

Cytotoxic Effect of Andrographis paniculata Associated with 2-Aminoethyl Dihydrogen Phosphate in Triple-Negative Breast Cells
 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining the Crude Extract of Andrographis Paniculate
- −
- Andrographis extract (standard)/10.0% andrographolide;
- −
- Andrographis extract (standard)/30.0% andrographolide;
- −
- Andrographis extract (standard)/50.0% andrographolide.
2.2. Formulation of 2aminoethyl Dihydrogen Phosphate
2.3. Cell Culture
2.4. Determination of Cytotoxic Activity using the MTT Colorimetric Method
2.5. Analysis of Cell Cycle Phases and Fragmented DNA using Flow Cytometry
2.6. Mitochondrial Electrical Potential using Flow Cytometry
2.7. Evaluation of the Number of Cells Expressing Markers Involved in Apoptosis and Progression using Flow Cytometry
2.8. In Silico Analysis of the Pharmacological Effect of the Associations
2.9. Statistical Analyses
3. Results
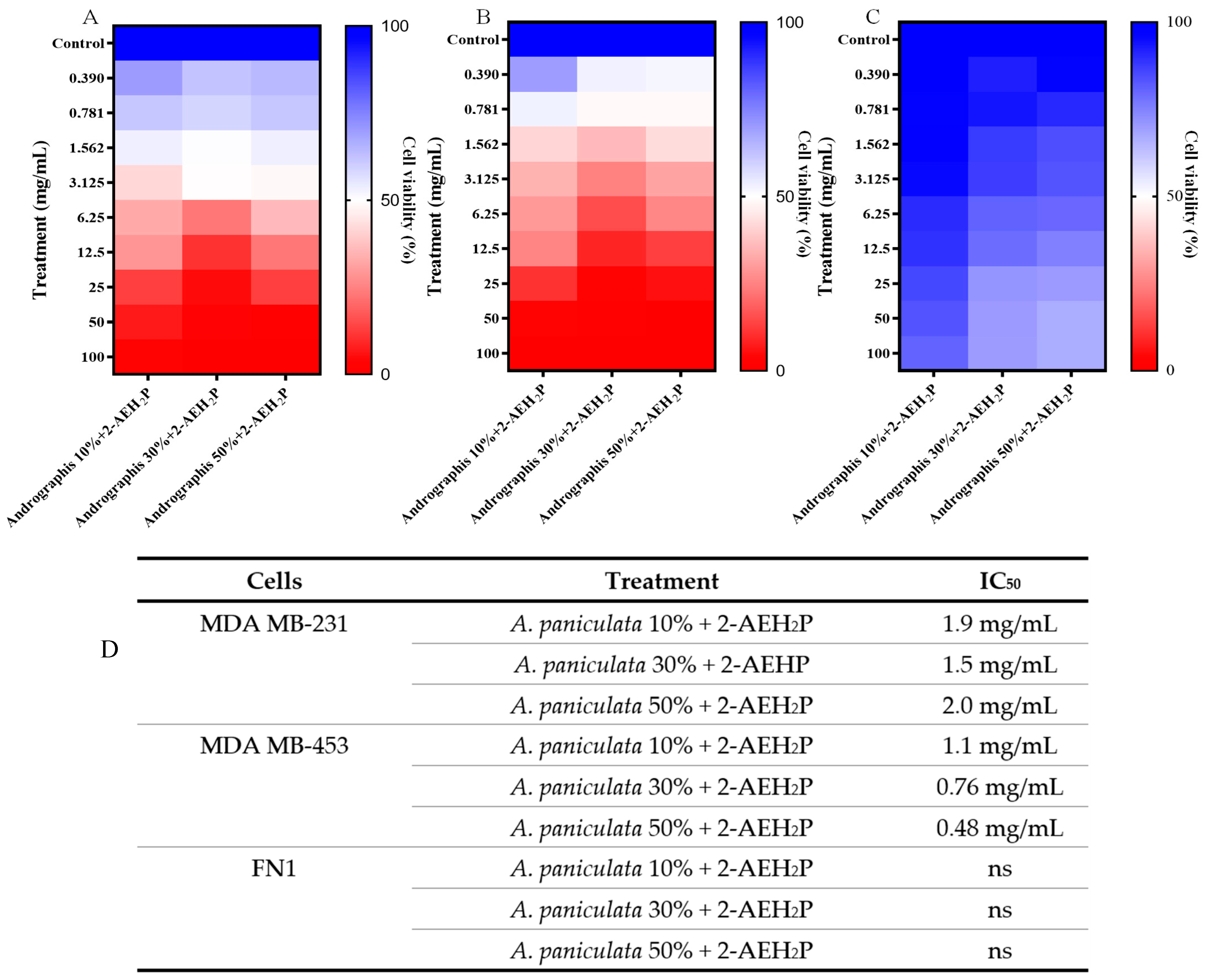
3.1. Cytotoxic Potential of A. paniculata and Its Association with 2-AEH2P
3.2. Distribution of Cell Populations Treated with A. paniculata and Its Association with 2-AEH2P in Cell Cycle Phases
3.3. Evaluation of the Mitochondrial Electrical Potential (ΔΨm) of Treatments with A. paniculata and Its Association with 2-AEH2P
3.4. Analysis of the Mechanisms of Action of Treatment with A. paniculata and Its Association with 2-AEH2P
3.5. Pharmacological Activity of the Association of A. paniculata with 2-AEH2P
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Bray, F.; Ilbawi, A.; Soerjomataram, I. Effect on Longevity of One-Third Reduction in Premature Mortality from Non-Communicable Diseases by 2030: A Global Analysis of the Sustainable Development Goal Health Target. Lancet Glob. Health 2018, 6, e1288–e1296. [Google Scholar] [CrossRef] [PubMed]
- Greenwell, M.; Rahman, P.K.S.M. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Li, Y.; Li, S.; Li, H.-B. Plant foods for the prevention and management of colon cancer. J. Funct. Foods 2018, 42, 95–110. [Google Scholar] [CrossRef]
- Esmeeta, A.; Adhikary, S.; Dharshnaa, V.; Swarnamughi, P.; Maqsummiya, Z.U.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Plant-derived bioactive compounds in colon cancer treatment: An updated review. Biomed. Pharmacother. 2022, 153, 113384. [Google Scholar] [CrossRef]
- Gupta, J.; Ahuja, A.; Gupta, R. Green Approaches for Cancers Management: An Effective Tool for Health Care. Anti-Cancer Agents Med. Chem. 2021, 22, 101–114. [Google Scholar] [CrossRef]
- Aiello, P.; Sharghi, M.; Mansourkhani, S.M.; Ardekan, A.P.; Jouybari, L.; Daraei, N.; Peiro, K.; Mohamadian, S.; Rezaei, M.; Heidari, M.; et al. Medicinal Plants in the Prevention and Treatment of Colon Cancer. Oxidative Med. Cell. Longev. 2019, 2019, 2075614. [Google Scholar] [CrossRef]
- Malik, Z.; Parveen, R.; Parveen, B.; Zahiruddin, S.; Khan, M.A.; Khan, A.; Massey, S.; Ahmad, S.; Husain, S.A. Anticancer potential of andrographolide from Andrographis paniculata (Burm.f.) Nees and its mechanisms of action. J. Ethnopharmacol. 2021, 272, 113936. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, Y.P. Comparison of different extraction methods and HPLC method development for the quantification of andrographolide from Andrographis paniculata (Burm.f.) Wall. ex Nees. Ann. Phytomed. Int. J. 2018, 7, 119–130. [Google Scholar] [CrossRef]
- Churiyah; Pongtuluran, O.B.; Rofaani, E. Tarwadi Antiviral and Immunostimulant Activities of Andrographis paniculata. HAYATI J. Biosci. 2015, 22, 67–72. [Google Scholar] [CrossRef]
- Khan, I.; Khan, F.; Farooqui, A.; Ansari, I.A. Andrographolide Exhibits Anticancer Potential Against Human Colon Cancer Cells by Inducing Cell Cycle Arrest and Programmed Cell Death via Augmentation of Intracellular Reactive Oxygen Species Level. Nutr. Cancer 2018, 70, 787–803. [Google Scholar] [CrossRef]
- Dai, Y.; Chen, S.-R.; Chai, L.; Zhao, J.; Wang, Y.; Wang, Y. Overview of pharmacological activities of Andrographis paniculata and its major compound andrographolide. Crit. Rev. Food Sci. Nutr. 2018, 59, S17–S29. [Google Scholar] [CrossRef]
- Conceição, T.D.O.; Cabral, L.G.D.S.; Laveli-Silva, M.G.; Pacheco, J.C.; Alves, M.G.; Rabelo, D.C.; Laiso, R.A.N.; Maria, D.A. New potential antiproliferative monophosphoester 2-aminoethyl dihydrogen phosphate in K-562 and K-562 MDR+ leukemia cells. Biomed. Pharmacother. 2021, 142, 112054. [Google Scholar] [CrossRef]
- da Silva, M.G.L.; Knop, L.B.; Maria, D.A. Meclizine chloridrate and methyl-β-cyclodextrin associated with monophosphoester synthetic phosphoethanolamine modulating proliferative potential in triple-negative breast cancer cells. J. Pharm. Pharmacol. 2019, 7, 408–420. [Google Scholar] [CrossRef]
- Cabral, L.G.d.S.; Hesse, H.; Freire, K.A.; de Oliveira, C.S.; Pedron, C.N.; Alves, M.G.; Carlstron, J.P.; Poyet, J.-L.; Oliveira, V.X.; Maria, D.A. The BR2 peptide associated with 2-aminoethyl dihydrogen phosphate is a formulation with antiproliferative potential for a triple-negative breast cancer model. Biomed. Pharmacother. 2022, 153, 113398. [Google Scholar] [CrossRef]
- da Silva, M.G.L.; Cabral, L.G.d.S.; Alves, M.G.; Conceição, T.d.O.; Hesse, H.; Laiso, R.A.N.; Rabelo, D.d.C.; Maria, D.A. 2-aminoethyl Dihydrogen Phosphate as a Modulator of Proliferative and Apoptotic Effects in Breast Cancer Cell Lines. J. Pharm. Pharmacol. 2021, 9, 83–97. [Google Scholar] [CrossRef]
- Silva, N.d.S. Extraction of bioactive compounds from unconventional food plants (pancs) from the semi-arid region. Federal University of Education, Sciences and Technology of Paraíba. Monography 2019, 1–30. Available online: https://repositorio.ifpb.edu.br/bitstream/177683/806/1/TCC%20NAYARA.pdf (accessed on 22 December 2023).
- Patil, R.; Jain, V. Andrographolide: A Review of Analytical Methods. J. Chromatogr. Sci. 2020, 59, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ahmed, H.; Yang, P.; Czinn, S.J.; Blanchard, T.G. Endoplasmic Reticulum Stress and IRE-1 Signaling Cause Apoptosis in Colon Cancer Cells in Response to Andrographolide Treatment. Oncotarget 2016, 7, 41432–41444. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Banerjee, V.; Czinn, S.; Blanchard, T. Increased Reactive Oxygen Species Levels Cause ER Stress and Cytotoxicity in Andrographolide Treated Colon Cancer Cells. Oncotarget 2017, 8, 26142–26153. [Google Scholar] [CrossRef]
- Mi, S.; Xiang, G.; Yuwen, D.; Gao, J.; Guo, W.; Wu, X.; Wu, X.; Sun, Y.; Su, Y.; Shen, Y.; et al. Inhibition of autophagy by andrographolide resensitizes cisplatin-resistant non-small cell lung carcinoma cells via activation of the Akt/mTOR pathway. Toxicol. Appl. Pharmacol. 2016, 310, 78–86. [Google Scholar] [CrossRef]
- Cheung, H.Y.; Cheung, S.H.; Li, J.; Cheung, C.S.; Lai, W.P.; Fong, W.F.; Leung, F.M. Andrographolide Isolated from Andrographis paniculate Induces Cell Cycle Arrest and Mitochondrial-Mediated Apoptosis in Human Leukemic HL-60 Cells. Planta Med. 2005, 71, 1106–1111. [Google Scholar] [CrossRef]
- Harjotaruno, S.; Widyawaruyanti, A.; Sismindari, S.; Zaini, N.C. Apoptosis Inducing Effect of Andrographolide on TD-47 Human Breast Cancer Cell Line. Afr. J. Tradit. Complement. Altern. Med. 2007, 4, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Alzaharna, M.; Alqouqa, I.; Cheung, H.-Y. Taxifolin synergizes Andrographolide-induced cell death by attenuation of autophagy and augmentation of caspase dependent and independent cell death in HeLa cells. PLoS ONE 2017, 12, e0171325. [Google Scholar] [CrossRef] [PubMed]
- Mir, H.; Kapur, N.; Singh, R.; Sonpavde, G.; Lillard, J.W., Jr.; Singh, S. Andrographolide inhibits prostate cancer by targeting cell cycle regulators, CXCR3 and CXCR7 chemokine receptors. Cell Cycle 2016, 15, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-M.; Kang, Y.-H.; Yang, X.; Wang, J.-F.; Zhang, Q.; Yang, B.-X.; Zhao, K.-L.; Xu, L.-P.; Yang, L.-P.; Ma, J.-X.; et al. Andrographolide radiosensitizes human esophageal cancer cell line ECA109 to radiation in vitro. Dis. Esophagus 2014, 29, 54–61. [Google Scholar] [CrossRef]
- Yang, P.-Y.; Hsieh, P.-L.; Wang, T.H.; Yu, C.-C.; Lu, M.-Y.; Liao, Y.-W.; Lee, T.-H.; Peng, C.-Y. Andrographolide Impedes Cancer Stemness and Enhances Radio-Sensitivity in Oral Carcinomas via MiR-218 Activation. Oncotarget 2016, 8, 4196–4207. [Google Scholar] [CrossRef]
- Shah, T.; Krishnamachary, B.; Wildes, F.; Wijnen, J.P.; Glunde, K.; Bhujwalla, Z.M. Molecular causes of elevated phosphoethanolamine in breast and pancreatic cancer cells. NMR Biomed. 2018, 31, e3936. [Google Scholar] [CrossRef]
- Lorenzo, V.P.; Mendonça Júnior FJ, B.; Scotti, L.; Scotti, M.T. Theoretical study of phosphoethanolamine: A synthetic anticancer agent with broad antitumor activity. J. Chem. 2016, 2016, 4068641. [Google Scholar] [CrossRef]
- Zhou, X.; Seto, S.W.; Chang, D.; Kiat, H.; Razmovski-Naumovski, V.; Chan, K.; Bensoussan, A. Synergistic effects of chinese herbal medicine: A comprehensive review of methodology and current research. Front. Pharmacol. 2016, 7, 201. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laiso, R.A.N.; Ferreira, J.C.; Rici, R.E.G.; Cabral, L.G.d.S.; Maria, D.A. Cytotoxic Effect of Andrographis paniculata Associated with 2-Aminoethyl Dihydrogen Phosphate in Triple-Negative Breast Cells. Curr. Issues Mol. Biol. 2024, 46, 527-541. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46010034
Laiso RAN, Ferreira JC, Rici REG, Cabral LGdS, Maria DA. Cytotoxic Effect of Andrographis paniculata Associated with 2-Aminoethyl Dihydrogen Phosphate in Triple-Negative Breast Cells. Current Issues in Molecular Biology. 2024; 46(1):527-541. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46010034
Chicago/Turabian StyleLaiso, Rosa Andrea Nogueira, Julia Carolina Ferreira, Rose Eli Grassi Rici, Laertty Garcia de Sousa Cabral, and Durvanei Augusto Maria. 2024. "Cytotoxic Effect of Andrographis paniculata Associated with 2-Aminoethyl Dihydrogen Phosphate in Triple-Negative Breast Cells" Current Issues in Molecular Biology 46, no. 1: 527-541. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46010034