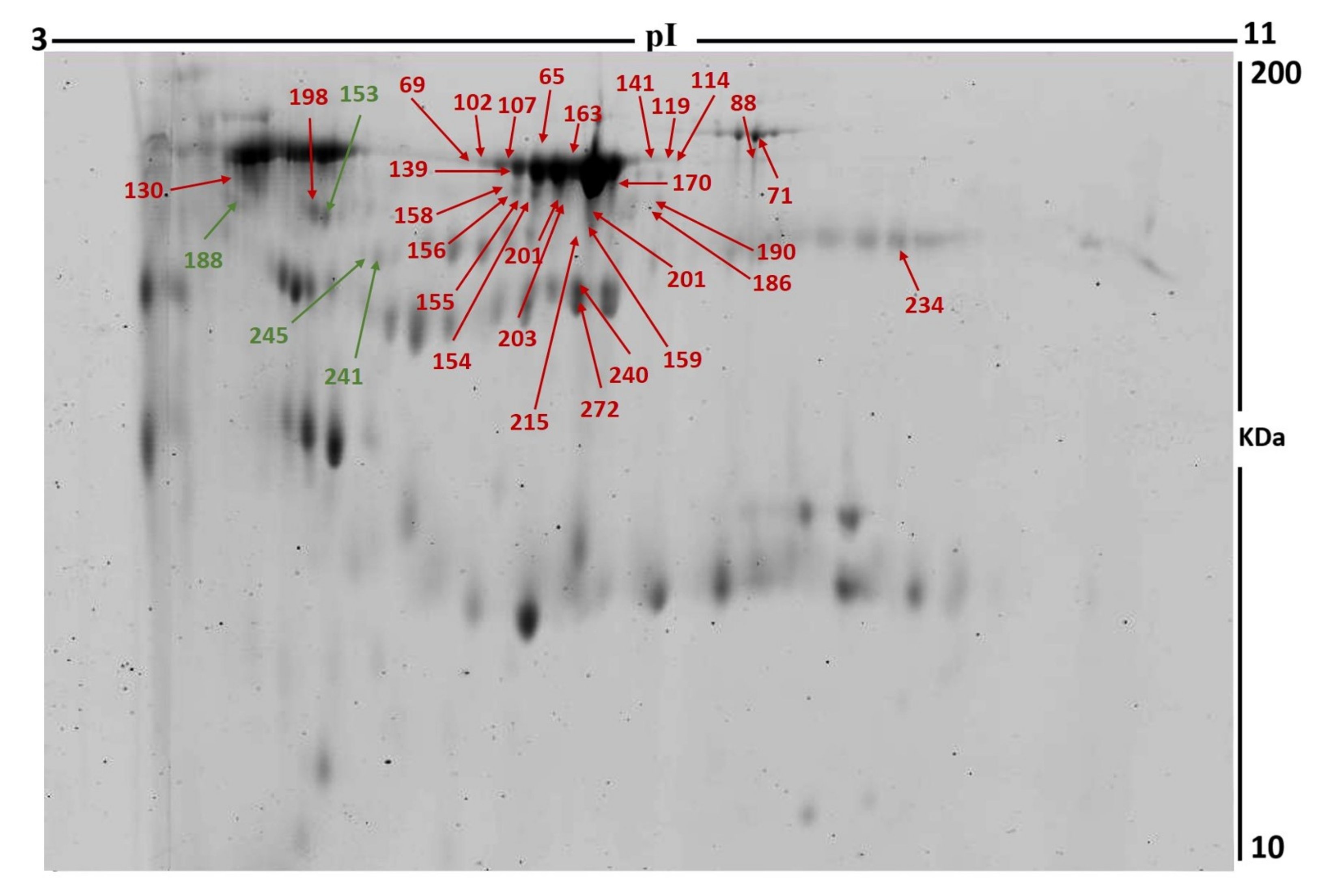
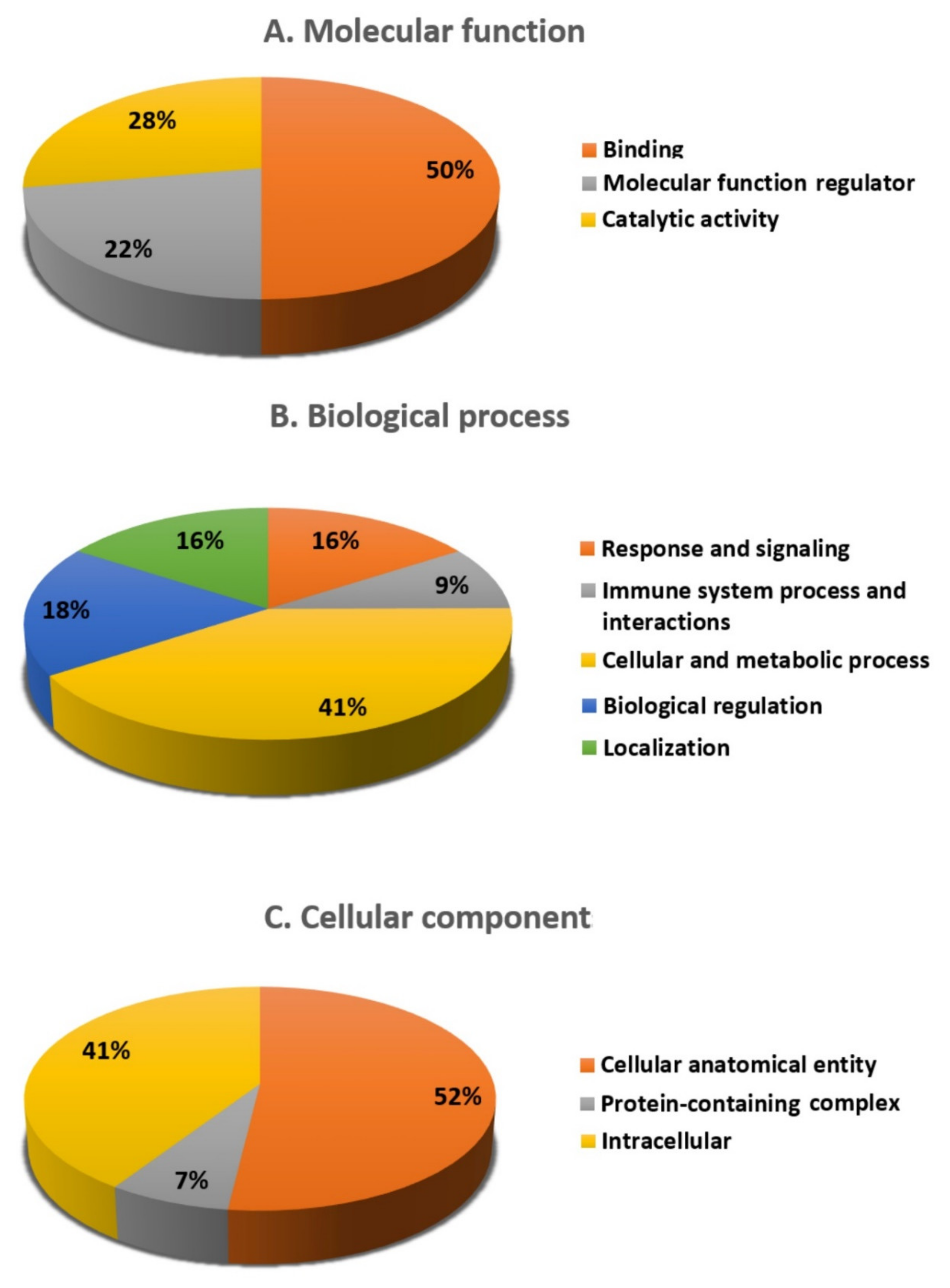
3.2. Proteomic Analysis and Identification of Differentially Expressed Proteins
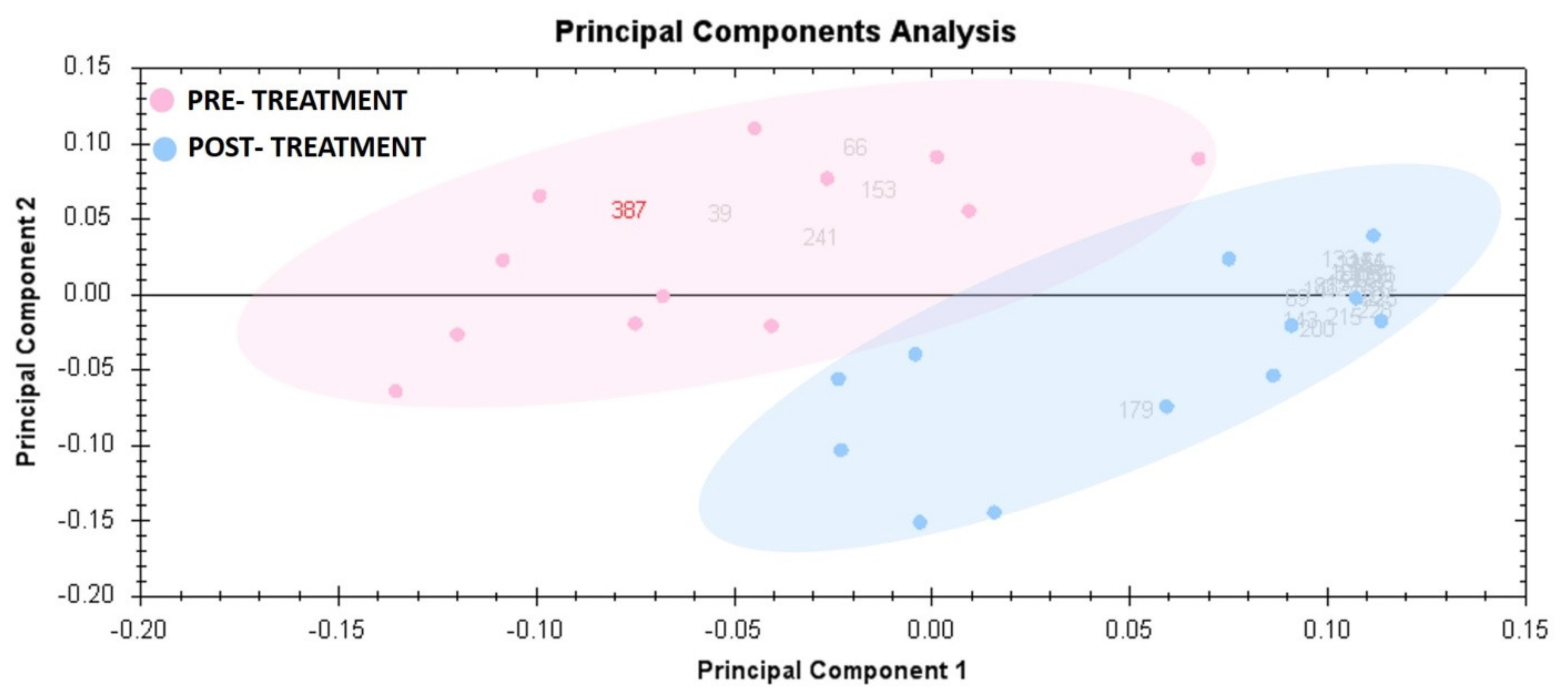
3.3. Principal Component Analysis
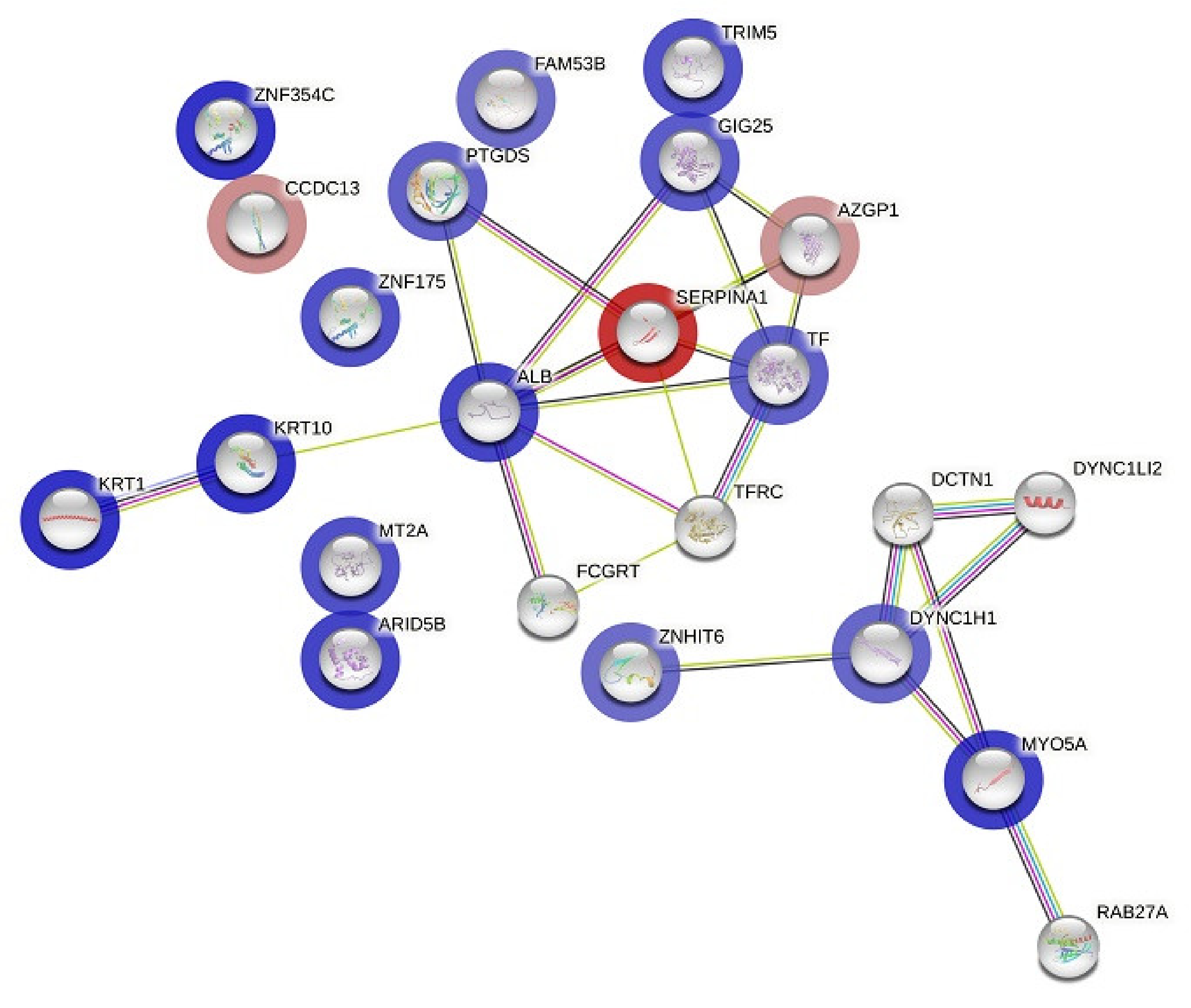
3.4. Protein–Protein Interaction (PPI) Network Construction
4. Discussion
4.1. Proteins Increased after Liraglutide Treatment
4.2. Proteins Decreased Post Liraglutide Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 10 October 2022).
- Raghavan, S.; Vassy, J.L.; Ho, Y.L.; Song, R.J.; Gagnon, D.R.; Cho, K.; Wilson, P.W.; Phillips, L.S. Diabetes mellitus–related all-cause and cardiovascular mortality in a national cohort of adults. J. Am. Heart Assoc. 2019, 8, e011295. [Google Scholar] [CrossRef] [PubMed]
- Lovshin, J.A.; Drucker, D.J. Incretin-based therapies for type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2009, 5, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Sauerwald, A.; Ritzel, R.; Holst, J.J.; Schmiegel, W. Influence of glucagon-like peptide 1 on fasting glycemia in type 2 diabetic patients treated with insulin after sulfonylurea secondary failure. Diabetes Care 1998, 21, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Hasanzad, M.; Sarhangi, N.; Nikfar, S.; Ostad, S.N.; Aghaei Meybodi, H.R. A narrative review of current trends in liraglutide: Insights into the unmet needs in management of type 2 diabetes and obesity. J. Diabetes Metab. Disord. 2020, 19, 1863–1872. [Google Scholar] [CrossRef]
- Palani, A.Z.; Halleck, Y.-Y.; Carrington, M. GLP-1 Receptor Agonists for the Treatment of Diabetes and Obesity; Chackalamannil, S., Rotella, D., Ward, S., Eds.; Elsevier: Oxford, UK, 2017. [Google Scholar]
- Ostawal, A.; Mocevic, E.; Kragh, N.; Xu, W. Clinical effectiveness of liraglutide in type 2 diabetes treatment in the real-world setting: A systematic literature review. Diabetes Ther. 2016, 7, 411–438. [Google Scholar] [CrossRef]
- Marso, S.P.; Daniels, G.H.; Brown-Frandsen, K.; Kristensen, P.; Mann, J.F.; Nauck, M.A.; Nissen, S.E.; Pocock, S.; Poulter, N.R.; Ravn, L.S. Liraglutide and cardiovascular outcomes in type 2 diabetes. N. Engl. J. Med. 2016, 375, 311–322. [Google Scholar] [CrossRef]
- Heuvelman, V.D.; Van Raalte, D.H.; Smits, M.M. Cardiovascular effects of glucagon-like peptide 1 receptor agonists: From mechanistic studies in humans to clinical outcomes. Cardiovasc. Res. 2020, 116, 916–930. [Google Scholar] [CrossRef]
- Hogan, A.E.; Gaoatswe, G.; Lynch, L.; Corrigan, M.A.; Woods, C.; O’Connell, J.; O’Shea, D. Glucagon-like peptide 1 analogue therapy directly modulates innate immune-mediated inflammation in individuals with type 2 diabetes mellitus. Diabetologia 2014, 57, 781–784. [Google Scholar] [CrossRef]
- Mali, N.; Su, F.; Ge, J.; Fan, W.X.; Zhang, J.; Ma, J. Efficacy of liraglutide in patients with diabetic nephropathy: A meta-analysis of randomized controlled trials. BMC Endocr. Disord. 2022, 22, 93. [Google Scholar] [CrossRef]
- Kristensen, S.L.; Rørth, R.; Jhund, P.S.; Docherty, K.F.; Sattar, N.; Preiss, D.; Køber, L.; Petrie, M.C.; McMurray, J.J. Cardiovascular, mortality, and kidney outcomes with GLP-1 receptor agonists in patients with type 2 diabetes: A systematic review and meta-analysis of cardiovascular outcome trials. Lancet Diabetes Endocrinol. 2019, 7, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Kawanami, D.; Takashi, Y. GLP-1 receptor agonists in diabetic kidney disease: From clinical outcomes to mechanisms. Front. Pharmacol. 2020, 11, 967. [Google Scholar] [CrossRef] [PubMed]
- Ekhzaimy, A.A.; Masood, A.; Benabdelkamel, H.; Elhassan, T.; Musambil, M.; Alfadda, A.A. Plasma proteomics reveals an improved cardio-metabolic profile in patients with type 2 diabetes post-liraglutide treatment. Diabetes Vasc. Dis. Res. 2022, 19, 14791641221094322. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Zhong, X.; Li, N.; Pan, T. Protective effects of liraglutide on glomerular podocytes in obese mice by inhibiting the inflammatory factor TNF-α-mediated NF-κB and MAPK pathway. Obes. Res. Clin. Pract. 2019, 13, 385–390. [Google Scholar] [CrossRef]
- Liljedahl, L.; Pedersen, M.H.; McGuire, J.N.; James, P. The impact of the glucagon-like peptide 1 receptor agonist liraglutide on the streptozotocin-induced diabetic mouse kidney proteome. Physiol. Rep. 2019, 7, e13994. [Google Scholar] [CrossRef]
- Chen, W.; Liu, X.; Ye, S. Effects of metformin on blood and urine pro-inflammatory mediators in patients with type 2 diabetes. J. Inflamm. 2016, 13, 34. [Google Scholar] [CrossRef]
- Wessel, D.; Flügge, U. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Masood, A.; Benabdelkamel, H.; Ekhzaimy, A.A.; Alfadda, A.A. Plasma-Based Proteomics Profiling of Patients with Hyperthyroidism after Antithyroid Treatment. Molecules 2020, 25, 2831. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Moustafa, A.; Sallam, R.; Bassas, A.; Duncan, M. Proteomic analysis of mature adipo cytes from obese patients in relation to aging. Exp. Gerontol. 2013, 48, 1196–1203. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Jammah, A.A.; Ekhzaimy, A.A. Differences in the plasma proteome of patients with hypothyroidism before and after thyroid hormone replacement: A proteomic analysis. Int. J. Mol. Sci. 2018, 19, 88. [Google Scholar] [CrossRef] [Green Version]
- Rigato, M.; Fadini, G.P. Comparative effectiveness of liraglutide in the treatment of type 2 diabetes. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 107. [Google Scholar]
- Yin, J.; Han, M.; Li, L.; Li, Y.; Liu, Z.a.; Yang, J.; Liu, Y. To assess liraglutide’s therapeutic effect in patients with type 2 diabetes mellitus using flash glucose monitoring system. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 4399. [Google Scholar] [CrossRef] [PubMed]
- Peradze, N.; Farr, O.M.; Perakakis, N.; Lázaro, I.; Sala-Vila, A.; Mantzoros, C.S. Short-term treatment with high dose liraglutide improves lipid and lipoprotein profile and changes hormonal mediators of lipid metabolism in obese patients with no overt type 2 diabetes mellitus: A randomized, placebo-controlled, cross-over, double-blind clinical trial. Cardiovasc. Diabetol. 2019, 18, 141. [Google Scholar]
- Davies, M.J.; Bain, S.C.; Atkin, S.L.; Rossing, P.; Scott, D.; Shamkhalova, M.S.; Bosch-Traberg, H.; Syrén, A.; Umpierrez, G.E. Efficacy and safety of liraglutide versus placebo as add-on to glucose-lowering therapy in patients with type 2 diabetes and moderate renal impairment (LIRA-RENAL): A randomized clinical trial. Diabetes Care 2016, 39, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Benabdelkamel, H.; Masood, A.; Okla, M.; Al-Naami, M.Y.; Alfadda, A.A. A proteomics-based approach reveals differential regulation of urine proteins between metabolically healthy and unhealthy obese patients. Int. J. Mol. Sci. 2019, 20, 4905. [Google Scholar] [CrossRef] [PubMed]
- Hennemann, G.; Krenning, E.P.; Docter, R. Thyroid hormone-binding plasma proteins. Front. Thyroidol. 1986, 1, 97–101. [Google Scholar]
- Benabdelkamel, H.; Masood, A.; Ekhzaimy, A.A.; Alfadda, A.A. Proteomics profiling of the urine of patients with hyperthyroidism after anti-thyroid treatment. Molecules 2021, 26, 1991. [Google Scholar] [CrossRef]
- Masood, A.; Benabdelkamel, H.; Jammah, A.A.; Ekhzaimy, A.A.; Alfadda, A.A. Identification of Protein Changes in the Urine of Hypothyroid Patients Treated with Thyroxine Using Proteomics Approach. ACS Omega 2021, 6, 2367–2378. [Google Scholar] [CrossRef]
- Cohen-Bucay, A.; Viswanathan, G. Urinary markers of glomerular injury in diabetic nephropathy. Int. J. Nephrol. 2012, 10, 11–24. [Google Scholar] [CrossRef]
- Siddiqui, K.; Joy, S.S.; George, T.P.; Mujammami, M.; Alfadda, A.A. Potential role and excretion level of urinary transferrin, KIM-1, RBP, MCP-1 and NGAL markers in diabetic nephropathy. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 5103. [Google Scholar] [CrossRef]
- Cai, L.; Klein, J.B.; Kang, Y.J. Metallothionein inhibits peroxynitrite-induced DNA and lipoprotein damage. J. Biol. Chem. 2000, 275, 38957–38960. [Google Scholar] [CrossRef] [PubMed]
- Cai, L. Metallothionein as an adaptive protein prevents diabetes and its toxicity. Nonlinearity Biol. Toxicol. Med. 2004, 2, 15401420490464367. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Ogawa, D.; Sogawa, N.; Asanuma, M.; Miyazaki, I.; Terami, N.; Hatanaka, T.; Horiguchi, C.S.; Nakatsuka, A.; Eguchi, J. Metallothionein deficiency exacerbates diabetic nephropathy in streptozotocin-induced diabetic mice. Am. J. Physiol.-Ren. Physiol. 2014, 306, F105–F115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snider, N.T. Kidney keratins: Cytoskeletal stress responders with biomarker potential. Kidney Int. 2016, 89, 738–740. [Google Scholar] [CrossRef]
- Shetty, S.; Gokul, S. Keratinization and its disorders. Oman Med. J. 2012, 27, 348. [Google Scholar] [CrossRef]
- Bing, C.; Bao, Y.; Jenkins, J.; Sanders, P.; Manieri, M.; Cinti, S.; Tisdale, M.J.; Trayhurn, P. Zinc-α2-glycoprotein, a lipid mobilizing factor, is expressed in adipocytes and is up-regulated in mice with cancer cachexia. Proc. Natl. Acad. Sci. USA 2004, 101, 2500–2505. [Google Scholar] [CrossRef]
- Russell, S.T.; Tisdale, M.J. Role of β-adrenergic receptors in the anti-obesity and anti-diabetic effects of zinc-α2-glycoprotien (ZAG). Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 590–599. [Google Scholar] [CrossRef]
- Yang, M.; Liu, R.; Li, S.; Luo, Y.; Zhang, Y.; Zhang, L.; Liu, D.; Wang, Y.; Xiong, Z.; Boden, G. Zinc-α2-glycoprotein is associated with insulin resistance in humans and is regulated by hyperglycemia, hyperinsulinemia, or liraglutide administration: Cross-sectional and interventional studies in normal subjects, insulin-resistant subjects, and subjects with newly diagnosed diabetes. Diabetes Care 2013, 36, 1074–1082. [Google Scholar]
- Balaz, M.; Vician, M.; Janakova, Z.; Kurdiova, T.; Surova, M.; Imrich, R.; Majercikova, Z.; Penesova, A.; Vlcek, M.; Kiss, A. Subcutaneous adipose tissue zinc-α2-glycoprotein is associated with adipose tissue and whole-body insulin sensitivity. Obesity 2014, 22, 1821–1829. [Google Scholar] [CrossRef]
- Elsheikh, M.; Elhefnawy, K.A.; Emad, G.; Ismail, M.; Borai, M. Zinc alpha 2 glycoprotein as an early biomarker of diabetic nephropathy in patients with type 2 diabetes mellitus. Braz. J. Nephrol. 2019, 41, 509–517. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.-M.; Zhang, S.; Zhao, J.-Y.; Liu, C.-Y. Adipokine zinc-alpha-2-glycoprotein as a novel urinary biomarker presents earlier than microalbuminuria in diabetic nephropathy. J. Int. Med. Res. 2016, 44, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Tanabe, J.; Ogura, Y.; Nagai, Y.; Sugaya, T.; Ohata, K.; Natsuki, Y.; Ichikawa, D.; Watanabe, S.; Inoue, K. Renoprotective effect of GLP-1 receptor agonist, liraglutide, in early-phase diabetic kidney disease in spontaneously diabetic Torii fatty rats. Clin. Exp. Nephrol. 2021, 25, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Xiang, Z.; Xiong, C.; Zhou, Q.; Wang, X.; Zou, H. Alpha1-antitrypsin in urinary extracellular vesicles: A potential biomarker of diabetic kidney disease prior to microalbuminuria. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 2037. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, L.; Zhang, J. Liraglutide exhibits anti-inflammatory activity through the activation of the PKA/CREB pathway. J. Inflamm. 2019, 16, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandström, C.S.; Ohlsson, B.; Melander, O.; Westin, U.; Mahadeva, R.; Janciauskiene, S. An association between Type 2 diabetes and alpha-antitrypsin deficiency. Diabet Med. 2008, 25, 1370–1373. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Treatment | Post-Treatment | p-Value | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
| Height (cm) | 155.2 ± 4.5 | 155.2 ± 4.5 | - |
| Weight (kg) | 87 ± 14 | 85.5 ± 11.5 | 0.40 |
| BMI (kg/m2) | 36.2 ± 6.1 | 36 ± 4.3 | 0.46 |
| HbA1C (%) | 9.6 ± 1.2 | 8.3 ± 1.7 | 0.006 * |
| Total cholesterol (mmol/L) | 4.5 ± 1.5 | 4.4 ± 1.3 | 0.43 |
| LDL (mmol/L) | 2.3 ± 1.4 | 2.3 ± 1.2 | 0.47 |
| HDL (mmol/L) | 1.3 ± 0.3 | 1.2 ± 0.5 | 0.44 |
| TG (mmol/L) | 1.9 ± 0.64 | 1.6 ± 0.42 | 0.13 |
| Urea (mmol/L) | 4.6 ± 2.3 | 4.4 ± 1.8 | 0.3 |
| Creatinine (µmol/L) | 61.3 ± 17.2 | 61.7 ± 19.1 | 0.47 |
| eGFR | 93.2 ± 26.7 | 96.1 ± 24.0 | 0.38 |
| A/C ratio | 17.3 ± 9.8 | 21.08 ± 18.0 | 0.3 |
| Sl No. | Spot No. a | Accession No. | Protein Name | MASCOT ID | p Value b (ANOVA) | Ratio c POST/PRE | Exp d |
|---|---|---|---|---|---|---|---|
| 1 | 159 | P02768 | Albumin | ALBU_HUMAN | 0.002 | 2.7 | UP |
| 2 | 201 | P04264 | Keratin, type II cytoskeletal 1 | K2C1_HUMAN | 0.003 | 2.7 | UP |
| 3 | 215 | P02768 | Albumin | ALBU_HUMAN | 0.003 | 2.3 | UP |
| 4 | 203 | Q14865 | AT-rich interactive domain-containing protein 5B | ARI5B_HUMAN | 0.003 | 2.4 | UP |
| 5 | 69 | P02768 | Albumin | ALBU_HUMAN | 0.005 | 2.4 | UP |
| 6 | 156 | P02768 | Albumin | ALBU_HUMAN | 0.005 | 2.5 | UP |
| 7 | 241 | P25311 | Zinc-alpha-2-glycoprotein | ZA2G_HUMAN | 0.005 | 0.48 | DOWN |
| 8 | 158 | P02768 | Albumin | ALBU_HUMAN | 0.006 | 2.4 | UP |
| 9 | 139 | Q86Y25 | Zinc finger protein 354C | Z354C_HUMAN | 0.009 | 2.7 | UP |
| 10 | 141 | Q9C035 | Tripartite motif-containing protein 5 | TRMIM5_HUMAN | 0.009 | 2.2 | UP |
| 11 | 153 | Q8IYE1 | Coiled-coil domain-containing protein 13 | CCD13_HUMAN | 0.009 | 0.45 | DOWN |
| 12 | 88 | Q9Y4I1 | Unconventional myosin-Va | MYO5A_HUMAN | 0.009 | 2.5 | UP |
| 13 | 154 | Q9Y473 | Zinc finger protein 175 | ZN175_HUMAN | 0.01 | 2.2 | UP |
| 14 | 155 | P02768 | Albumin | ALBU_HUMAN | 0.011 | 2.3 | UP |
| 15 | 227 | P02795 | Metallothionein-2 | MT2_HUMAN | 0.016 | 2.2 | UP |
| 16 | 114 | P02787 | Serotransferrin | TRFE_HUMAN | 0.019 | 2.4 | UP |
| 17 | 186 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.022 | 2.7 | UP |
| 18 | 163 | P02768 | Albumin | ALBU_HUMAN | 0.023 | 2.6 | UP |
| 19 | 245 | P25311 | Zinc-alpha-2-glycoprotein | ZA2G_HUMAN | 0.025 | 0.53 | DOWN |
| 20 | 65 | P02768 | Albumin | ALBU_HUMAN | 0.028 | 1.9 | UP |
| 21 | 272 | Q9NWK9 | Box C/D snoRNA protein 1 | BCD1_HUMAN | 0.028 | 1.7 | UP |
| 22 | 130 | P01011 | Alpha-1-antichymotrypsin | AACT_HUMAN | 0.037 | 2 | UP |
| 23 | 71 | P02787 | Serotransferrin | TRFE_HUMAN | 0.037 | 1.9 | UP |
| 24 | 188 | P01009 | Alpha-1-antitrypsin | A1AT_HUMAN | 0.04 | 0.19 | DOWN |
| 25 | 119 | P02768 | Albumin | ALBU_HUMAN | 0.04 | 2.4 | UP |
| 26 | 102 | Q14204 | Cytoplasmic dynein 1 heavy chain 1 | DYHC1_HUMAN | 0.043 | 2.1 | UP |
| 27 | 107 | P02768 | Albumin | ALBU_HUMAN | 0.052 | 2.8 | UP |
| 28 | 240 | P41222 | Prostaglandin-H2 D-isomerase | PTGDS_HUMAN | 0.053 | 1.9 | UP |
| 29 | 190 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.055 | 2.7 | UP |
| 30 | 234 | Q14204 | Cytoplasmic dynein 1 heavy chain 1 | DYHC1_HUMAN | 0.055 | 1.8 | UP |
| 31 | 198 | Q14153 | Protein FAM53B | FA53B_HUMAN | 0.055 | 1.7 | UP |
| 32 | 170 | P02768 | Albumin | ALBU_HUMAN | 0.066 | 2.4 | UP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafiullah, M.; Benabdelkamel, H.; Masood, A.; Ekhzaimy, A.A.; Musambil, M.; Joy, S.S.; Alfadda, A.A. Urinary Proteome Differences in Patients with Type 2 Diabetes Pre and Post Liraglutide Treatment. Curr. Issues Mol. Biol. 2023, 45, 1407-1421. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb45020092
Rafiullah M, Benabdelkamel H, Masood A, Ekhzaimy AA, Musambil M, Joy SS, Alfadda AA. Urinary Proteome Differences in Patients with Type 2 Diabetes Pre and Post Liraglutide Treatment. Current Issues in Molecular Biology. 2023; 45(2):1407-1421. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb45020092
Chicago/Turabian StyleRafiullah, Mohamed, Hicham Benabdelkamel, Afshan Masood, Aishah A. Ekhzaimy, Mohthash Musambil, Salini Scaria Joy, and Assim A. Alfadda. 2023. "Urinary Proteome Differences in Patients with Type 2 Diabetes Pre and Post Liraglutide Treatment" Current Issues in Molecular Biology 45, no. 2: 1407-1421. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb45020092