The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro

 and
and
Abstract
:1. Introduction
2. Materials and Methods
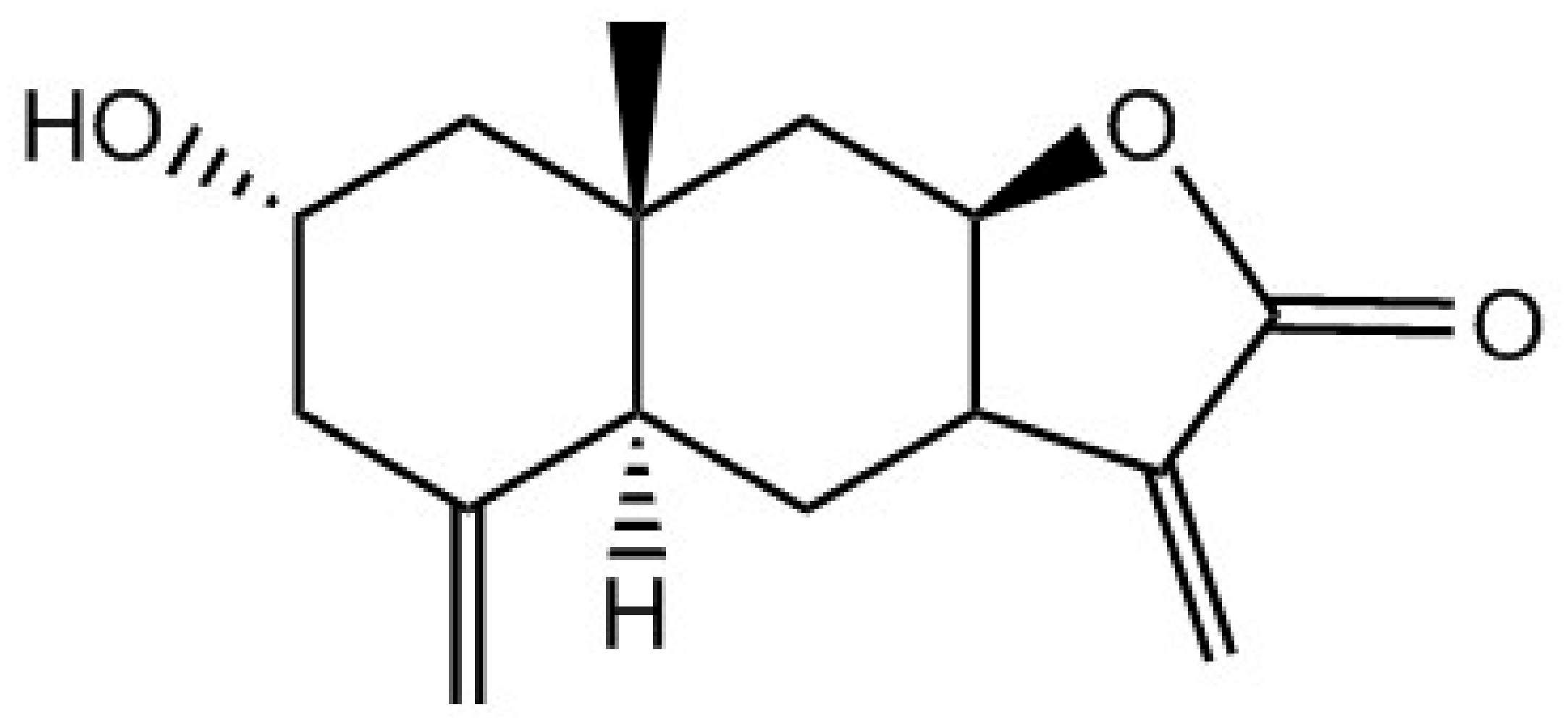
2.1. Chemicals and Reagents
2.2. Cell Lines and Cell Culture
2.3. MTT Assay
2.4. Immunofluorescence Staining
2.5. Western Blot Analysis
2.6. Cellular Tubulin Polymerization Assays
2.7. Flow Cytometry Analysis of Apoptosis
2.8. Cell Cycle Distribution
2.9. Statistical Analysis
3. Results
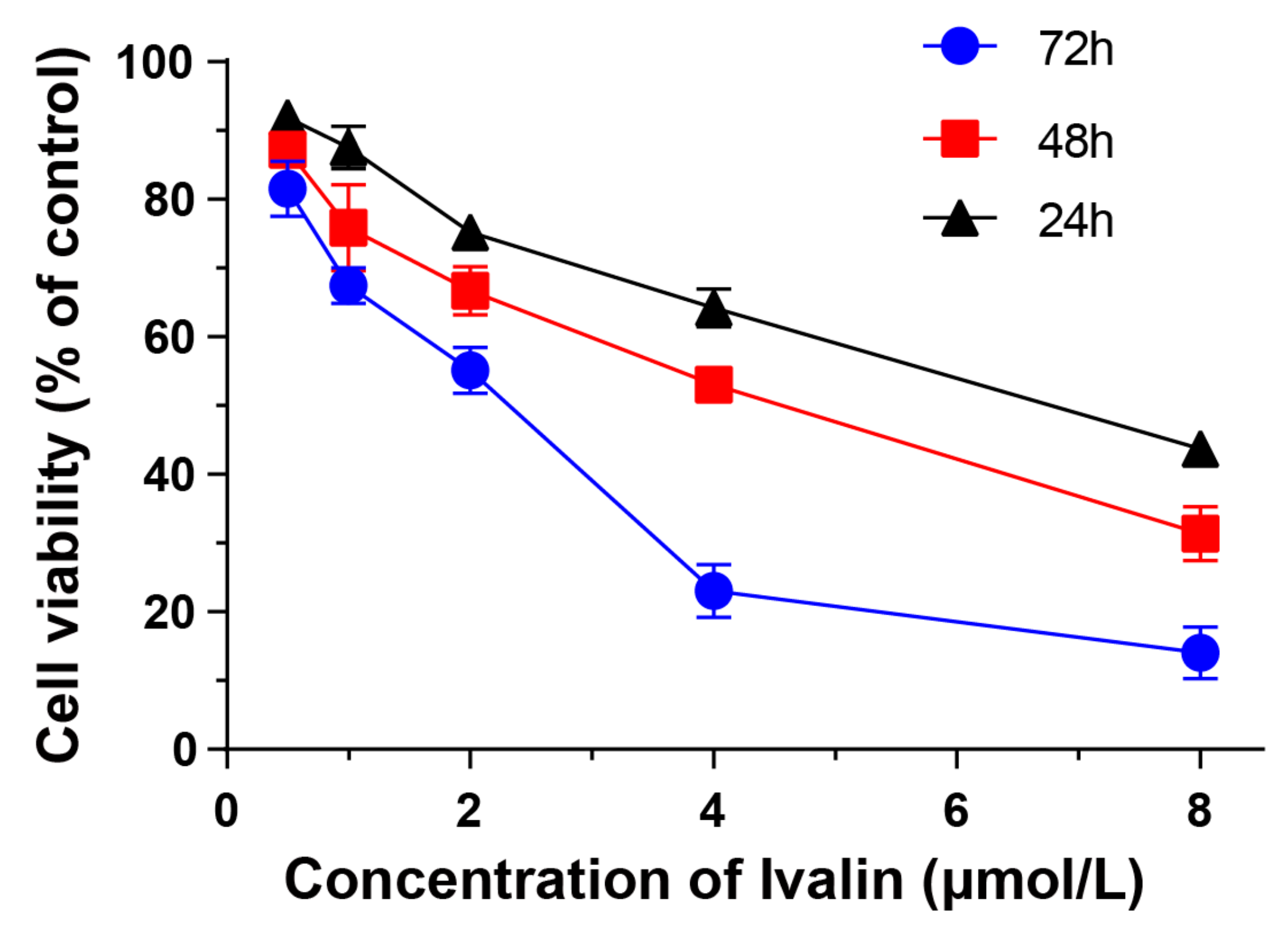
3.1. Cytotoxicity of Ivalin in Human Hepatocellular Carcinoma Cell Lines
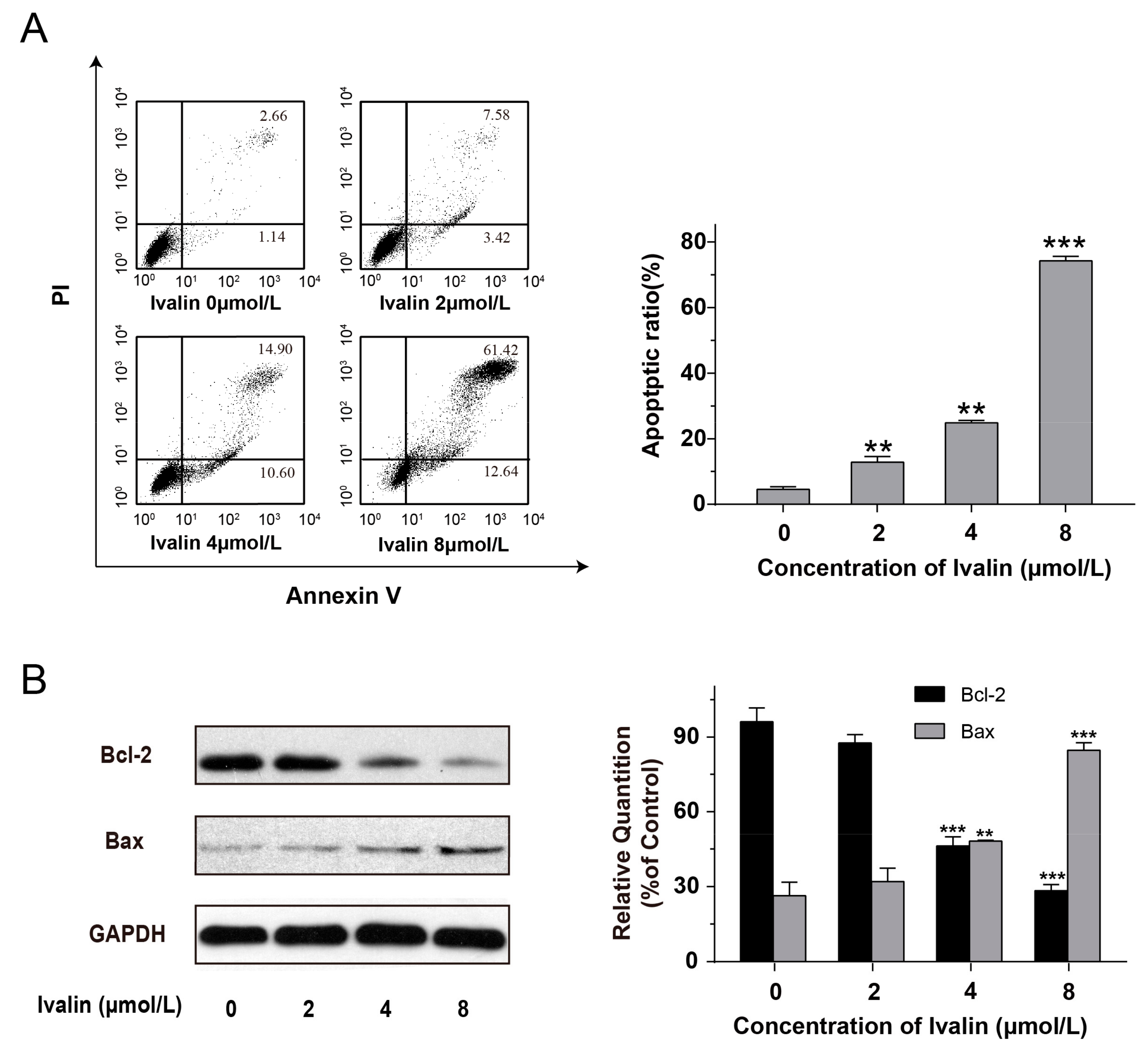
3.2. Effects of Ivalin on Apoptosis in SMMC-7721 Cells
3.3. Effects of Ivalin on Cell Cycle in SMMC-7721 Cells
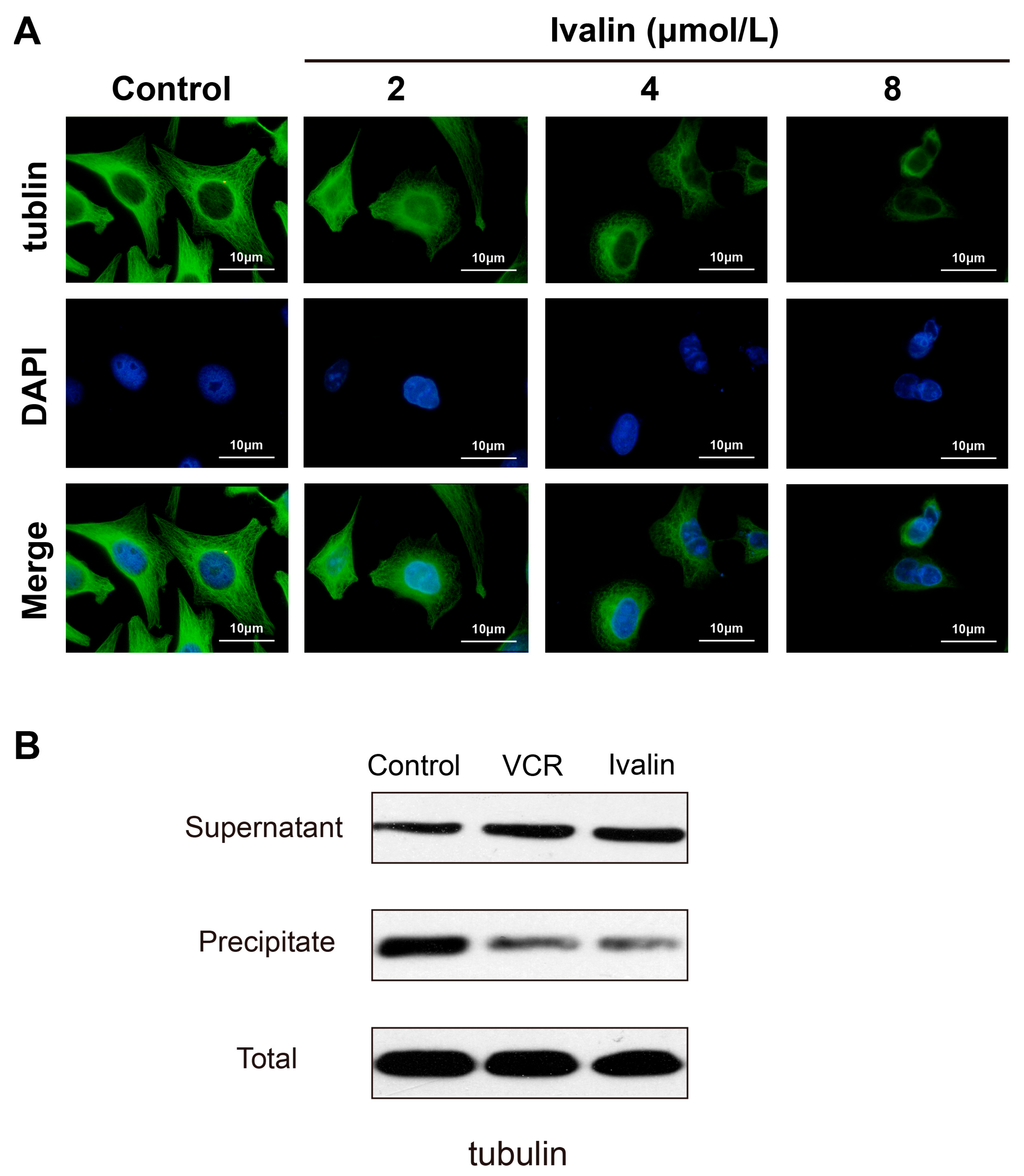
3.4. Effects of Ivalin on the Microtubule Tissue of SMMC-7721 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, E.K.; Vauthey, J. Focus on treatment of large hepatocellular carcinoma. Ann. Surg. Oncol. 2004, 11, 1035–1036. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qin, X.; Zhou, J.; Tu, Z.; Bi, X.; Li, W.; Fan, X.; Zhang, Y. Tissue factor pathway inhibitor-2 inhibits the growth and invasion of hepatocellular carcinoma cells and is inactivated in human hepatocellular carcinoma. Oncol. Lett. 2011, 2, 779–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Li, L.; Ma, H.G.; Sun, P.; Wang, Q.L.; Zhang, T.T.; Shen, Y.M.; Zhu, W.M.; Li, X. Bisindolylmaleimide alkaloid BMA-155Cl induces autophagy and apoptosis in human hepatocarcinoma HepG-2 cells through the NF-kappa B p65 pathway. Acta Pharm. Sin. 2017, 38, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Kavallaris, M. Microtubules and resistance to tubulin-binding agents. Nat. Rev. Cancer 2010, 10, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.J. Microtubule dynamics and kinetochore function in mitosis. Ann. Rev. Cell Biol. 1988, 4, 527–549. [Google Scholar] [CrossRef]
- Rusan, N.M.; Fagerstrom, C.J.; Yvon, A.M.C.; Wadsworth, P. Cell cycle-dependent changes in microtubule dynamics in living cells expressing green fluorescent protein-alpha tubulin. Mol. Biol. Cell 2001, 12, 971–980. [Google Scholar] [CrossRef]
- Jordan, M.A.; Thrower, D.; Wilson, L. Mechanism of inhibition of cell proliferation by vinca alkaloids. Cancer Res. 1991, 51, 2212–2222. [Google Scholar]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, G.; Gill, R.K.; Soni, R.; Bariwal, J. Recent developments in tubulin polymerization inhibitors: An overview. Eur. J. Med. Chem. 2014, 87, 89–124. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Kanakkanthara, A.; Miller, J.H. Microtubule-targeting agents are clinically successful due to both mitotic and interphase impairment of microtubule function. Bioorg. Med. Chem. 2014, 22, 5050–5059. [Google Scholar] [CrossRef] [PubMed]
- Ganapathi, R.N.; Ganapathi, M.K. Mechanisms regulating resistance to inhibitors of topoisomerase II. Front. Pharmacol. 2013, 4, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, E.A. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Mol. Cancer Ther. 2009, 8, 2086–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.H.; Qi, J.; Liu, F.Y.; Lin, S.Q.; Zhang, C.Y.; Xie, W.D.; Zhang, H.Y.; Li, X. Ivalin Inhibits Proliferation, Migration and Invasion by Suppressing Epithelial Mesenchymal Transition in Breast Cancer Cells. Nutr. Cancer Int. J. 2018, 70, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-D.; Wang, X.-R.; Ma, L.-S.; Li, X.; Row, K.-H. Sesquiterpenoids from Carpesium divaricatum and their cytotoxic activity. Fitoterapia 2012, 83, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Sananes, M.R.; Kapuy, O.; Hunt, T.; Novak, B. Switches and latches: A biochemical tug-of-war between the kinases and phosphatases that control mitosis. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3584–3594. [Google Scholar] [CrossRef] [PubMed]
- Luduena, R.F. Multiple forms of tubulin: Different gene products and covalent modifications. In International Review of Cytology—A Survey of Cell Biology; Jeon, K.W., Ed.; Elsevier: Amsterdam, The Netherlands, 1997; Volume 178, pp. 207–275. [Google Scholar]
- Nogales, E. Structural insights into microtubule function. In Annual Review of Biophysics and Biomolecular Structure; Stroud, R.M., Olson, W.K., Sheetz, M.P., Eds.; Annual Reviews: Palo Alto, CA, USA, 2001; pp. 277–302. [Google Scholar]
- Jordan, M.A.; Wilson, L. Microtubules and actin filaments: Dynamic targets for cancer chemotherapy. Curr. Opin. Cell Biol. 1998, 10, 123–130. [Google Scholar] [CrossRef]
- Giannakakou, P.; Sackett, D.; Fojo, T. Tubulin/microtubules: Still a promising target for new chemotherapeutic agents. J. Natl. Cancer Inst. 2000, 92, 182–183. [Google Scholar] [CrossRef]
- Wilson, L.; Jordan, M.A. Microtubule dynamics: Taking aim at a moving target. Chem. Biol. 1995, 2, 569–573. [Google Scholar] [CrossRef]
- Podolski-Renić, A.; Banković, J.; Dinić, J.; Ríos-Luci, C.; Fernandes, M.X.; Ortega, N.; Kovačević-Grujičić, N.; Martín, V.S.; Padrón, J.M. DTA0100, dual topoisomerase II and microtubule inhibitor, evades paclitaxel resistance in P-glycoprotein overexpressing cancer cells. Eur. J. Pharm. Sci. 2017, 105, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.M.; Zhang, X.F.; Huan, X.J.; Song, S.S.; Wang, W.; Tian, Q.T.; Sun, Y.M.; Chen, Y.; Ding, J.; Wang, Y.Q.; et al. Dual targeting of microtubule and topoisomerase II by alpha-carboline derivative YCH337 for tumor proliferation and growth inhibition. Oncotarget 2015, 6, 8960–8973. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, S.B. How to make taxol from scratch. Nature 1994, 367, 593–594. [Google Scholar] [CrossRef] [PubMed]
- Hastie, S.B. Interactions of colchicine with tubulin. Pharmacol. Ther. 1991, 51, 377–401. [Google Scholar] [CrossRef]
- Na, G.C.; Timasheff, S.N. Thermodynamic linkage between tubulin self-association and the binding of vinblastine. Biochemistry 1980, 19, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Wendell, K.; Gardiner, S.; Derry, W.B.; Copp, H.; Wilson, L. Mitotic block induced in HeLa cells by low concentrations of paclitaxel (Taxol) results in abnormal mitotic exit and apoptotic cell death. Cancer Res. 1996, 56, 816–825. [Google Scholar] [PubMed]
- Duesbery, N.S.; Vande Woude, G.F. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. Biol. Cell. 1998, 90, 461–466. [Google Scholar]
- Doree, M.; Labbe, J.C.; Picard, A. M phase-promoting factor: Its identification as the M phase-specific H1 histone kinase and its activation by dephosphorylation. J. Cell Sci. 1989, 1989, 39–51. [Google Scholar] [CrossRef]
- Cheng, E. Molecular Control of Mitochondrial Apoptosis by the BCL-2 Family. Blood 2009, 114, 1577–1578. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | IC50 (μmol/L) |
|---|---|
| SMMC-7721 | 4.34 ± 0.10 |
| HepG-2 | 5.45 ± 0.13 |
| Plc-prf-5 | 11.33 ± 1.00 |
| Hu-7 | 13.01 ± 0.42 |
| HL7702 | 25.86 ± 0.87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Lin, S.; Zhang, C.; Ma, J.; Han, Z.; Jia, F.; Xie, W.; Li, X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina 2019, 55, 470. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080470
Liu F, Lin S, Zhang C, Ma J, Han Z, Jia F, Xie W, Li X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina. 2019; 55(8):470. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080470
Chicago/Turabian StyleLiu, Fangyuan, Shiqi Lin, Caiyun Zhang, Jiahui Ma, Zhuo Han, Fujuan Jia, Weidong Xie, and Xia Li. 2019. "The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro" Medicina 55, no. 8: 470. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080470