
Optimizing Embryo Collection for Application of CRISPR/Cas9 System and Generation of Fukutin Knockout Rat Using This Method
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
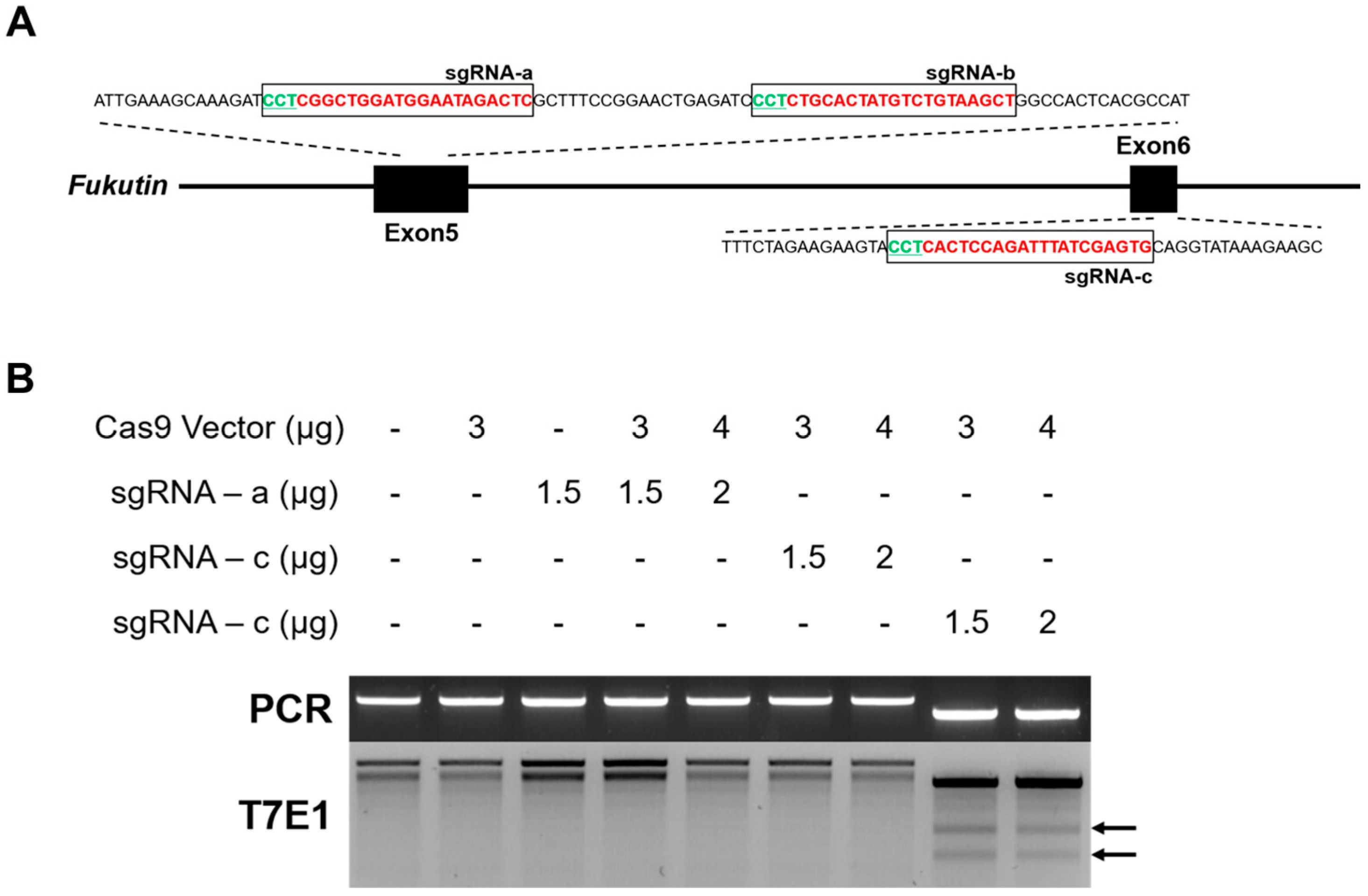
2.2. Generation of sgRNA and Selection of Optimal Target
2.3. PCR Analysis and T7E1 Assay
2.4. DNA Microinjection into Fertilized Eggs
2.5. Hormone Injection and Embryo Transfer into Surrogate Mother
2.6. Analysis of Skeletal Muscle Fiber
2.7. Statistical Analysis
3. Results
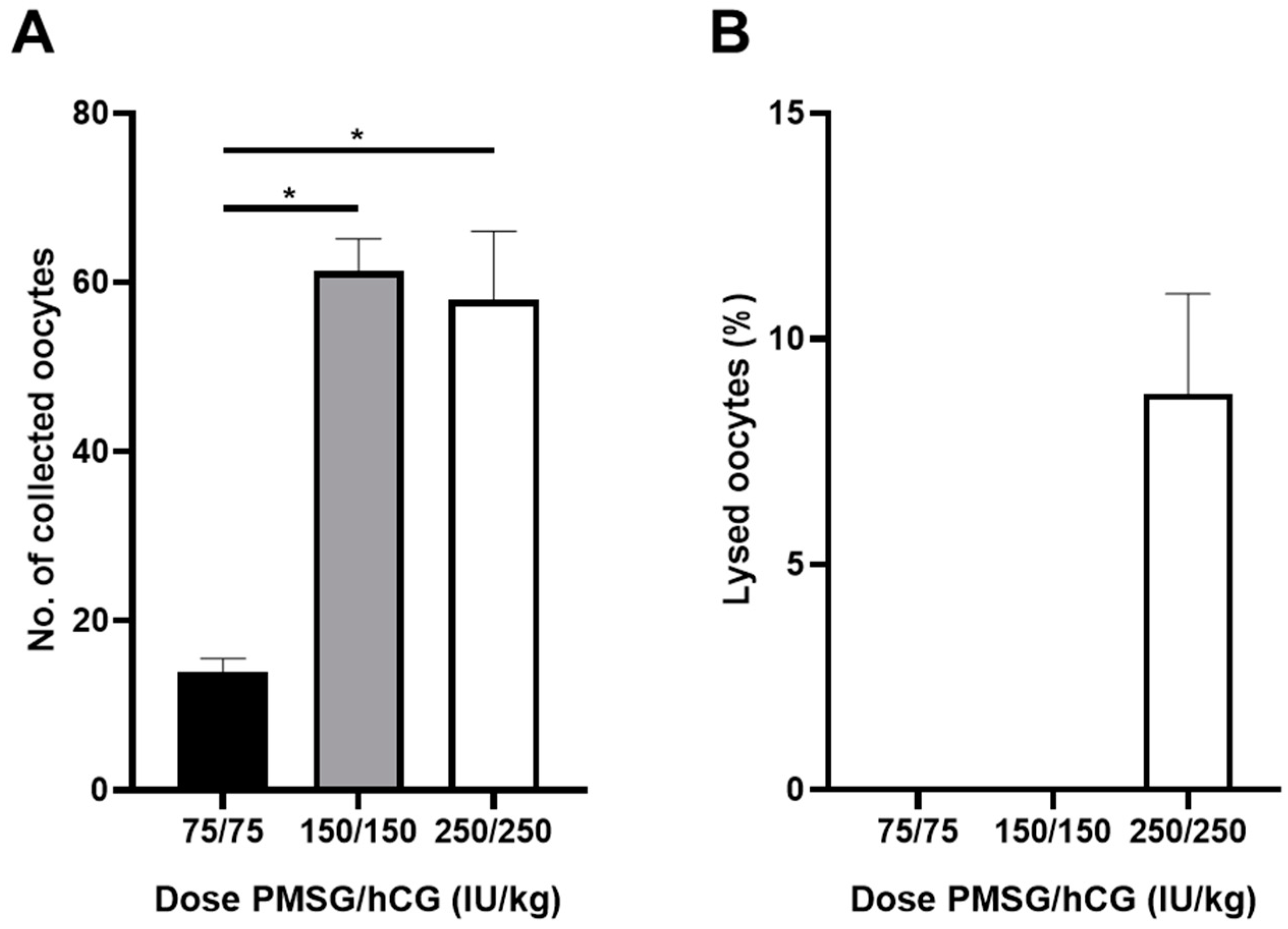
3.1. Establishment of Optimal Condition of Embryo Experimental System in Rat Animal Model
3.2. Construction of CRISPR/Cas9-Based Rat Fukutin Gene Knockout System
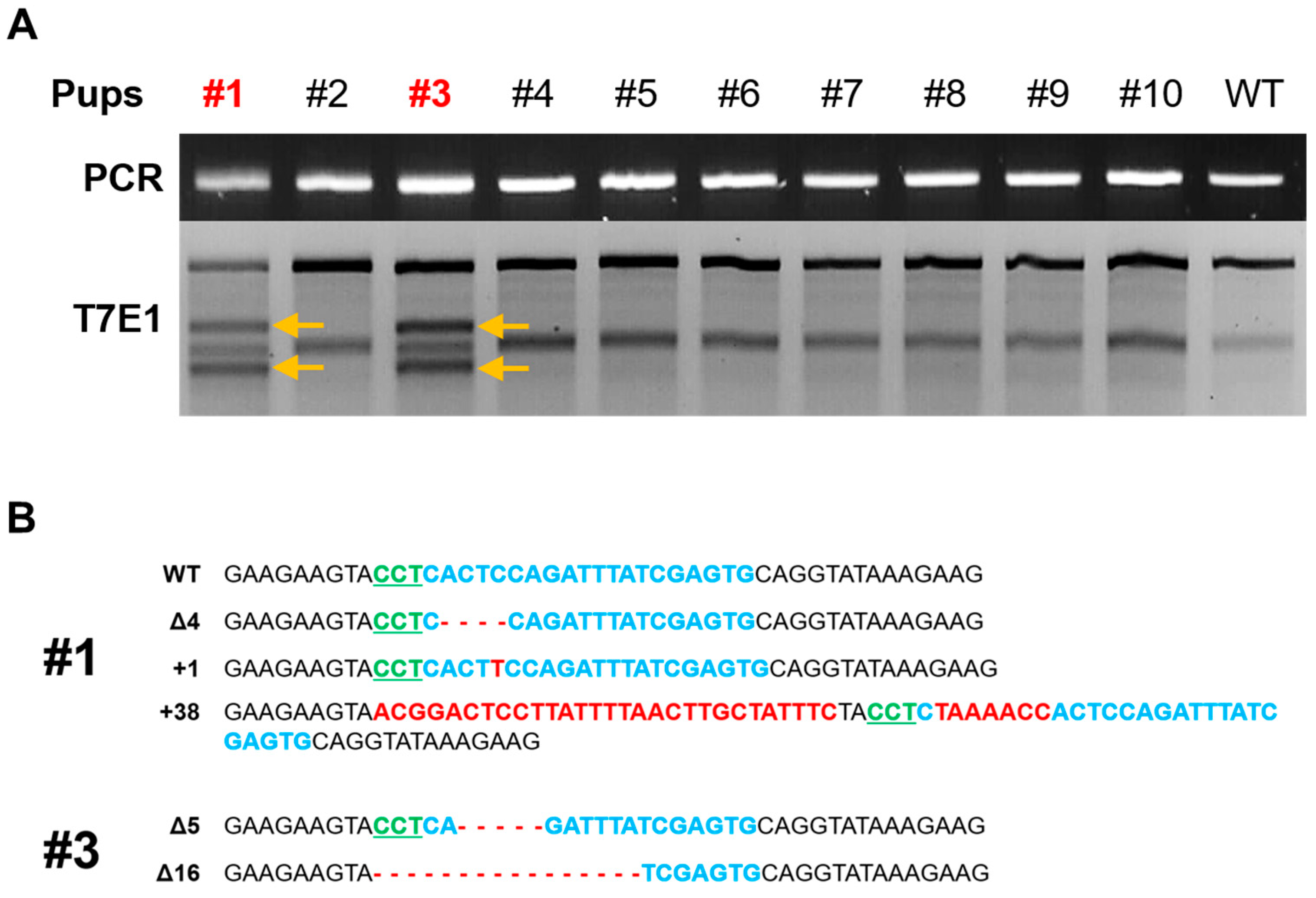
3.3. Production of Fukutin K/O Rat and Confirmation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mepham, T.B.; Combes, R.D.; Balls, M.; Barbieri, O.; Blokhuis, H.J.; Costa, P.; Crilly, R.E.; de Cock Buning, T.; Delpire, V.C.; O’Hare, M.J. The Use of Transgenic Animals in the European Union: The Report and Recommendations of ECVAM Workshop 28. Altern. Lab. Anim. 1998, 26, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Aitman, T.; Dhillon, P.; Geurts, A.M. A RATional choice for translational research? Dis. Model. Mech. 2016, 9, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, P.M.; Jacob, H.J. Rats! Dis. Model. Mech. 2009, 2, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Getz, G.S.; Reardon, C.A. Animal models of atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Dalkilic, I.; Kunkel, L.M. Muscular dystrophies: Genes to pathogenesis. Curr. Opin. Genet. Dev. 2003, 13, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Nakahori, Y.; Miyake, M.; Matsumura, K.; Kondo-Iida, E.; Nomura, Y.; Segawa, M.; Yoshioka, M.; Saito, K.; Osawa, M.; et al. An ancient retrotransposal insertion causes Fukuyama-type congenital muscular dystrophy. Nature 1998, 394, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Osawa, M.; Suzuki, H. Congenital progressive muscular dystrophy of the Fukuyama type—Clinical, genetic and pathological considerations. Brain Dev. 1981, 3, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Nakamura, H.; Tanaka, J. Cortical dysplasia in congenital muscular dystrophy with central nervous system involvement (Fukuyama type). J. Neuropathol. Exp. Neurol. 1984, 43, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Ruggieri, V.; Lubieniecki, F.; Meli, F.; Diaz, D.; Ferragut, E.; Saito, K.; Brockington, M.; Muntoni, F.; Fukuyama, Y.; Taratuto, A.L. Merosin-positive congenital muscular dystrophy with mental retardation, microcephaly and central nervous system abnormalities unlinked to the Fukuyama muscular dystrophy and muscular-eye-brain loci: Report of three siblings. Neuromuscul. Disord. 2001, 11, 570–578. [Google Scholar] [CrossRef]
- Sato, T.; Adachi, M.; Nakamura, K.; Zushi, M.; Goto, K.; Murakami, T.; Ishiguro, K.; Shichiji, M.; Saito, K.; Ikai, T.; et al. The gross motor function measure is valid for Fukuyama congenital muscular dystrophy. Neuromuscul. Disord. 2017, 27, 45–49. [Google Scholar] [CrossRef]
- Chan, Y.M.; Keramaris-Vrantsis, E.; Lidov, H.G.; Norton, J.H.; Zinchenko, N.; Gruber, H.E.; Thresher, R.; Blake, D.J.; Ashar, J.; Rosenfeld, J.; et al. Fukutin-related protein is essential for mouse muscle, brain and eye development and mutation recapitulates the wide clinical spectrums of dystroglycanopathies. Hum. Mol. Genet. 2010, 19, 3995–4006. [Google Scholar] [CrossRef]
- Saito, F.; Masaki, T.; Saito, Y.; Nakamura, A.; Takeda, S.; Shimizu, T.; Toda, T.; Matsumura, K. Defective peripheral nerve myelination and neuromuscular junction formation in fukutin-deficient chimeric mice. J. Neurochem. 2007, 101, 1712–1722. [Google Scholar] [CrossRef]
- Ujihara, Y.; Kanagawa, M.; Mohri, S.; Takatsu, S.; Kobayashi, K.; Toda, T.; Naruse, K.; Katanosaka, Y. Elimination of fukutin reveals cellular and molecular pathomechanisms in muscular dystrophy-associated heart failure. Nat. Commun. 2019, 10, 5754. [Google Scholar] [CrossRef]
- Southern, W.M.; Nichenko, A.S.; Qualls, A.E.; Portman, K.; Gidon, A.; Beedle, A.M.; Call, J.A. Mitochondrial dysfunction in skeletal muscle of fukutin-deficient mice is resistant to exercise- and 5-aminoimidazole-4-carboxamide ribonucleotide-induced rescue. Exp. Physiol. 2020, 105, 1767–1777. [Google Scholar] [CrossRef]
- Gaina, G.; Popa Gruianu, A. Muscular dystrophy: Experimental animal models and therapeutic approaches (Review). Exp. Ther. Med. 2021, 21, 610. [Google Scholar] [CrossRef]
- Cho, S.W.; Lee, J.; Carroll, D.; Kim, J.S.; Lee, J. Heritable gene knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA ribonucleoproteins. Genetics 2013, 195, 1177–1180. [Google Scholar] [CrossRef]
- Li, W.; Teng, F.; Li, T.; Zhou, Q. Simultaneous generation and germline transmission of multiple gene mutations in rat using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 684–686. [Google Scholar] [CrossRef]
- Mukumoto, S.; Mori, K.; Ishikawa, H. Efficient induction of superovulation in adult rats by PMSG and hCG. Exp. Anim. 1995, 44, 111–118. [Google Scholar] [CrossRef]
- Miyoshi, K. Development of a culture medium for rat 1-cell embryos. J. Mamm. Ova Res. 2016, 33, 11–16. [Google Scholar] [CrossRef]
- Montague, T.G.; Cruz, J.M.; Gagnon, J.A.; Church, G.M.; Valen, E. CHOPCHOP: A CRISPR/Cas9 and TALEN web tool for genome editing. Nucleic Acids Res. 2014, 42, W401–W407. [Google Scholar] [CrossRef]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Haeussler, M.; Schonig, K.; Eckert, H.; Eschstruth, A.; Mianne, J.; Renaud, J.B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 2016, 17, 148. [Google Scholar] [CrossRef] [PubMed]
- Cradick, T.J.; Fine, E.J.; Antico, C.J.; Bao, G. CRISPR/Cas9 systems targeting beta-globin and CCR5 genes have substantial off-target activity. Nucleic Acids Res. 2013, 41, 9584–9592. [Google Scholar] [CrossRef] [PubMed]
- de Souza, N. Missing the target? Nat. Methods 2013, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shao, Y.; Guan, Y.; Li, L.; Wu, L.; Chen, F.; Liu, M.; Chen, H.; Ma, Y.; Ma, X.; et al. Large genomic fragment deletion and functional gene cassette knock-in via Cas9 protein mediated genome editing in one-cell rodent embryos. Sci. Rep. 2015, 5, 17517. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.A.; Pardo, P.S.; Cox, G.A.; Boriek, A.M. Early mechanical dysfunction of the diaphragm in the muscular dystrophy with myositis (Ttnmdm) model. Am. J. Physiol. Cell Physiol. 2008, 295, C1092–C1102. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, H.; Taniguchi, M.; Meno, C.; Taniguchi, Y.; Takeda, S.; Horie, M.; Otani, H.; Toda, T. Basement membrane fragility underlies embryonic lethality in fukutin-null mice. Neurobiol. Dis. 2005, 19, 208–217. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PMSG/hCG (IU/kg) | 2–4 Cell (%) | 8–16 Cell (%) | Blastocyst (%) | |||
|---|---|---|---|---|---|---|
| M16 | mR1ECM | M16 | mR1ECM | M16 | mR1ECM | |
| 75/75 | 85.0 ± 7.63 | 100.0 ± 0.00 * | 0.0 ± 0.00 | 52.9 ± 18.06 * | 0.0 ± 0.00 | 24.5 ± 19.25 * |
| 150/150 | 100.0 ± 0.00 † | 98.8 ± 0.62 | 0.0 ± 0.00 | 78.3 ± 2.69 *,† | 0.0 ± 0.00 | 49.0 ± 1.56 *,† |
| 250/250 | 79.7 ± 9.86 | 93.4 ± 3.97 * | 0.0 ± 0.00 | 66.8 ± 7.13 * | 0.0 ± 0.00 | 45.1 ± 6.94 * |
| Match Name | Coordinate | Spacer + PAM | Number of Mismatch | Out-of-Frame Score |
|---|---|---|---|---|
| Fukutin sgRNA-a | Chr 5: 73158279-73158257 | GAGTCTATTCCATCCAGCCGAGG | 0 | 77.5790797987 |
| Fukutin sgRNA-b | Chr 5: 73158319-73158299 | AGCTTACAGACATAGTGCAGAGG | 0 | 73.0049498974 |
| Fukutin sgRNA-c | Chr 5: 73160527-73160505 | CACTCGATAAATCTGGAGTGAGG | 0 | 84.1311264059 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seol, D.-W.; Park, B.-J.; Koo, D.-B.; Kim, J.-S.; Jeon, Y.-H.; Lee, J.-E.; Park, J.-S.; Jang, H.; Wee, G. Optimizing Embryo Collection for Application of CRISPR/Cas9 System and Generation of Fukutin Knockout Rat Using This Method. Curr. Issues Mol. Biol. 2024, 46, 3752-3762. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050234
Seol D-W, Park B-J, Koo D-B, Kim J-S, Jeon Y-H, Lee J-E, Park J-S, Jang H, Wee G. Optimizing Embryo Collection for Application of CRISPR/Cas9 System and Generation of Fukutin Knockout Rat Using This Method. Current Issues in Molecular Biology. 2024; 46(5):3752-3762. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050234
Chicago/Turabian StyleSeol, Dong-Won, Byoung-Jin Park, Deog-Bon Koo, Ji-Su Kim, Yong-Hyun Jeon, Jae-Eon Lee, Joon-Suk Park, Hoon Jang, and Gabbine Wee. 2024. "Optimizing Embryo Collection for Application of CRISPR/Cas9 System and Generation of Fukutin Knockout Rat Using This Method" Current Issues in Molecular Biology 46, no. 5: 3752-3762. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050234