Oxidative Stress and Antioxidant Biomarkers in Clinical and Experimental Models of Non-Alcoholic Fatty Liver Disease

1
Department of Biochemistry, College of Biosciences, Federal University of Agriculture, Abeokuta, Nigeria
2
Biochemistry Division, Department of Chemical Sciences, Faculty of Natural Sciences, Ajayi Crowther University, Oyo, Nigeria
*
Author to whom correspondence should be addressed.
Medicina 2019, 55(2), 26; https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55020026
Submission received: 5 January 2019
/
Accepted: 18 January 2019
/
Published: 24 January 2019
Abstract
:Non-alcoholic fatty liver disease (NAFLD) is a term that covers a range of hepatic disorders involving fat deposits in the liver. NAFLD begins with simple steatosis and progresses into non-alcoholic steatohepatitis (NASH) characterised by inflammation, fibrosis, apoptosis, oxidative stress, lipid peroxidation, mitochondrial dysfunction and release of adipokines and pro-inflammatory cytokines. Oxidative stress and antioxidants are known to play a vital role in the pathogenesis and severity of NAFLD/NASH. A number of oxidative stress and antioxidant markers are employed in the assessment of the pathological state and progression of the disease. In this article, we review several biomarkers of oxidative stress and antioxidants that have been measured at clinical and experimental levels. Also included is a comprehensive description of oxidative stress, sources and contribution to the pathogenesis of NAFLD/NASH.
Keywords:
liver; NAFLD; NASH; biomarkers; reactive species; oxidative stress; lipid peroxidation; antioxidants1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is a range of hepatic disorders associated with fatty deposits in liver, which occur in the absence of alcohol consumption or alcohol abuse [1]. NAFLD begins with an initial stage of fatty liver also known as hepatic steatosis (excessive fat loading in the hepatocytes). The progression from steatosis into cirrhosis of the liver due to inflammation and fibrosis results in irreversible damage to the liver [2]. This condition is called non-alcoholic steatohepatitis (NASH)—a term first introduced by Ludwig et al. [3] in clinical subjects with no history of alcohol consumption or abuse.
NAFLD is one of the most common chronic hepatic pathology. It has a worldwide distribution with an estimated prevalence of 25% for NAFLD and about 5% for NASH [4]. The highest prevalence of NAFLD is observed in Western countries (17% to 46%) where it is the most common chronic liver disease (CLD) in adults with a high prevalence of NASH in the United States (16%). The World Gastroenterology Organisation suggests that the prevalence of NAFLD had doubled over the last 20 years. NAFLD and NASH are closely associated with diabetes and obesity, and together are considered one of the major causes of liver disease in Western countries [5].
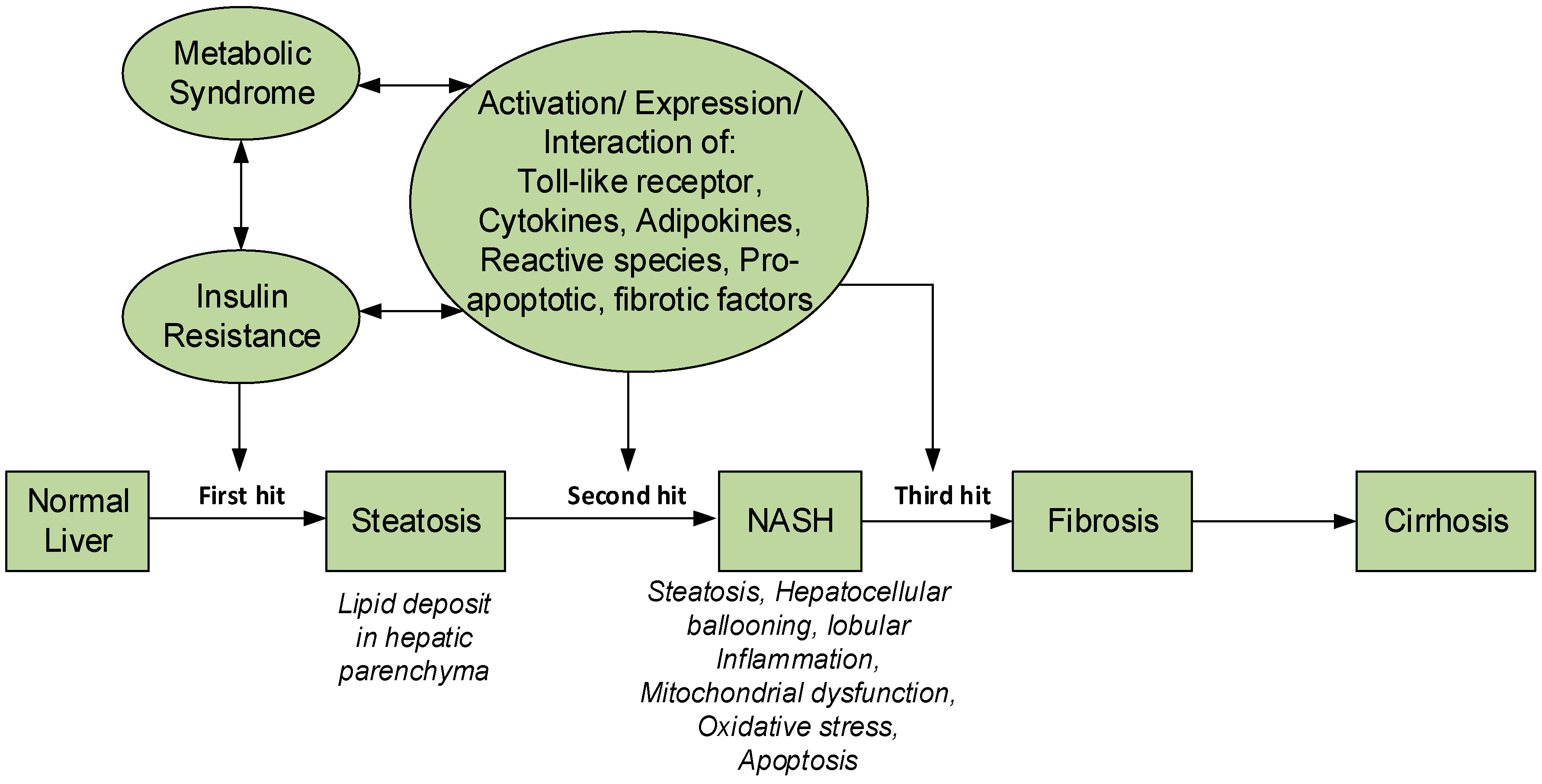
The pathophysiology of NASH was originally explained by the “two-hit” hypothesis [6]. In this hypothesis, the first hit is responsible for producing steatosis (fat accumulation in liver), and the second hit is from oxidative stress causing lipid peroxidation. The pathogenesis and progression of NAFLD is complex, and was not completely explained by the “two hit” hypothesis. Currently, NASH is described by the “multiple hit” hypothesis [7]. In this hypothesis, metabolic syndrome plays a major role due to insulin resistance and the inflammatory process mediated by interaction of different proteins and immune system. The components of the multiple “hits” are yet to be fully defined and they may vary in different patients. However, from available information, the “first hit” is caused by metabolic syndrome and insulin resistance, increased fat loading in hepatocytes leading to steatosis and liver injury. The accumulation of fat in the liver occur as a result of imbalance between the rate of influx and removal of triglycerides—a mechanism thought to protect hepatocytes from the lipotoxicity that may result from excessive influx of free fatty acids (FFAs) [8]. Most of the FFAs stored as triglycerides originate from increased lipolysis in peripheral tissues as a result of adipose tissue insulin resistance (IR), attended by increased lipogenesis due to hyperinsulinemia and diet fat. The subsequent “hits” are responsible for the inflammation, fibrosis, apoptosis, oxidative stress (OS), and hepatic lipid peroxidation, mitochondrial dysfunction, release of adipokines and pro-inflammatory cytokines [7] (Figure 1). Major histopathological features of NASH are steatosis, hepatocellular ballooning, lobular inflammation and in some cases, fibrosis [7,8] and may progress into cirrhosis if left unchecked.
An understanding of the mechanism of OS, its regulation as well as its role in NAFLD is vital. This will provide researchers in the area of NAFLD/NASH with the best choice of OS/antioxidant (AO) biomarkers useful in pre-clinical investigations as well as in clinical diagnosis of NAFLD. Development of potent drugs in the treatment of NAFLD will also take the AO action required to counteract the OS associated with NAFLD into consideration. In this article, we review the role of oxidative stress in NAFLD and several AO and OS biomarkers that have been measured in pre-clinical and clinical evaluations.
2. Oxidative Stress
OS refers to an imbalance between the production of reactive species (RS) and AO defenses [9]. A more encompassing definition describes it as “an imbalance between oxidants and AOs in favour of the oxidants, leading to a disruption of redox signalling and control and/or molecular damage” [9,10]. RS (the oxidants) are chemically reactive species containing oxygen (reactive oxygen species, ROS), or nitrogen (reactive nitrogen species, RNS), etc. (Table 1). ROS (which are the most extensively studied RS) are oxygen-containing molecules that exhibit higher chemical reactivity than oxygen (O2). Some ROS are free radicals (e.g., hydroxyl radical, superoxide radical, peroxyl radical etc.), others are non-radicals (e.g., hydrogen peroxide, hypochlorous acid, lipid peroxides etc.). RNS that are free radicals include nitric oxide, nitrogen dioxide etc., and those that are non-radicals include dinitrogen trioxide, peroxinitrite, etc. (Table 1). OS has been classified according to severity as “eustress” (physiological oxidative stress) and “distress” (toxic oxidative burden which damages biomolecules) [10,11]. In other words, low exposure to OS is useful for redox signalling, whereas high exposure results in disruption of redox signalling and causes damage to important biomolecules.
3. Oxidative Damage to Macromolecules
OS is associated with many diseases, especially those with an inflammatory mechanism [12]. OS is known in several hepatic diseases with high levels of ROS and RNS, which is an important description of the severity and progression of the disease [13,14]. ROS is constantly generated in the cell due to partial reduction of O2 or as a result of transfer of energy to O2. ROS can attack vital cell components like polyunsaturated fatty acids, proteins, and nucleic acids, and also carbohydrates in a few cases [15,16,17]. They can disrupt membrane properties like fluidity and ion transport, cause loss of enzyme activity, disruption of the protein synthesis mechanism and induction of DNA damage, ultimately leading to cell death. Damage resulting from OS is often called “oxidative damage”.
Oxidative damage to macromolecules (lipids, proteins, DNA etc.) results in formation of oxidative damage products (Table 2), which are often measured as biomarkers of OS [18,19,20,21]. Some of the most important oxidative damage products of lipids that are frequently measured includes malondialdehyde, lipid peroxides, 8-isoprostane, and 4-hydroxy-2-nonenal (4-HNE). Oxidation of protein can result in protein cross linkage, formation of protein carbonyls, and modification of amino acid. Important products of oxidative modification of amino acids that are frequently measured include 3-nitrotyrosine (a product of ROS-mediated nitration of tyrosine), 2-oxohistidine and hydroxyproline (Table 2). DNA/RNA oxidation can cause single or double strand fragmentation or modification of bases or sugar. 8-Hydroxy-2′-deoxyguanosine (8-OH-dG) and 8-hydroxyguanine (8-OH-G) are the most commonly measured DNA/RNA damage products.
4. Regulation of OS
There are several mechanisms for the cellular regulation of OS, which are extremely important to the cell homeostasis. This is achieved through the antioxidant system, which can control the formation of ROS or RNS and also repair oxidative damage to cells. An antioxidant is any substance that can inhibit the oxidation of the cell components such as DNA, proteins and lipids. Several levels of antioxidative defense mechanism are used to prevent oxidative damage [22,23]. The antioxidants can be derived from the diet or endogenously.
Endogenously derived antioxidants are classified as enzymic or non-enzymic antioxidants (Table 3). Enzymic antioxidants of endogenous origin include superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione reductase, etc. SOD catalyse the dismutation of superoxide radical to hydrogen peroxide. The hydrogen peroxide formed in this process and other processes is detoxified by the action of catalase or glutathione peroxidase utilising the power of reduced glutathione (GSH). The latter becomes oxidised (to GSSG) and can be reduced back to GSH through the action of glutathione reductase. Some of the most physiologically vital non-enzymic antioxidants include ascorbate, glutathione, α-tocopherol (vitamin E), Ubiquinone, Thioredoxin (TRX), Bilirubin etc. These antioxidant substances are effective in the detoxification of free radicals and reactive species in the cell (Table 3).
5. Sources and Role of OS in NAFLD/NASH
The sources of OS, as well as its role in NASH has been extensively reviewed by Koek et al. [24] and Tariq et al. [25]. OS results from excessive generation of reactive species (RS) or depletion of physiological redox homeostasis. RS (ROS or RNS) from the inflammatory response, the mitochondria, endoplasmic reticulum and peroxisomes also contribute to OS associated with NAFLD/NASH. Their sources and role in NAFLD/NASH are summarized in Table 4.
The mitochondria are important contributors to the OS observed in NAFLD. In the hepatocytes, the mitochondria are not only involved in respiration but they participate in anaplerotic pathways including gluconeogenesis and other biosynthetic activities. During NAFLD, mitochondrial β-oxidation produces ROS that damage hepatocytes contributing to inflammation, and other responses [28]. This increase in ROS production is related to increases in free fatty acid delivery in NAFLD. An increase in fatty acid induces oxidative metabolism, causing an increase in the level of OS and inflammation. The peroxisomal β-oxidation also contributes to the OS in NAFLD through generation of H2O2. Other sources of OS include the mitochondrial electron transport chain, the activities of microsomal cytochrome P450 enzymes, endoplasmic reticulum stress, the activity of xanthine oxidase and inflammatory responses. Their various contributions to OS in NAFLD are highlighted in Table 4.
OS has been reported to play a significant role in the pathophysiological mechanism of NAFLD and NASH [44,45]. Studies conducted in humans and animal models showed a strong association between the level of OS and the severity of NASH [26,46]. Although clinical and experimental studies have reported higher levels of lipid peroxidation in NASH patients, the levels of circulating antioxidants have been less reported.
6. Antioxidant and OS Markers Measured in NAFLD/NASH
Several biomarkers of oxidative stress and antioxidants have been detected in clinical and experimental models of NAFLD and NASH. Most of these are assayed predominantly in the liver, serum, plasma, and in a few cases, in whole blood samples. Major assay procedures for the detection of these markers include colorimetry, ELISA, and immunohistochemistry.
6.1. Antioxidant Markers Measured in Clinical NAFLD/NASH
At the clinical level, a number of antioxidants have been determined in the evaluation of NAFLD/NASH. They include the enzymic antioxidants: CAT, SOD, GPx, GR [47,48,49,50,51,52,53,54], and non-enzymic antioxidants: ascorbic Acid, GSH, α-Tocopherol, Ubiquinone, Thioredoxin (TRX) and Bilirubin [48,49,50,51,52,53,54,55,56,57,58] (Table 5). In general, the activities/concentrations of these antioxidants tend to decrease in NAFLD/NASH patients with a few exceptions where they increased. These variations appear to be sample dependent: for instance, those determined in the liver samples of patient generally showed a decrease in their activities/levels in all the data reviewed. However, there are variations with respect to other samples such as blood, plasma and serum, where antioxidant levels tend to increase in most cases of NAFLD and NASH.
6.2. Oxidative Stress Markers Measured in Clinical NAFLD/NASH
Oxidative stress biomarkers that have been determined in clinical models of NAFLD include nitric oxide (NO), lipid damage products (lipid peroxides, TBARS (MDA), Hydroperoxides, 8-Isoprostane, 4-HNE), protein oxidation products (protein carbonyl, Nitrotyrosine), DNA oxidation product (8-OH-dG) and CYP2E1 (Table 6). The concentrations/activities of these biomarkers increase generally in all the clinical data reviewed, except where the increase is not significant [52] (Table 6). The most frequently reported OS markers include the NO, MDA, 8-OH-dG and CYP2E1.
6.3. Antioxidant Markers Measured in Experimental Models of NAFLD/NASH
Extensive data on antioxidants used to assess NAFLD/NASH in experimental models have been reviewed. They mainly cover the enzymic antioxidants: SOD, CAT, GPx, GR and the non-enzymic antioxidant—GSH in about five rodent models. In nearly all the experimental data reviewed, these antioxidants were measured in the liver and their activities decreased generally in most of the models (mainly NASH), except in a few cases as shown in Table 7.
6.4. Oxidative Stress Markers Measured in Experimental Models of NAFLD/NASH
Oxidative stress biomarkers measured at experimental level of NAFLD/NASH include the reactive species (H2O2, nitrite/nitrate), lipid oxidation products (MDA, Lipid peroxide, 8-Isoprostanes, 4-HNE), protein damage products (Protein carbonyl, Dityrosine, Hydroxyproline, Nitrotyrosine), DNA damage product (8-OH-dG), and oxidative enzymes (CYP2E1, NADPH Oxidase, Xanthine Oxidase), as summarized in Table 8.
The levels or activities of these biomarkers are found to consistently increase in all of the experimental models, except where the increases were not significant. In all the experimental models, MDA is the most measured oxidative stress biomarker as presented in Table 8. Next to MDA is 4-HNE (another product of lipid oxidation), 8-OH-dG and CYP2E1. Others like H2O2, nitrite/nitrate, lipid peroxide, 8-Isoprostanes, Protein carbonyl, Dityrosine, Hydroxyproline, Nitrotyrosine), NADPH Oxidase and Xanthine Oxidase are not frequently measured. However, their levels/activities increase in a similar pattern to MDA, 8-OH-dG, 4-HNE, CYP2E1.
7. Conclusions
Oxidative stress plays an important role in the pathophysiology of NAFLD/NASH. Several markers of oxidative stress and antioxidants have been shown to be very useful in assessing the redox state in NAFLD/NASH. Among the oxidative stress biomarkers reviewed, TBARS, MDA, CYP2E1 and 4-HNE are unique; they are represented in both clinical and experimental measurements and they consistently increase. Antioxidants of interest in clinical and pre-clinical assessment of NAFLD/NASH include GSH, SOD, CAT, and GPx, which appear to be most reliably detected in liver samples.
Author Contributions
Writing-Original Draft Preparation, A.O.; Writing-Review & Editing, O.A.A.
Funding
This review received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Angulo, P. Non-alcoholic fatty liver disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Manne, V.; Handa, P.; Kowdley, K.V. Pathophysiology of Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis. Clin. Liver Dis. 2018, 22, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, J.; Viggiano, T.R.; McGill, D.B.; Oh, B.J. Non-alcoholic steatohepatitis: Mayo clinic experiences with a hitherto unnamed disease. Mayo Clin. Proc. 1980, 55, 434–438. [Google Scholar] [PubMed]
- Kim, W.R.; Lake, J.R.; Smith, J.M.; Skeans, M.A.; Schladt, D.P.; Edwards, E.B.; Harper, A.M.; Wainright, J.L.; Snyder, J.J.; Israni, A.K.; et al. OPTN/SRTR 2015 Annual Data Report: Liver. Am. J. Transplant. 2017, 17, 174–251. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of Liver (EASL). Background Media Information: Fast facts about liver disease. Int. Liver Congr. 2016, 1–5. [Google Scholar]
- Day, C.P.; James, O. Steatohepatitis: A tale of two ‘hits’? Gastroenterol. 1998, 114, 842–845. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of nonalcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Anstee, Q.M.; Valenti, L. Genetic Predisposition in NAFLD and NASH: Impact on Severity of Liver Disease and Response to Treatment. Curr. Pharm. Des. 2013, 19, 5219–5238. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.C. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Sies, H. On the history of oxidative stress: Concept and some aspects of current development. Curr. Opin. Toxicol. 2018, 7, 122–126. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224C, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hong, M.; Tan, H.; Wang, N.; Feng, Y. Insights into the Role and Interdependence of Oxidative Stress and Inflammation in Liver Diseases. Oxid. Med. Cell. Longev. 2016, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.; Wang, N.; Zhang, Z.; Lao, L.; Wong, C.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef] [PubMed]
- Muriel, P. Role of free radicals in liver diseases. Hepatol. Int. 2009, 3, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Barzilai, A.; Yamamoto, K. DNA damage responses to oxidative stress. Dna Repair 2004, 3, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- d’Ischia, M.; Manini, P.; Napolitano, A. Oxidative Damage to Carbohydrates and Amino Acids. In Oxidative Stress, Disease and Cancer; Word Scientific: Singapore, 2006; pp. 333–356. [Google Scholar] [CrossRef]
- Hassan, H.A.; El-Aal, M.A. Lipid Peroxidation End-Products as a Key of Oxidative Stress: Effect of Antioxidant on Their Production and Transfer of Free Radicals. In Lipid Peroxidation; Catala, A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 63–88. [Google Scholar] [CrossRef]
- Sitte, N. Oxidative Damage to Proteins. In Aging at the Molecular Level; Zglinicki, T., Ed.; Springer-Science: Newcastle, UK, 2003. [Google Scholar] [CrossRef]
- Gackowski, D.; Speina, E.; Zielinska, M.; Kowalewski, J.; Rozalski, R.; Siomek, A.; Paciorek, T.; Tudek, B.; Olinski, R. Products of oxidative DNA damage and repair as possible biomarkers of susceptibility to lung cancer. Cancer Res. 2003, 15, 4899–4902. [Google Scholar]
- Nishida, N.; Arizumi, T.; Takita, M.; Kitai, S.; Yada, N.; Hagiwara, S.; Inoue, T.; Minami, Y.; Ueshima, K.; Sakurai, T.; et al. Reactive oxygen species induce epigenetic instability through the formation of 8-hydroxydeoxyguanosine in human hepatocarcinogenesis. Dig. Dis. 2013, 31, 459–466. [Google Scholar] [CrossRef]
- Simioni, C.; Zauli, G.; Martelli, A.M.; Vitale, M.; Sacchetti, G.; Gonelli, A.; Neri, L.M. Oxidative stress: Role of physical exercise and antioxidant nutraceuticals in adulthood and aging. Oncotarget 2018, 9, 17181–17198. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncola, J.; Cronin, M.T.D.; Mazura, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Koek, G.H.; Liedorp, P.R.; Bast, A. The role of oxidative stress in non-alcoholic steatohepatitis. Clin. Chim. Acta 2011, 412, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Tariq, Z.; Green, C.J.; Hodson, L. Are oxidative stress mechanisms the common denominator in the progression from hepatic steatosis towards non-alcoholic steatohepatitis (NASH)? Liver Int. 2014, 34, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Serviddio, G.; Sastre, J.; Bellanti, F.; Viña, J.; Vendemiale, G.; Altomare, E. Mitochondrial involvement in non-alcoholic steatohepatitis. Mol. Asp. Med. 2008, 29, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Vendemiale, G.; Grattagliano, I.; Caraceni, P.; Caraccio, G.; Domenicali, M.; Dall’Agata, M.; Trevisani, F.; Guerrieri, F.; Bernardi, M.; Altomare, E. Mitochondrial oxidative injury and energy metabolism alteration in rat fatty liver: Effect of the nutritional status. Hepatology 2001, 33, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Satapati, S.; Kucejova, B.; Duarte, J.A.; Fletcher, J.A.; Reynolds, L.; Sunny, N.E.; He, T.; Nair, L.A.; Livingston, K.A.; Fu, X.; et al. Mitochondrial metabolism mediates oxidative stress and inflammation in fatty liver. J. Clin. Investig. 2015, 125, 4447–4462. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.; Sanya, A.J. The metabolic abnormalities associated with non-alcoholic fatty liver disease. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 709–731. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P.; Lindor, K.D. Insulin resistance and mitochondrial abnormalities in NASH: A cool look into a burning issue. Gastroenterology 2001, 120, 1281–1285. [Google Scholar] [CrossRef]
- Gonzalez, F.J.; Peters, J.M.; Cattley, R.C. Mechanism of action of the nongenotoxic peroxisome proliferators: Role of the peroxisome proliferator-activator receptor alpha. J. Natl. Cancer Inst. 1998, 90, 1702–1709. [Google Scholar] [CrossRef]
- Pessayre, D.; Mansouri, A.; Fromenty, B. Nonalcoholic steatosis and steatohepatitis. V. Mitochondrial dysfunction in steatohepatitis. Am. J. Physiol. Gastrointest. Liv. Physiol. 2002, 282, 193–199. [Google Scholar] [CrossRef]
- Caldwell, S.H.; Swerdlow, R.H.; Khan, E.M.; Iezzoni, J.C.; Hespenheide, E.E.; Parks, J.K.; Parker, W.D. Mitochondrial abnormalities in non-alcoholic steatohepatitis. J. Hepatol. 1999, 31, 430–434. [Google Scholar] [CrossRef]
- Weltman, M.D.; Farrell, G.C.; Hall, P.; Ingelman-Sundberg, M.; Liddle, C. Hepatic cytochrome P450 2E1 is increased in patients with nonalcoholic steatohepatitis. Hepatology 1998, 27, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Gorski, J.C.; Asghar, M.S.; Asghar, A.; Foresman, B.; Hall, S.D.; Crabb, D.W. Hepatic cytochrome P450 2E1 activity in nondiabetic patients with nonalcoholic steatohepatitis. Hepatology 2003, 37, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Matsunami, T.; Sato, Y.; Ariga, S.; Sato, T.; Kashimura, H.; Hasegawa, Y.; Yukawa, M. Regulation of oxidative stress and inflammation by hepatic adiponectin receptor 2 in an animal model of nonalcoholic steatohepatitis. Int. J. Clin. Exp. Pathol. 2010, 22, 472–481. [Google Scholar]
- Leclercq, A.; Farrell, G.C.; Field, J.; Bell, D.R.; Gonzalez, F.J.; Robertson, G.R. CYP2E1 and CYP4A as microsomal catalysts of lipid peroxides in murine nonalcoholic steatohepatitis. J. Clin. Investig. 2000, 105, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jhaveri, R.; Huang, J. Endoplasmic reticulum stress, hepatocyte CD1 and NKT cell abnormalities in murine fatty livers. Lab. Investig. 2007, 87, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, S.; Negoro, M.; Kitamura, T.; Fujimoto, Y. Xanthine oxidase-derived reactive oxygen species mediate 4-oxo-2-nonenal-induced hepatocyte cell death. Toxicol. Appl. Pharmacol. 2010, 249, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Baskol, G.; Baskol, M.; Kocer, D. Oxidative stress and antioxidant defenses in serum of patients with non-alcoholic steatohepatitis. Clin. Biochem. 2007, 40, 776–780. [Google Scholar] [CrossRef]
- Crespo, J.; Cayon, A.; Fernandez-Gil, P.; Hernández-Guerra, M.; Mayorga, M.; Domínguez-Díez, A.; Fernández-Escalante, J.C.; Pons-Romero, F. Gene expression of tumor necrosis factor alpha and TNF-receptors, p55 and p75, in nonalcoholic steatohepatitis patients. Hepatology 2001, 34, 1158–1163. [Google Scholar] [CrossRef]
- Imajo, K.; Fujita, K.; Yoneda, M.; Nozaki, Y.; Ogawa, Y.; Shinohara, Y.; Kato, S.; Mawatari, H.; Shibata, W.; Kitani, H.; et al. Hyperresponsivity to low-dose endotoxin during progression to nonalcoholic steatohepatitis is regulated by leptin-mediated signaling. Cell Metab. 2012, 16, 44–54. [Google Scholar] [CrossRef]
- Dostert, K.; Dorn, C.; Weiss, T.; Gabele, E.; Hellerbrand, C. Enhanced TLR4 expression in steatotic hepatocytes. Z. Gastroenterol. 2012, 50, 1–11. [Google Scholar] [CrossRef]
- Ucar, F.; Sezer, S.; Erdogan, S.; Akyol, S.; Armutcu, F.; Akyol, O. The relationship between oxidative stress and nonalcoholic fatty liver disease: Its effects on the development of nonalcoholic steatohepatitis. Redox Rep. 2013, 18, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Gambino, R.; Musso, G.; Cassader, M. Redox Balance in the Pathogenesis of Nonalcoholic Fatty Liver Disease: Mechanisms and Therapeutic Opportunities. Antioxid. Redox Signal. 2011, 15, 1325–1365. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Stiuso, P.; Scognamiglio, I.; Murolo, M.; Ferranti, P.; De Simone, C.; Rizzo, M.R.; Tuccillo, C.; Caraglia, M.; Loguercio, C.; Federico, A. Serum Oxidative Stress Markers and Lipidomic Profile to Detect NASH Patients Responsive to an Antioxidant Treatment: A Pilot Study. Oxidat. Med. Cell. Longev. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Leghi, G.E.; Domenici, F.A.; Vannucchi, H. Influence of oxidative stress and obesity in patients with nonalcoholic steatohepatitis. ARQ Gastroenterol. 2015, 52, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Koruk, M.; Taysi, S.; Savas, M.C.; Yilmaz, O.; Akcay, F.; Karakok, M. Oxidative stress and enzymatic antioxidant status in patients with nonalcoholic steatohepatitis. Ann. Clin. Lab. Sci. 2004, 34, 57–62. [Google Scholar] [PubMed]
- Köroğlu, E.; Canbakan, B.; Atay, K.; Hatemi, I.; Tuncer, M.; Dobrucalı, A.; Sonsuz, A.; Gültepe, I.; Şentürk, H. Role of oxidative stress and insulin resistance in disease severity of non-alcoholic fatty liver disease. Turk. J. Gastroenterol. 2016, 27, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Yesilova, Z.; Yaman, H.; Oktenli, C.; Ozcan, A.; Uygun, A.; Cakir, E.; Sanisoglu, S.Y.; Erdil, A.; Ates, Y.; Aslan, M.; et al. Systemic markers of lipid peroxidation and antioxidants in patients with nonalcoholic Fatty liver disease. Am. J. Gastroenterol. 2005, 100, 850–855. [Google Scholar] [CrossRef]
- Ergün, Y.; Kurutaş, E.B.; Özdil, B.; Güneşaçar, R.; Ergün, Y. Nitric Oxide and Nitrotyrosine Levels in Relation with Oxidative Stress-Related Markers in Non-Alcoholic Fatty Liver Disease. Turk. Klin. J. Gastroenterohepatol 2011, 18, 1–8. [Google Scholar]
- Videla, L.A.; Rodrigo, R.; Orellana, M.; Fernandez, V.; Tapia, G.; Quiñones, L.; Varela, N.; Contreras, J.; Lazarte, R.; Csendes, A.; et al. Oxidative stress-related parameters in the liver of non-alcoholic fatty liver disease patients. Clin. Sci. 2004, 106, 261–268. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, A.; Duseja, A.; Das, A.; Dhiman, R.K.; Chawla, Y.K.; Kohli, K.K.; Bhansali, A. Patients with Nonalcoholic Fatty Liver Disease (NAFLD) have Higher Oxidative Stress in Comparison to Chronic Viral Hepatitis. J. Clin. Exp. Hepatol. 2013, 3, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Nakashima, T.; Yoh, T.; Furutani, M.; Hirohama, A.; Kakisaka, Y.; Nakajima, Y.; Ishikawa, H.; Mitsuyoshi, H.; Okanoue, T.; et al. Serum thioredoxin levels as a predictor of steatohepatitis in patients with nonalcoholic fatty liver disease. J. Hepatol. 2003, 38, 32–38. [Google Scholar] [CrossRef]
- Salomone, F.; Volti, G.L.; Rosso, C.; Grosso, G.; Bugianesi, E. Unconjugated bilirubin, a potent endogenous antioxidant, is decreased in patients with nonalcoholic steatohepatitis and advanced fibrosis. J. Gastroenterol. Hepatol. 2013, 28, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Hjelkrem, M.; Morales, A.; Williams, C.D.; Harrison, S.A. Unconjugated hyperbilirubinemia is inversely associated with non-alcoholic steatohepatitis (NASH). Aliment. Pharmacol. Ther. 2012, 35, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Rastogi, A.; Maras, J.S.; Sarin, S.K. Unconjugated hyperbilirubinemia in patients with non-alcoholic fatty liver disease: A favorable endogenous response. Clin. Biochem. 2012, 45, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Horoz, M.; Bolukbas, C.; Bolukbas, F.F.; Sabuncu, T.; Aslan, M.; Sarifakiogullari, S.; Gunaydin, N.; Erel, O. Measurement of the total antioxidant response using a novel automated method in subjects with nonalcoholic steatohepatitis. BMC Gastroenterol. 2005, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Farhan, A.R. Evaluation of Melatonin Hormone and Nitric oxide Levels in Non-alcoholic Fatty Liver Patients in Relation to Obesity and Oxidative Stress. J. Pharm. Sci. Res. 2018, 10, 1167–1169. [Google Scholar]
- Loguercio, C.; De Girolamo, V.; de Sio, I.; Tuccillo, C.; Ascione, A.; Baldi, F.; Budillon, G.; Cimino, L.; Di Carlo, A.; Di Marino, M.P.; et al. Non-alcoholic fatty liver disease in an area of southern Italy: Main clinical, histological, and pathophysiological aspects. J. Hepatol. 2001, 35, 568–574. [Google Scholar] [CrossRef]
- Oliveira, C.P.; Faintuch, J.; Rascovski, A.; Furuya, C.K., Jr.; Bastos, M.S.; Matsuda, M.; Della Nina, B.I.; Yahnosi, K.; Abdala, D.S.; Vezozzo, D.C.; et al. Lipid peroxidation in bariatric candidates with nonalcoholic fatty liver disease (NAFLD)—Preliminary findings. Obes. Surg. 2005, 15, 502–505. [Google Scholar] [CrossRef]
- Seki, S.; Kitada, T.; Yamada, T.; Sakaguchi, H.; Nakatani, K.; Wakasa, K. In situ detection of lipid peroxidation and oxidative DNA damage in non-alcoholic fatty liver diseases. J. Hepatol. 2002, 37, 56–62. [Google Scholar] [CrossRef]
- Fujita, N.; Miyachi, H.; Tanaka, H.; Takeo, M.; Nakagawa, N.; Kobayashi, Y.; Iwasa, M.; Watanabe, S.; Takei, Y. Iron overload is associated with hepatic oxidative damage to DNA in nonalcoholic steatohepatitis. Cancer Epidemiol. Biomark. Prev. 2009, 18, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Nobili, V.; Parola, M.; Alisi, A.; Marra, F.; Piemonte, F.; Mombello, C.; Sutti, S.; Povero, D.; Maina, V.; Novo, E.; et al. Oxidative stress parameters in paediatric non-alcoholic fatty liver disease. Int. J. Mol. Med. 2010, 26, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Chtioui, H.; Semela, D.; Ledermann, M.; Zimmermann, A.; Dufour, J.F. Expression and activity of the cytochrome P450 2E1 in patients with non-alcoholic steatosis and steatohepatitis. Liver Int. 2007, 27, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, S.; Hamada, A.; Jobu, K.; Yokota, J.; Onogawa, M.; Kyotani, S.; Miyamura, M.; Saibara, T.; Onishi, S.; Nishioka, Y. Effects of Eriobotrya japonica seed extract on oxidative stress in rats with non-alcoholic steatohepatitis. J. Pharm. Pharmacol. 2010, 62, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Marcolin, E.; Forgiarini, L.F.; Rodrigues, G.; Tieppo, J.; Borghetti, G.S.; Bassani, V.L.; Picada, J.N.; Marroni, N.P. Quercetin Decreases Liver Damage in Mice with Non-Alcoholic Steatohepatitis. Basic Clin. Pharmacol. Toxicol. 2013, 112, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Marcolin, E.; Forgiarini, L.F.; Tieppo, J.; Dias, A.S.; Freitas, L.A.R.; Marroni, N.P. Methione- and choline-deficient diet induces hepatic changes characteristic of non-alcoholic steatohepatitis. Arq. Gastroenterol. 2011, 48, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.M.; Wu, W.J.; Fu, N.; Liang, B.L.; Wang, R.Q.; Li, L.X.; Zhao, S.X.; Zhao, J.M.; Yu, J. Antioxidants vitamin E and 1-aminobenzotriazole prevent experimental non-alcoholic steatohepatitis in mice. Scand. J. Gastroenterol. 2009, 44, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, N.; Aghazadeh, S.; Yazdanparast, R. Effects of Teucrium polium on Insulin Resistance in Nonalcoholic Steatohepatitis. J. Acupunct. Meridian Stud. 2010, 3, 104–110. [Google Scholar] [CrossRef]
- Huang, F.; Wang, J.; Yu, F.; Tang, Y.; Ding, G.; Yang, Z.; Sun, Y. Protective Effect of Meretrix meretrix Oligopeptides on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. Mar. Drugs 2018, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lu, Y. Hepatoprotective Effects of Sophoricoside against Fructose-Induced Liver Injury via Regulating Lipid Metabolism, Oxidation, and Inflammation in Mice. J. Food Sci. 2018, 83, 552–558. [Google Scholar] [CrossRef]
- Song, L.; Qu, D.; Zhang, Q.; Jiang, J.; Zhou, H.; Jiang, R.; Li, Y.; Zhang, Y.; Yan, H. Phytosterol esters attenuate hepatic steatosis in rats with nonalcoholic fatty liver disease rats fed a high-fat diet. Sci. Rep. 2017, 7, 41604. [Google Scholar] [CrossRef] [PubMed]
- Korish, A.A.; Arafah, M.M. Camel milk ameliorates steatohepatitis, insulin resistance and lipid peroxidation in experimental non-alcoholic fatty liver disease. BMC Complement. Altern. Med. 2013, 13, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Elshazly, S.M. Ameliorative effect of nicorandil on high fat diet induced non-alcoholic fatty liver disease in rats. Eur. J. Pharmacol. 2015, 748, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Janevski, M.; Antonas, K.N.; Sullivan-Gunn, M.J.; McGlynn, M.A.; Lewandowski, P.A. The effect of cocoa supplementation on hepatic steatosis, reactive oxygen species and LFABP in a rat model of NASH. Comp. Hepatol. 2011, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Caballero, F.; Fernández, A.; Matías, N.; Martínez, L.; Fucho, R.; Elena, M.; Caballeria, J.; Morales, A.; Fernández-Checa, J.C.; García-Ruiz, C. Specific Contribution of Methionine and Choline in Nutritional Nonalcoholic Steatohepatitis. J. Biol. Chem. 2010, 285, 18528–18536. [Google Scholar] [CrossRef] [PubMed]
- Schröder, T.; Kucharczyk, D.; Bär, F.; Page, R.; Derer, S.; Jendrek, T.; Sünderhauf, A.; Brethack, A.K.; Hirose, M.; Möller, S.; et al. Mitochondrial gene polymorphisms alter hepatic cellular energy metabolism and aggravate diet-induced non-alcoholic steatohepatitis. Mol. Metab. 2016, 5, 283–295. [Google Scholar] [CrossRef]
- Sutti, S.; Jindal, A.; Locatelli, I.; Vacchiano, M.; Gigliotti, L.; Bozzola, C.; Albano, E. Adaptive Immune Responses Triggered by Oxidative Stress Contribute to Hepatic Inflammation in NASH. Hepatology 2014, 59, 886–897. [Google Scholar] [CrossRef]
- Nazmy, M.H.; Abdel-Ghany, M.I. Serum markers versus histopathological scoring for discrimination between experimental fatty liver and non-alcoholic steatohepatitis. Int. Res. J. Med. Med. Sci. 2015, 3, 51–59. [Google Scholar]
- Sugatani, J.; Wada, T.; Osabe, M.; Yamakawa, K.; Yoshinari, K.; Miwa, M. Dietary Inulin Alleviates Hepatic Steatosis and Xenobiotics-Induced Liver Injury in Rats Fed a High-Fat and High-Sucrose Diet: Association with the Suppression of Hepatic Cytochrome P450 and Hepatocyte Nuclear Factor 4α Expression. Drug Metab. Dispos. 2006, 34, 1677–1687. [Google Scholar] [CrossRef]
- Takahashi, Y.; Sugimoto, K.; Soejima, Y.; Kumagai, A.; Koeda, T.; Shojo, A.; Nakagawa, K.; Harada, N.; Yamaji, R.; Inui, H.; et al. Inhibitory Effects of Eucalyptus and Banaba Leaf Extracts on Nonalcoholic Steatohepatitis Induced by a High-Fructose/High-Glucose Diet in Rats. Biomed. Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Watanabe, M.; Fuda, H.; Okabe, H.; Joko, S.; Miura, Y.; Hui, S.; Hamaoka, Y.N.; Miki, E.; Chiba, H. Oyster extracts attenuate pathological changes in non-alcoholic steatohepatitis (NASH) mouse model. J. Funct. Foods 2016, 20, 516–531. [Google Scholar] [CrossRef]
- Ji, G.; Wang, Y.; Deng, Y.; Li, X.; Jiangm, Z. Resveratrol ameliorates hepatic steatosis and inflammation in methionine/choline deficient diet-induced steatohepatitis through regulating autophagy. Lipids Health Dis. 2015, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Sakaida, I.; Okita, K. The role of oxidative stress in NASH and fatty liver model. Hepatol. Res. 2005, 33, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, K.; Takayama, F.; Mankura, M.; Hidaka, Y.; Egashira, T.; Ogino, T.; Kawasaki, H.; Morim, A. Beneficial effects of fermented green tea extract in a rat model of non-alcoholic steatohepatitis. J. Clin. Biochem. Nutr. 2009, 44, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S.; Leo, M.A.; Mak, K.M.; Xu, Y.; Cao, Q.; Ren, C.; Ponomarenko, A.; DeCarli, L.M. Model of nonalcoholic steatohepatitis. Am. J. Clin. Nutr. 2004, 79, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.Y.; Park, H.J.; Manautou, J.E.; Koo, S.I.; Bruno, R.S. Green tea extract protects against nonalcoholic steatohepatitis in ob/ob mice by decreasing oxidative and nitrative stress responses induced by proinflammatory enzymes. J. Nutr. Biochem. 2012, 23, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, J.; Spincemaille, P.; Vanhorebeek, I.; Van den Berghe, G.; Vander Elst, I.; Windmolders, P.; van Pelt, J.; van der Merwe, S.; Bedossa, P.; Nevens, F.; et al. Dietary intervention, but not losartan, completely reverses non-alcoholic steatohepatitis in obese and insulin resistant mice. Lipids Health Dis. 2017, 23, 46–56. [Google Scholar] [CrossRef]
- Lee, S.J.; Kang, J.H.; Iqbal, W.; Kwon, O.S. Proteomic Analysis of Mice Fed Methionine and Choline Deficient Diet Reveals Marker Proteins Associated with Steatohepatitis. PLoS ONE 2015, 10, e0120577. [Google Scholar] [CrossRef]
- Jung, Y.A.; Choi, Y.K.; Jung, G.S.; Seo, H.Y.; Kim, H.S.; Jang, B.K.; Kim, J.G.; Lee, I.K.; Kim, M.K.; Park, K.G. Sitagliptin attenuates methionine/choline-deficient diet-induced steatohepatitis. Diabetes Res. Clin. Pract. 2014, 105, 47–57. [Google Scholar] [CrossRef]
- Kathirve, E.; Chen, P.; Morgan, K.; French, S.W.; Morgan, T.R. Oxidative stress and regulation of anti-oxidant enzymes in cytochrome P4502E1 transgenic mouse model of non-alcoholic fatty liver. J. Gastroenterol. Hepatol. 2010, 25, 1136–1143. [Google Scholar] [CrossRef]
- Lieber, C.S.; Leo, M.A.; Mak, K.M.; Xu, Y.; Cao, Q.; Ren, C.; Ponomarenko, A.; DeCarli, L.M. Acarbose attenuates experimental non-alcoholic steatohepatitis. Biochem. Biophys. Res. Commun. 2004, 12, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ausman, L.M.; Greenberg, A.S.; Russell, R.M.; Wang, X. Dietary lycopene and tomato extract supplementations inhibit nonalcoholic steatohepatitis-promoted hepatocarcinogenesis in rats. Int. J. Cancer 2010, 126, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Takayama, F.; Egashira, T.; Imao, M.; Mori, A. Fermented papaya preparation halts the progression of non-alcoholic steatohepatitis in rats. J. Biophys. Chem. 2013, 4, 84–90. [Google Scholar] [CrossRef]
Figure 1.
Pathophysiology of NASH and the multi-hit hypothesis.


{kind=link}
Table 1.
Major ROS and RNS of physiological importance.
| Major ROS | Sources | |
| Free Radicals | Hydroxyl radical () | Decomposition of ONOO− or, HOCl |
| Superoxide radical () | Electron transport systems, and one-electron reduction of O2 by respiratory burst via the action of membrane bound NADPH oxidase | |
| Peroxyl radical (ROO•) | Produced in the Fenton reaction | |
| Non-Radicals | Hydrogen peroxide (H2O2) | Activated macrophages during inflammation |
| Hypochlorous acid (HClO) | Combined activities of NADPH oxidase and myeloperoxidase (MPO) in phagocytes | |
| Lipid peroxides (ROOH) | Formed from oxidation of polyunsaturated fatty acid via lipid-peroxyl radical reaction | |
| Major RNS | Sources | |
| Free Radicals | Nitric oxide (NO•) | Nitric Oxide Synthase (NOS) |
| Nitrogen dioxide () | Activated neutrophils | |
| Non-radicals | Dinitrogen trioxide (N2O3) | Produced in pathological conditions where (inducible nitric oxide (iNOS) is upregulated |
| Peroxinitrite (ONOO−) | Produced in pathological conditions where iNOS is upregulated | |
| Nitrite () | Oxidation product from NO, formed during NOS activation in inflammatory diseases | |
| Nitryl ion () | Activated neutrophils | |
Table 2.
Some important oxidative damage products.
| Macromolecule | Oxidative Damage | Damage Products |
|---|---|---|
| Lipids | Lipid oxidation/peroxidation | Malondialdehyde (MDA) Lipid peroxide, (lipid endoperoxides and lipid hydroperoxides) 8-Isoprostane 4-hydroxy-2-nonenal (4-HNE) |
| Proteins | Protein Oxidation, protein cross linkage, Oxidative modification of amino acids | Protein carbonyl compounds, 3-Nitrotyrosine (product of ROS-mediated nitration of tyrosine), 2-oxohistidine, hydroxyproline etc. |
| DNA | RNA/DNA fragmentation (single and double-strand breaks) Modification of base, sugar | 8-hydroxy-2′-deoxyguanosine (8-OH-dG), 8-hydroxyguanine (8-OH-G) |
Table 3.
Physiologically important antioxidants and their functions.
| Antioxidant Type | Antioxidant Name | Functions |
|---|---|---|
| Enzymic antioxidants | Superoxide dismutase (SOD) | Converts to H2O2 and O2 |
| Catalase (CAT) | Converts H2O2 to 2H2O and O2 | |
| Glutathione peroxidase (GPx) | Detoxifies H2O2 and Lipid peroxides using reduced glutathione (GSH) producing the oxidised form of glutathione (GSSG) | |
| Glutathione reductase (GR) | Reduces GSSG to GSH | |
| Non-enzymic antioxidants | Ascorbic Acid (AA) | Detoxifies Superoxide radical, Hydroxyl radical, and H2O2 |
| Reduced Glutathione (GSH) | Neutralizes Superoxide radical, Hydroxyl radical, and H2O2; co-substrate for glutathione peroxidase | |
| α-Tocopherol | Detoxifies H2O2; protects against membrane lipid peroxidation (LPO) | |
| Ubiquinone | Detoxifies Lipid peroxides | |
| Thioredoxin (TRX) | General thiol redox control of protein activity via reversible disulfide formation | |
| Bilirubin | Effective in quenching/scavenging secondary oxidants produced during OS |
Table 4.
Sources of oxidative stress in NAFLD/NASH.
| Source | Contribution to OS in NAFLD/NASH | References |
|---|---|---|
| Mitochondrial metabolism (β-oxidation) | Production of ROS as a result of electron leakage during mitochondrial β-oxidation | [26,27,28,29,30] |
| Peroxisomal β-oxidation | Generation of H2O2 during peroxisomal β-oxidation which is converted into hydroxyl radical contributing to OS | [31] |
| Mitochondrial electron transport chain | inhibition of electron transport chain by TNF-α and lipid peroxidation products | [32,33] |
| Microsomal Cytochrome P450 enzymes | ROS generation due to Increase in activity of cytochrome P4502E1 (CYP2E1) involved in lipooxygenation of longchain fatty acids | [34,35,36,37] |
| Endoplasmic reticulum (ER) stress | Endoplasmic reticulum stress response, promotes OS via increased expression of CHOP (also called DDIT –DNA Damage Inducible Transcript-3 protein) | [38] |
| Xanthine Oxidase (XO) | Increase in XO activity generates superoxide anions, due to induction by 4-HNE (a product of lipid peroxidation) | [39,40] |
| Inflammatory Response | Abnormal inflammatory response mediated by gut microflora resulting in increase in pro-oxidants | [41,42,43] |
Table 5.
Antioxidant markers measured in clinical NAFLD/NASH.
| Antioxidant Marker | Sample | Level/Activity/Expression in Sample | Clinical Case | Reference(s) |
|---|---|---|---|---|
| SOD | Serum | Decreased, Increased (nsc) | NASH | [47,48,49] |
| Serum/Liver | Decreased | NAFLD | [50] | |
| Plasma | Decreased | NAFLD | [51] | |
| Blood | Increased | NAFLD | [52] | |
| Liver | Decreased | NAFLD | [53] | |
| Blood | Increased | NAFLD | [54] | |
| CAT | Serum | Decreased, Increased (nsc) | NASH | [47,48] |
| Plasma | Decreased | NAFLD | [51] | |
| Blood | Increased | NAFLD | [52] | |
| Liver | Decreased | NAFLD | [53] | |
| Blood | Decreased | NAFLD | [54] | |
| GPx | Serum | Decreased | NASH | [48,49] |
| Blood | Increased | NAFLD | [54] | |
| Liver | Decreased | NAFLD | [53] | |
| GR | Serum | Increased | NASH | [49] |
| Blood | Increased | NAFLD | [54] | |
| Ascorbic Acid | Serum | nsc | NASH | [48] |
| Serum | Decreased | NAFLD | [50] | |
| GSH | Serum | Increased | NASH | [49] |
| Blood | Increased | NAFLD | [52] | |
| Liver | Decreased | NAFLD | [50,53] | |
| Blood | Decreased | NAFLD | [54] | |
| α-Tocopherol | Serum | Increased | NASH | [48] |
| Serum | Decreased | NAFLD | [50] | |
| Ubiquinone | Serum | Decreased | NAFLD | [51] |
| Thioredoxin (TRX) | Serum | Increased | NAFLD | [55] |
| Bilirubin | Serum | Decreased | NASH | [56,57] |
| Serum | Decreased | NAFLD | [58] |
nsc: no significant change.
Table 6.
Oxidative stress markers measured in clinical NAFLD/NASH.
| OS Marker | Sample | Level/Activity/Expression in Sample | Clinical Case | Reference(s) |
|---|---|---|---|---|
| Lipid peroxides | Plasma | Increased | NASH | [59] |
| NO• | Serum | Increased | NASH | [47,49] |
| Serum | Increased | NAFLD | [50,60] | |
| Blood | Increased | NAFLD | [52] | |
| TBARS/MDA | Serum | Increased | NAFLD | [47] |
| Serum | Increased | NAFLD/NASH | [49,51,61] | |
| Serum | Increased | NAFLD | [50] | |
| Blood | Increased | NAFLD | [54] | |
| Hydroperoxides | liver | Increased | NASH | [62] |
| 8-Isoprostane | Plasma | Increased | NASH | [48] |
| 4-HNE | Liver | Increased | NASH | [63] |
| Protein carbonyl | Liver | Increased | NAFLD | [53] |
| Nitrotyrosine | Blood | nsc | NAFLD | [52] |
| 8-OH-dG | Liver | Increased | NASH | [63,64] |
| Liver | Increased | NAFLD | [65] | |
| CYP2E1 | Liver | Increased | NASH | [34,35] |
| Liver | nsc | Steatosis/NASH | [66] | |
| Liver | Increased | NAFLD | [53] |
nsc: no significant change.
Table 7.
Antioxidant markers measured in experimental NAFLD/NASH.
| Antioxidant Marker | Sample | Level/Activity/Expression in Sample | Experimental Model | Experimental Specie | Reference(s) |
|---|---|---|---|---|---|
| SOD | Liver | Decreased | NASH (MCD) | Wistar Rats | [67] |
| Liver | Increased | NASH (MCD) | C57BL/6 Mice | [68] | |
| Liver | Increased | NASH (MCD) | C57BL/6 mice | [69] | |
| Liver | Decreased | NASH (MCD) | C57BL6/J mice | [70] | |
| Liver | Decreased | NASH (MCD) | N-Mary rats | [71] | |
| Liver | Decreased | NAFLD (HFD) | Mice | [72] | |
| Liver | Decreased | NASH (HF) | Kunming mice | [73] | |
| Liver | Increased | NAFLD (HFD) | Rat | [74] | |
| CAT | Liver | Decreased | NASH (MCD) | Wistar Rats | [67] |
| Liver | Decreased | NASH (MCD) | C57BL/6 Mice | [68] | |
| Liver | Increased | NASH (MCD) | C57BL/6 mice | [69] | |
| Liver | Decreased | NASH (HCD) | Wistar Rats | [75] | |
| Liver | Decreased | NAFLD (HFD) | Sprague-Dawley rats | [74] | |
| GPx | Liver | Decreased | NASH (MCD) | Wistar Rats | [67] |
| Liver | Increase | NASH (MCD) | N-Mary rats | [71] | |
| Liver | Decreased | NAFLD (HFD) | Mice | [72] | |
| Liver | Decreased | NASH (HF) | Kunming mice | [73] | |
| Liver | nsc | NAFLD (HFD) | Sprague-Dawley rats | [74] | |
| GR | Liver | Decreased | NASH (MCD) | N-Mary rats | [71] |
| GSH | Liver | Decreased | NASH (MCD) | Wistar Rat | [67] |
| Liver | Decreased | NASH (MCD) | N-Mary rats | [71] | |
| Liver | Decreased | NAFLD (HFD) | Wistar Rats | [76] | |
| Liver | Decreased | NAFLD (HCD) | Wistar Rats | [75] | |
| Liver/RBC | Increased | NASH (HF-MCD) | Sprague-Dawley rats | [77] | |
| Liver | Decreased | NASH (MCD) | Mice | [78] |
MCD: Methionine/Choline Deficient Diet; HF: High fructose Diet; HFD: High Fat Diet; HCD: High Cholesterol diet; HF-MCD: High fat- methionine choline deficient diet; nsc: no significant change.
Table 8.
Oxidative stress markers measured in experimental NAFLD/NASH.
| OS Marker | Sample | Level/Activity/Expression in Sample | Experimental Model | Experimental Specie | Reference(s) |
|---|---|---|---|---|---|
| H2O2 | Liver | Increased | NASH (MCD) | C57BL/6J-mtFVB/N mice | [79] |
| Nitrite/nitrate | Liver | Nsc | NAFLD (HFD) | Wistar Rats | [76] |
| TBARS (MDA) | Liver | Increased | NASH | Rat | [80] |
| Liver | Increased | NASH (MCD) | C57BL/6 Mice | [70] | |
| Liver | Increased | Steatosis/NASH (HFD) | Albino rats | [81] | |
| Liver | Increased | NASH (MCD) | C57BL/6 mice | [69] | |
| Liver | Decreased | NAFLD (HCD) | Wistar Rats | [75] | |
| Liver | Increased | Steatosis (HFD/HSD) | Wistar rats | [82] | |
| Liver | Increased | HF/HGD | Wistar rats | [83] | |
| Liver | Increased | NAFLD (HFD) | C57BL/6J mice | [84] | |
| Liver | Increased | NASH (MCD) | C57BL/6 mice | [85] | |
| Liver | Increased | NAFLD/NASH (CDAA diet) | Wistar Rats | [86] | |
| Liver | Increased | NAFLD (HFD) | Sprague-Dawley rats | [74] | |
| Liver | Increased | NASH (MCD) | C57BL6/J mice | [70] | |
| Liver | Increased | NASH | Rat | [80] | |
| Liver | Increased | NAFLD (HFD) | Wistar Rats | [76] | |
| Liver | Increased | NASH (CDHF diet) | Wistar Rats | [87] | |
| Liver | Increased | NASH (MCD) | N-Mary rats | [71] | |
| Liver | Increased | NAFLD (HFD) | Mice | [72] | |
| Liver | Increased | NASH (HF) | Kunming mice | [73] | |
| Liver | Nsc | NASH (MCD) | C57BL/6J-mtFVB/N mice | [79] | |
| Lipid peroxide | Liver | Increased | NASH (MCD) | Wistar Rat | [67] |
| 8-Isoprostanes | Liver | Increased | NASH (HFMCD) | Sprague Dawley Rat | [77] |
| 4-HNE | Liver | Increased | NASH (MCD) | Wistar Rat | [67] |
| Liver | Increased | NASH | Rat | [80] | |
| Liver | Increased | NASH (HFD) | Sprague Dawley Rat | [88] | |
| Liver | Increased | NASH (CDHF diet) | Wistar Rats | [87] | |
| Liver | Increased | NASH | leptin-deficient (ob/ob) mice | [89] | |
| Liver | Increased | NASH (HF-HSD) | C57BL/6 J mice | [90] | |
| Liver | Increased | NASH (MCD) | C57BL6 mice | [54,91,92] | |
| Protein carbonyl | Liver | Increased | NASH (MCD) | N-Mary rats | [93] |
| Nitrotyrosine | Liver | Increased | NAFLD | CYP2E1 transgenic (Tg) mice | [93] |
| Dityrosine | Liver | Increased | NAFLD (HFD) | C57BL/6J mice | [84] |
| Hydroxyproline | Liver | Increased | NAFLD/NASH (CDAA diet) | Wistar Rats | [86] |
| Liver | Increased | NASH (MCD/WD) | C57BL6 mice | [54] | |
| 8-OH-dG | Liver | Increased | NASH (MCD) | Wistar Rat | [69] |
| Liver | Decreased | NASH (HF-MCD) | Sprague-Dawley rats | [79] | |
| Liver | Increased | NAFLD (HFD) | C57BL/6J mice | [86] | |
| CYP2E1 | Liver | Increased | NAFLD | CYP2E1 transgenic (Tg) mice | [92,93] |
| Liver | Increased | NASH (HFD) | Sprague Dawley Rat | [88] | |
| Liver | Increased | NASH (HFD) | Sprague Dawley Rat | [94] | |
| Liver | Increased | Steatosis (HFD/HSD) | Wistar rats | [82] | |
| Liver | Increased | NASH (HFD) | Sprague-Dawley rats | [95] | |
| Liver | Increased | NASH (CDHF diet) | Wistar rats | [96] | |
| NADPH Oxidase | liver | Increased | NASH | ob/ob mice | [89] |
| Xanthine Oxidase | Liver | Increased | NAFLD (HFD) | Sprague-Dawley rats | [74] |
MCD: Methionine/Choline Deficient Diet; HF: High fructose Diet; HFD: High Fat Diet; HCD: High Cholesterol diet; HSD: High Sucrose diet; CDAA: Choline Deficient L-Amino Acid-defined; CDHF: Choline Deficient High Fat diet; HFMCD: High fat methionine choline deficient diet; HF-HSD: High fat-high sucrose diet; WD: Western diet; HF/HGD: High-Fructose/High-Glucose Diet; nsc: no significant change.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ore, A.; Akinloye, O.A. Oxidative Stress and Antioxidant Biomarkers in Clinical and Experimental Models of Non-Alcoholic Fatty Liver Disease. Medicina 2019, 55, 26. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55020026
AMA Style
Ore A, Akinloye OA. Oxidative Stress and Antioxidant Biomarkers in Clinical and Experimental Models of Non-Alcoholic Fatty Liver Disease. Medicina. 2019; 55(2):26. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55020026
Chicago/Turabian StyleOre, Ayokanmi, and Oluseyi Adeboye Akinloye. 2019. "Oxidative Stress and Antioxidant Biomarkers in Clinical and Experimental Models of Non-Alcoholic Fatty Liver Disease" Medicina 55, no. 2: 26. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55020026