An Experimental Study to Evaluate the Protective Effects of Solanum lycopersicum Seed Essential Oil on Diabetes-Induced Testicular Injuries

Abstract
:1. Introduction
2. Materials and Methods
2.1. Essential Oil Extraction
2.2. Antioxidant Activity Measurement with 2,2-Diphenyl-1-Picrylhydrazyl (DPPH)
2.3. Animals and Experimental Design
2.4. Hormone Assay
2.5. Antioxidant Enzymes Activities
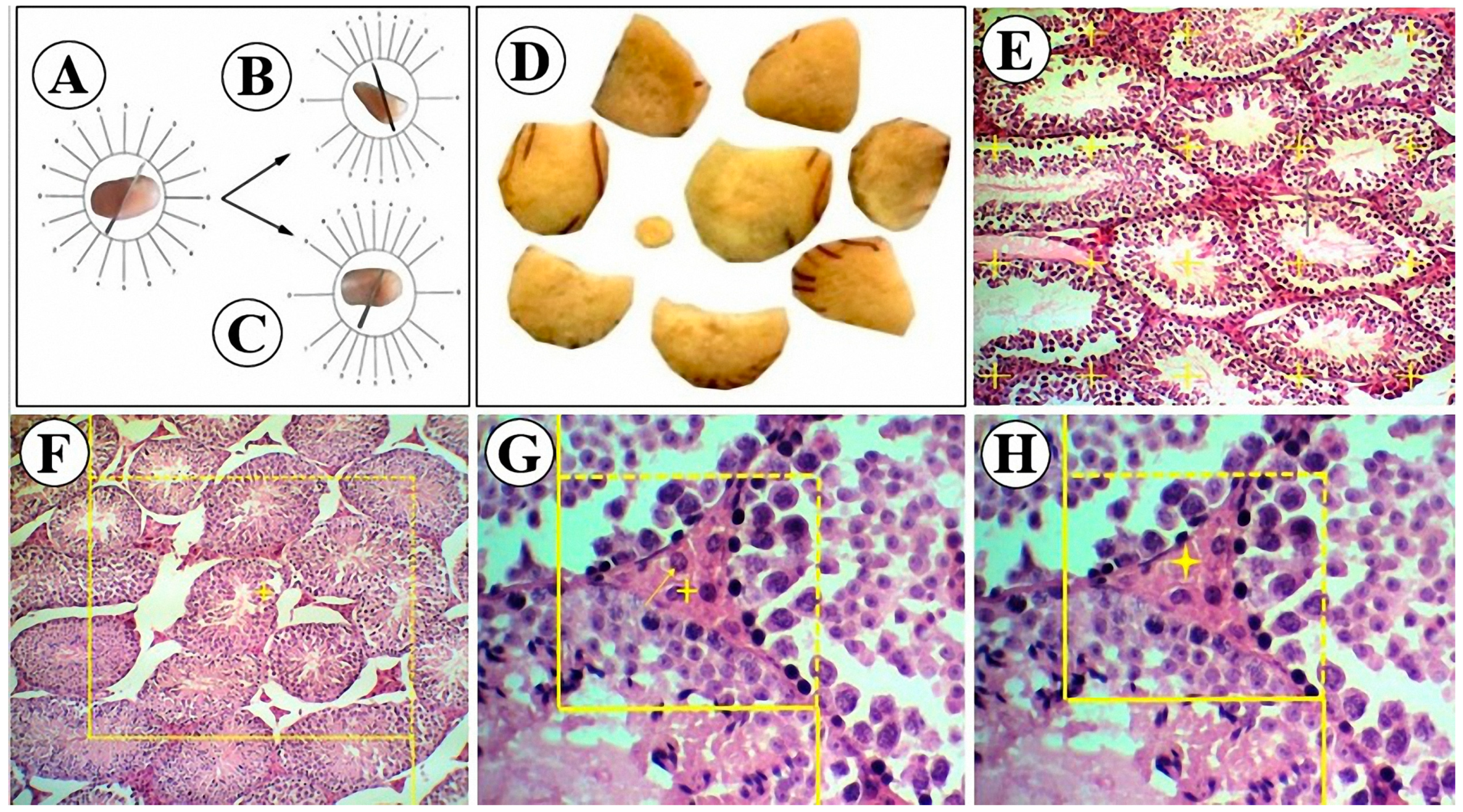
2.6. Stereological Measurements
2.6.1. Volume Estimation
2.6.2. Tubule Length Estimation
2.6.3. Germinal Epithelium Height Estimation
2.6.4. Number Estimation
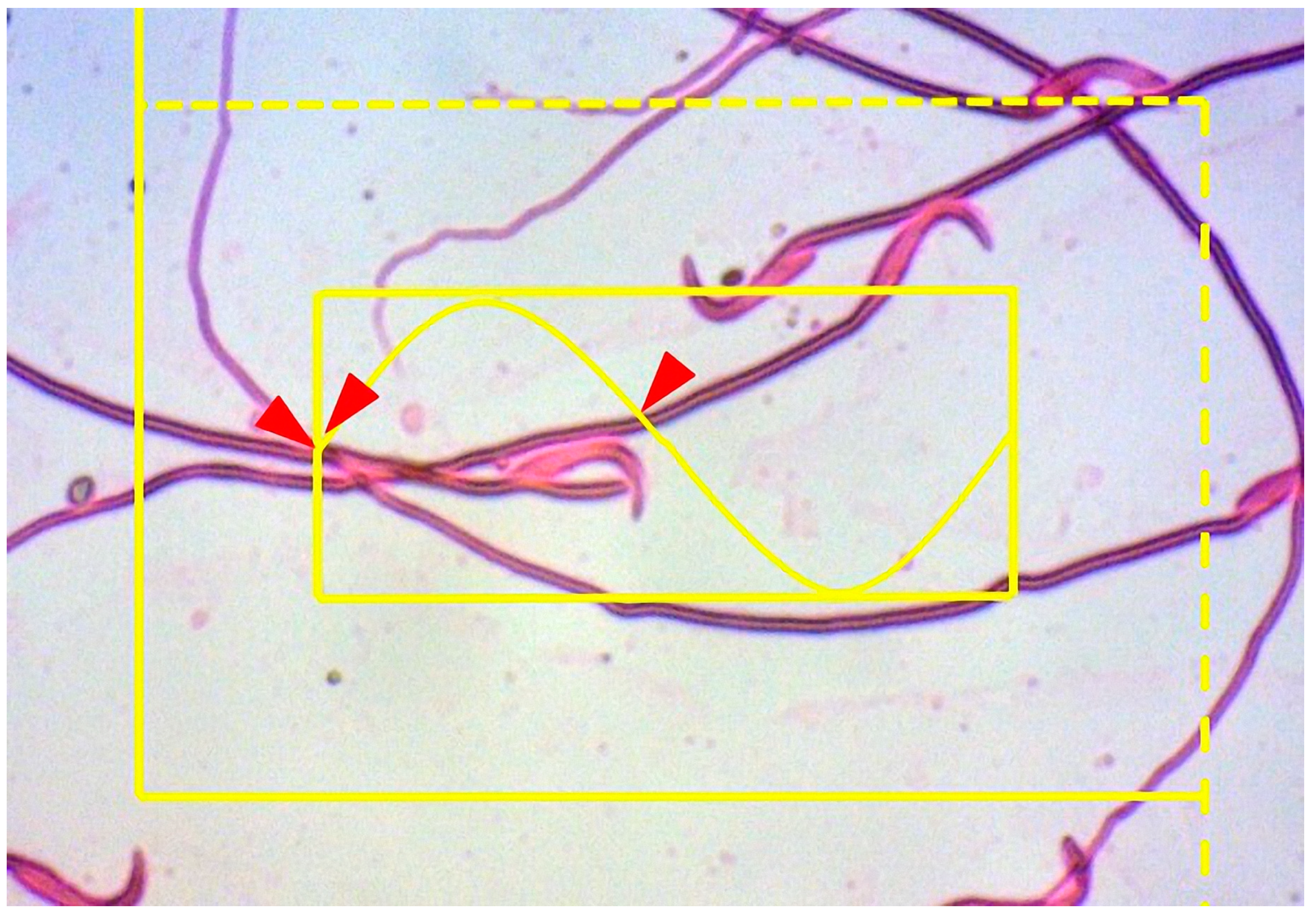
2.6.5. Estimation of the Sperm Tail Length
2.7. Immunohistochemical Assay
2.8. Statistical Analysis
3. Results
3.1. GC/MS Analysis of SL Seed Essential Oil
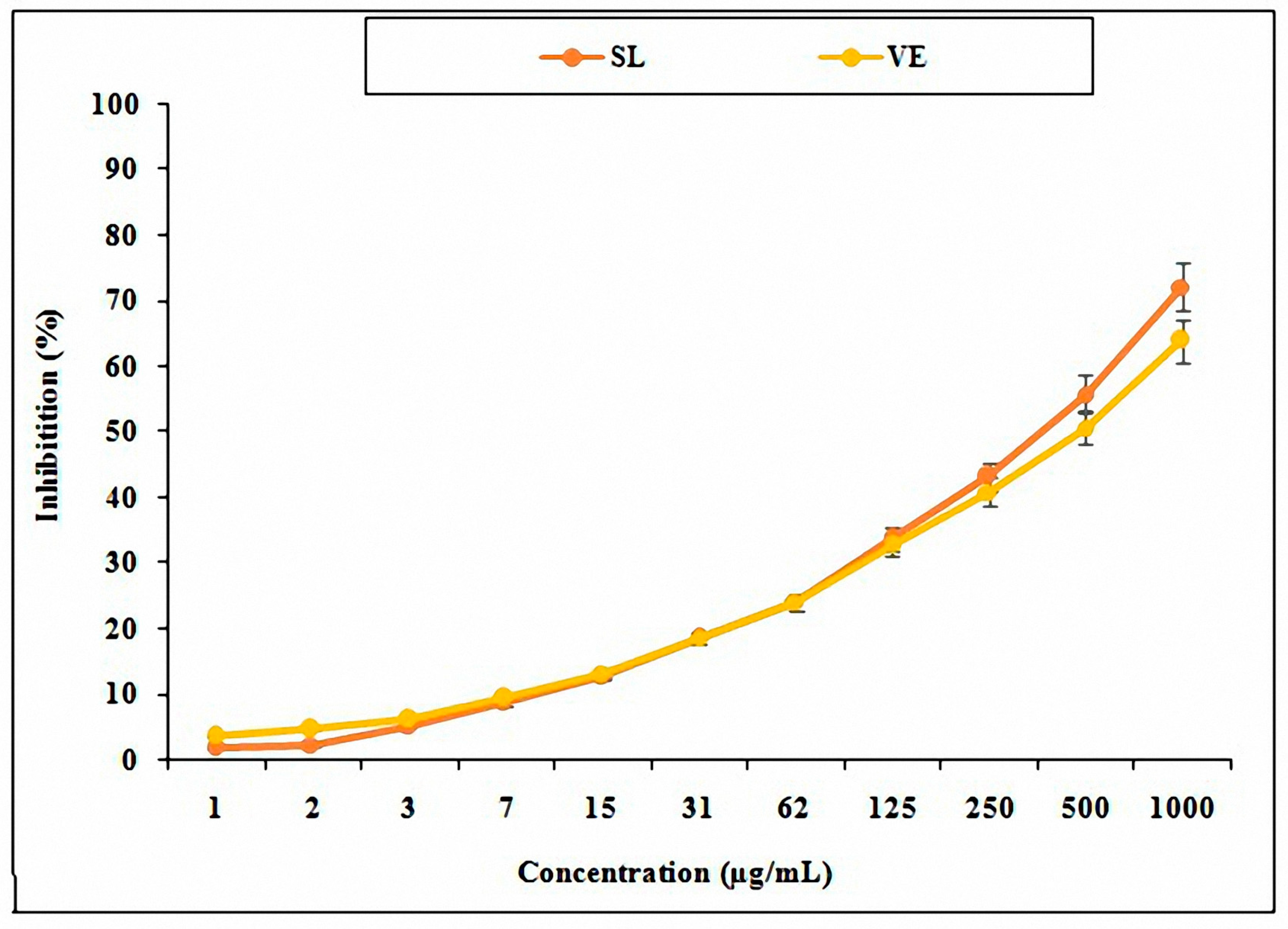
3.2. Antioxidant Activity of SL Seed Essential Oil
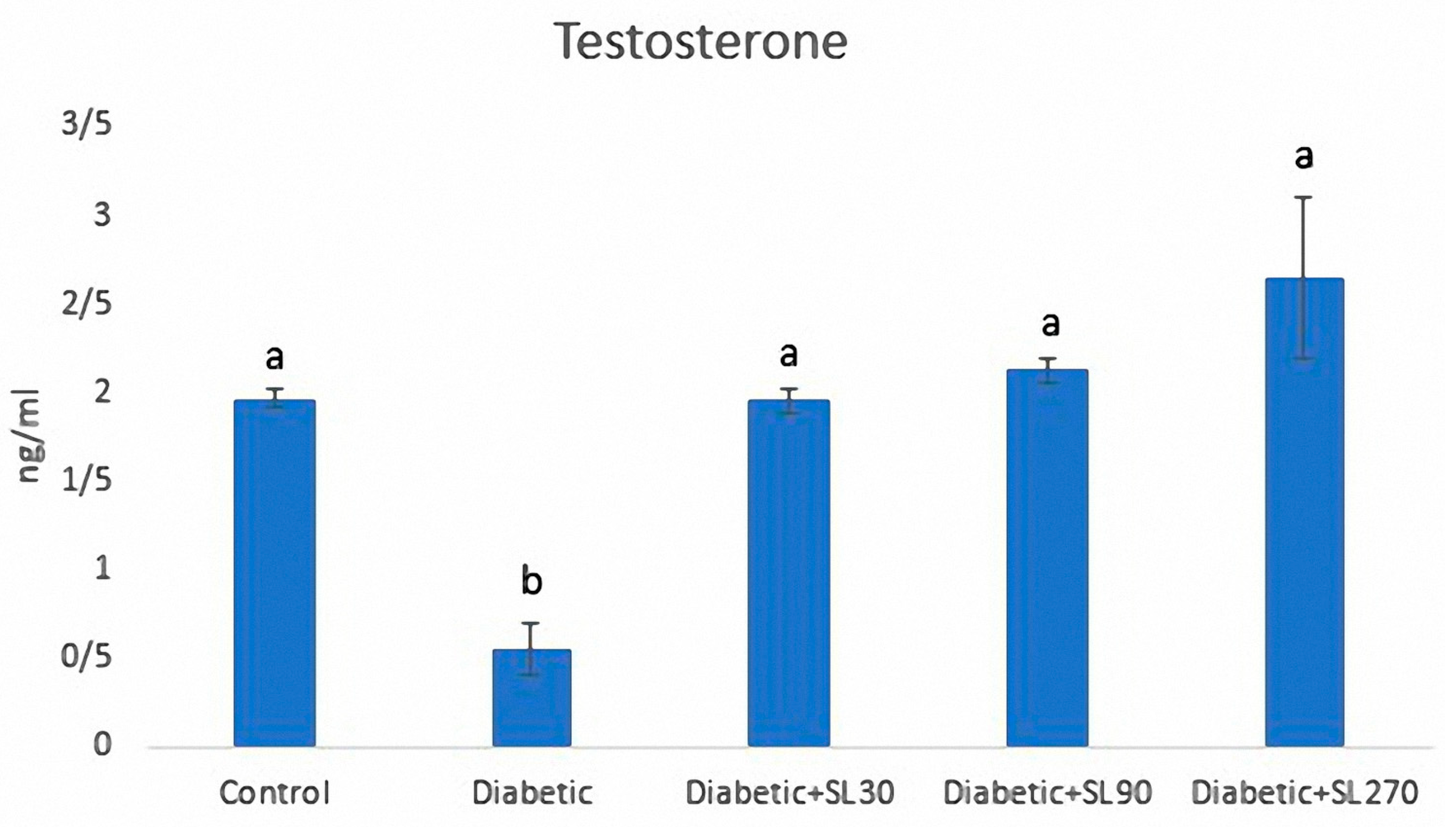
3.3. Serum Testosterone Levels
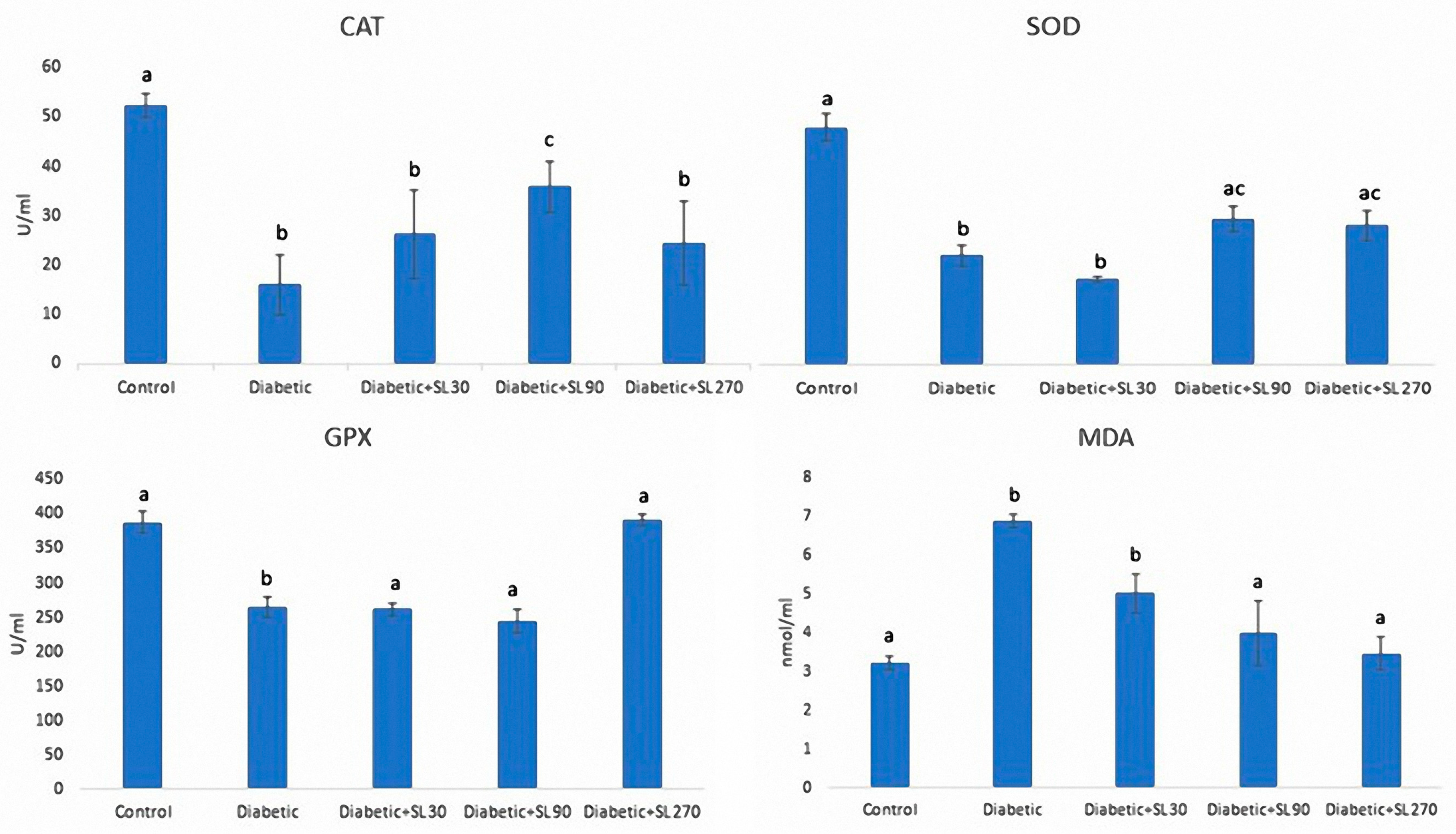
3.4. Antioxidant Enzymes and MDA Activities
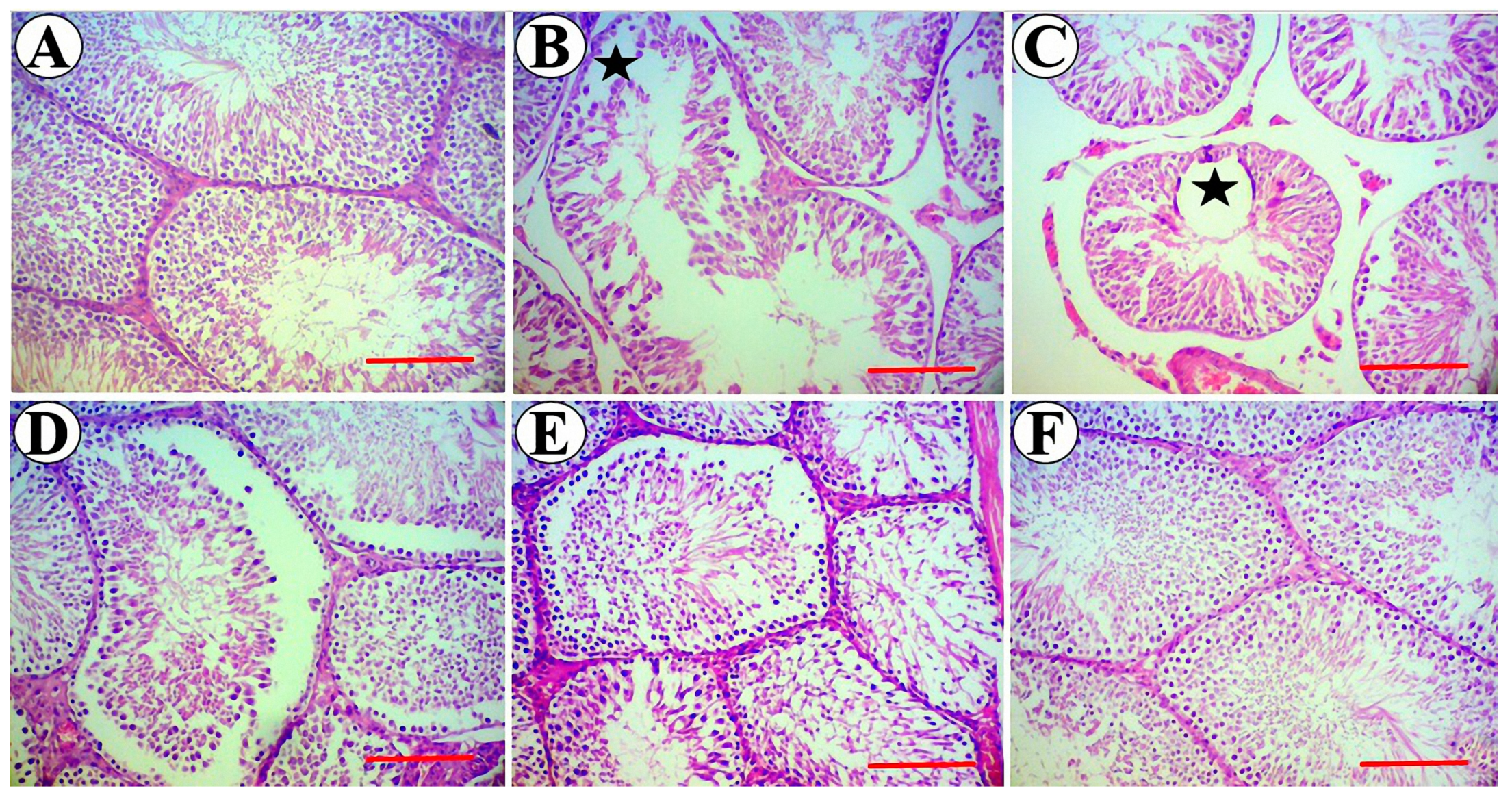
3.5. Testis Stereological Parameters
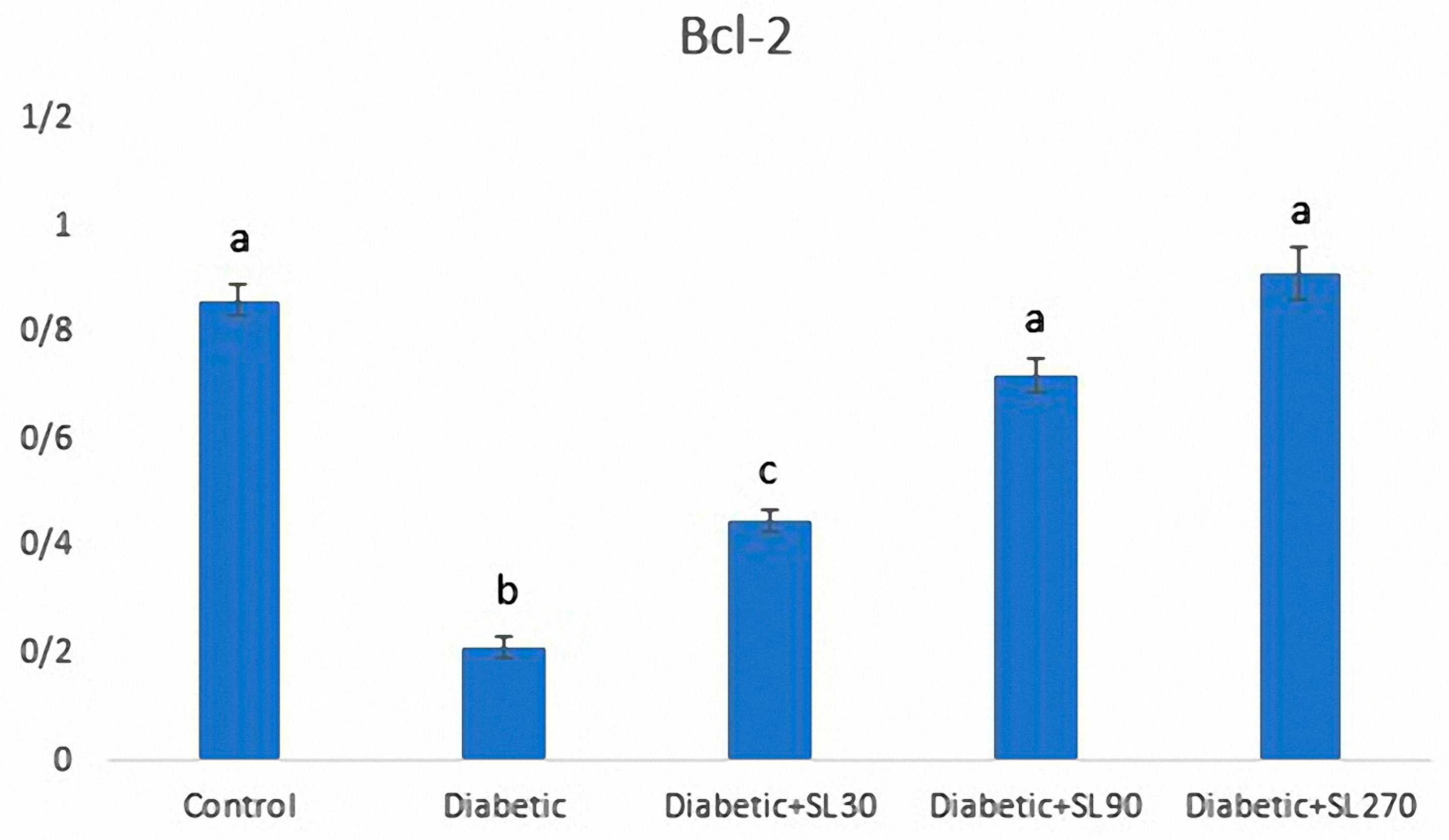
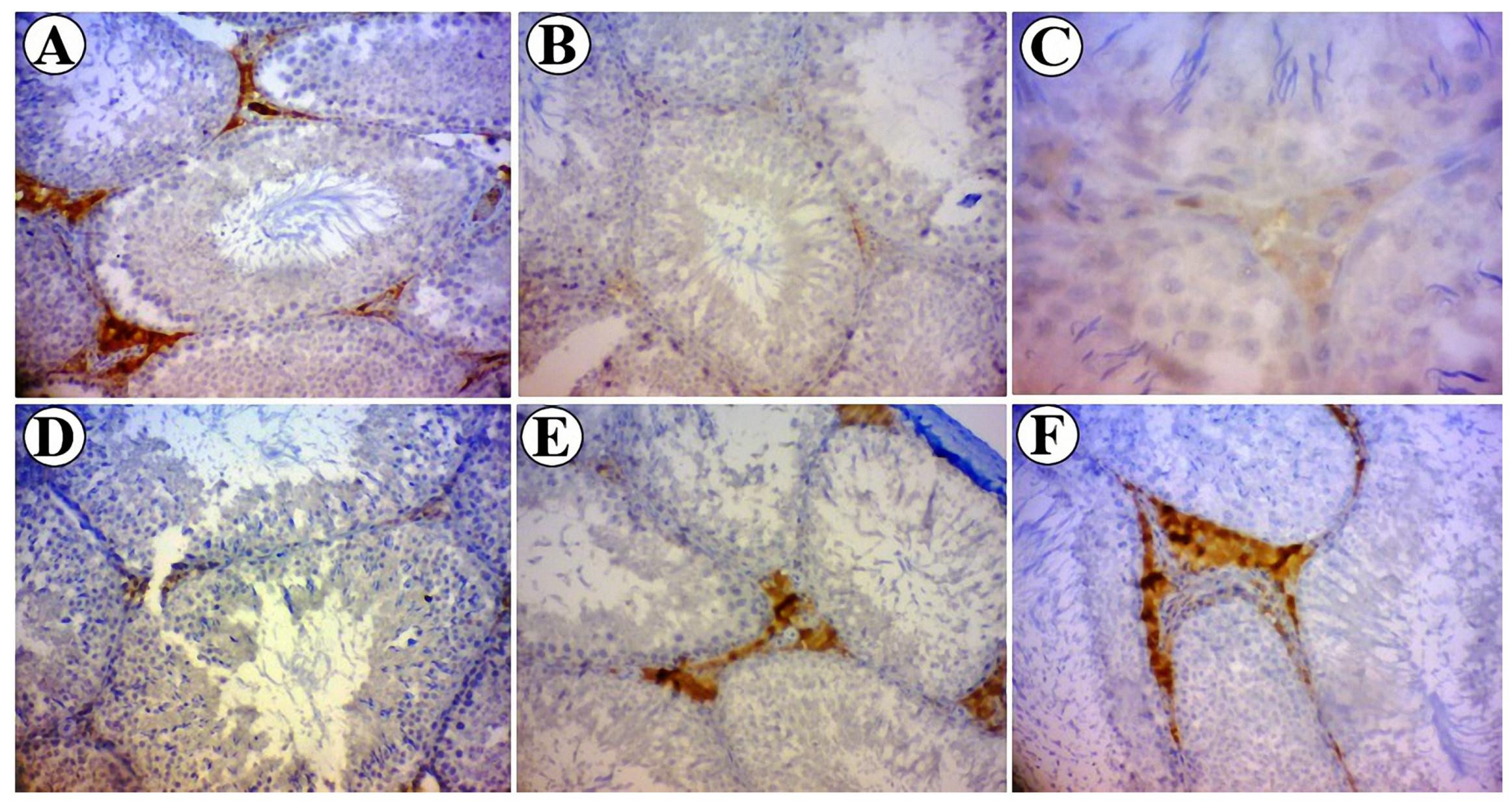
3.6. Expression of Bcl-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maiti, R.D.; Das, J.U.; Ghosh, D. Antidiabetic effect of aqueous extracts of seeds of Tamarindusindica tamarind seed in streptozotocin induce diabetic rat. J. Ethnopharmacol. 2004, 92, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Vignera, S.; Condorelli, R.; Vicari, E.; D’Agata, R.; Calogero, A.E. Diabetes mellitus and sperm parameters. J. Androlol. 2012, 33, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Agbaje, I.; Rogers, D.; McVicar, C.; McClure, N.; Atkinson, A.; Mallidis, C.; Lewis, S.E.M. Insulin dependant diabetes mellitus: Implications for male reproductive function. Hum. Reprod. 2007, 22, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.B.A.; Petersen, C.G.; Mauri, A.L.; Vagnini, L.D.; Renzi, A.; Petersen, B.; Franco, J.G., Jr. Association between bodymass index and sperm quality and sperm DNA integrity. A large population study. Andrologia 2017, 50, e12889. [Google Scholar] [CrossRef] [PubMed]
- Scarano, W.R.; Messias, A.G.; Oliva, S.U.; Klinefelter, G.R.; Kempinas, W.G. Sexual behavior, sperm quantity and quality after short-term streptozotocin-induced hyperglycaemia in rats. Int. J. Androl. 2006, 29, 482–488. [Google Scholar] [CrossRef] [PubMed]
- De Young, L.; Yu, D.; Bateman, R.M.; Brock, G.B. Oxidative stress and antioxidant therapy: Their impact in diabetesassociatederectile dysfunction. J. Androl. 2004, 25, 830–836. [Google Scholar] [CrossRef]
- Sainio-Pollanen, S.; Henriksen, K.; Parvinen, M.; Simell, O.; Pollanen, P. Stage-specific degeneration of germ cells in the seminiferous tubules of non-obese diabetic mice. Int. J. Androl. 1997, 20, 243–253. [Google Scholar] [CrossRef]
- Cai, L.; Chen, S.; Evans, T.; Deng, D.X.; Mukherjee, K.; Chakrabarti, S. Apoptotic germ-cell death and testicular damage in experimental diabetes: Prevention by endothelin antagonism. Urol. Res. 2000, 28, 342–347. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, S.; Wang, Z.; Li, Y.; Guo, W.; Lin, C.; Cai, L. Repetitive exposures to low-dose X-rays attenuate testicular apoptotic cell death in streptozotocin-induce diabetes rats. Toxicol. Lett. 2010, 192, 356–364. [Google Scholar] [CrossRef]
- Koh, P.O. Streptozotocin-induced diabetes increases apoptosis through JNK phosphorylation and Bax activation in rat testes. J. Vet. Med. Sci. 2007, 69, 969–971. [Google Scholar] [CrossRef]
- Artimani, T.; Amiri, I.; Soleimani Asl, S.; Saidijam, M.; Hasanvand, D.; Afshar, S. Amelioration of diabetes-induced testicular and sperm damage in rats by cerium oxide nanoparticle treatment. Andrologia 2018, 50, e13089. [Google Scholar] [CrossRef]
- Al-Roujeaie, A.S.; Abuohashish, H.M.; Ahmed, M.M.; Alkhamees, O.A. Effect of rutin on diabetic-induced erectile dysfunction: Possible involvement of testicular biomarkers in male rats. Andrologia 2017, 49, e12737. [Google Scholar] [CrossRef]
- Guneli, E.; Tugyan, K.; Ozturk, H.; Gumustekin, M.; Cilaker, S.; Uysal, N. Effect of Melatonin on Testicular Damage in Streptozotocin-Induced Diabetes Rats. Eur. Surg. Res. 2008, 40, 354–360. [Google Scholar] [CrossRef]
- Sebai, H.; Selmi, S.; Rtibi, K.; Gharbi, N.; Sakly, M. Protective Effect of Lavandula stoechas and Rosmarinus officinalis essential oils against reproductive damage and oxidative stress in alloxan-induced diabetic rats. J. Med. Food 2015, 18, 241–249. [Google Scholar] [CrossRef]
- Sakhaee, E.; Emadi, L.; Zari, O.; Kheirandish, R.; EsmailiNejad, M.R.; ShafieiBafti, H. Effects of Cuminumcyminum L. essential oil on some epididymal sperm parameters and histopathology of testes following experimentally induced copper poisoning in mice. Andrologia 2016, 48, 542–547. [Google Scholar] [CrossRef]
- Nasimi, P.; Vahdati, A.; Tabandeh, M.R.; Khatamsaz, S. Cytoprotective and anti-apoptotic effects of Saturejakhuzestanica essential oil against busulfan-mediated sperm damage and seminiferous tubules destruction in adult male mice. Andrologia 2016, 48, 74–81. [Google Scholar] [CrossRef]
- Sariözkan, S.; Türk, G.; Güvenç, M.; Yüce, A.; Özdamar, S.; Cantürk, F.; Yay, A.H. Effects of Cinnamon (C. zeylanicum) Bark Oil Against Taxanes-Induced Damages in Sperm Quality, Testicular and Epididymal Oxidant/Antioxidant Balance, Testicular Apoptosis, and Sperm DNA Integrity. Nutr. Cancer 2016, 68, 481–494. [Google Scholar] [CrossRef]
- Liu, Q.; Duan, R.J.; Zhou, Y.F.; Wei, H.K.; Peng, J.; Li, J.L. Supplementing oregano essential oil to boar diet with strengthened fish oil: Effects on semen antioxidant status and semen quality parameters. Andrologia 2017, 49, e12764. [Google Scholar] [CrossRef]
- Elmi, A.; Ventrella, D.; Barone, F.; Carnevali, G.; Filippini, G.; Pisi, A.; Benvenuti, S.; Scozzoli, M.; Bacci, M.L. In Vitro Effects of Tea Tree Oil (Melaleuca Alternifolia Essential Oil) and its Principal Component Terpinen-4-ol on Swine Spermatozoa. Molecules 2019, 24, 1071. [Google Scholar] [CrossRef]
- Willcox, J.K.; Catignani, G.L.; Lazarus, S. Tomatoes and cardiovascular health. Crit. Rev. Food Sci. Nutr. 2003, 43, 1–18. [Google Scholar] [CrossRef]
- Bose, K.S.; Agrawal, B.K. Effect of lycopene from cooked tomatoes on serum antioxidant enzymes, lipid peroxidation rate and lipid profile in coronary heart disease. Singap. Med. J. 2007, 48, 415–420. [Google Scholar]
- Rafi, M.M.; Yadav, P.N.; Reyes, M. Lycopene inhibits LPS-induced pro-inflammatorymediator inducible nitric oxide synthase in mouse macrophage cells. J. Food Sci. 2007, 72, S069–S074. [Google Scholar] [CrossRef]
- Dou, W.; Zhang, J.; Sun, A.; Zhang, E.; Ding, L.; Mukherjee, S.; Wei, X.; Chou, G.; Wang, Z.T.; Mani, S. Protective effect of naringenin against experimental colitis via suppression of Toll-like receptor 4/NF-kappa B signaling. Br. J. Nutr. 2013, 110, 599–608. [Google Scholar] [CrossRef]
- Thomson, M.; Ali, M. Garlic [Aliium sativum]: A review of its potential use as an anti-cancer agent. Curr. Cancer Drug Targets 2003, 3, 67–81. [Google Scholar] [CrossRef]
- Etminan, M.; Takkouche, B.; Caamaño-Isorna, F. The role of tomato products andlycopene in the prevention of prostate cancer: A meta-analysis of observational studies. Cancer Epidemiol. Prev. Biomark. 2004, 13, 340–345. [Google Scholar]
- Canene-Adams, K.; Lindshield, B.L.; Wang, S.; Jeffery, E.H.; Clinton, S.K.; Erdman, J.W., Jr. Combinations of tomato and broccoli enhance antitumor activity in dunning r3327-h prostate adenocarcinomas. Cancer Res. 2007, 2, 836–843. [Google Scholar] [CrossRef]
- Rissanen, T.; Voutilainen, S.; Nyyssonen, K.; Salonen, J.T. Lycopene, atherosclerosis, and coronary heart disease. Exp. Biol. Med. 2002, 227, 900–907. [Google Scholar] [CrossRef]
- Frederiksen, H.; Rasmussen, S.E.; Schroder, M.; Bysted, A.; Jakobsen, J.; Frandsen, H.; Ravn-Haren, G.; Mortensen, A. Dietary supplementation with an extract of lycopene rich tomatoes does not reduce atherosclerosis in Watanabe Heritable Hyperlipidemic rabbits. Br. J. Nutr. 2007, 97, 6–10. [Google Scholar] [CrossRef]
- Neuman, I.; Nahum, H.; Ben-Amotz, A. Reduction of exercise-induced asthma oxidative stress by lycopene, a natural antioxidant. Allergy 2000, 55, 1184–1189. [Google Scholar] [CrossRef]
- Wood, L.G.; Garg, M.L.; Blake, R.J.; Gibson, P.G. Carotenoid concentrations in asthmatics versus healthy controls. Asia Pac. J. Clin. Nutr. 2004, 13, S74. [Google Scholar]
- Barbosa-Filho, J.M.; Alencar, A.A.; Nunes, X.P.; Tomaz, A.C.A.; Sena-Filho, J.G.; Athayde-Filho, P.F.; Silva, M.S.; Souza, M.F.; Da-Cunha, E.V.L. Sources of alpha-, beta-, gamma-, delta-and epsilon-carotenes: A twentieth century review. Rev. Bras. Farmacogn. 2008, 18, 135–154. [Google Scholar] [CrossRef]
- Gonçalves, M.C.R.; Diniz, M.F.F.M.; Borba, J.D.C.; Nunes, X.P.; Barbosa-Filho, J.M. Berinjela (Solanum melongena L.)—Mitoourealidade no combate as dislipidemias? Rev. Bras. Farmacogn. 2006, 16, 252–257. [Google Scholar] [CrossRef]
- Hosseinimehr, S.J.; Mahmoudzadeh, A.; Ahmadi, A.; Ashrafi, S.A.; Shafaghati, N.; Hedayati, N. The radioprotective effect of Zataria multiflora against genotoxicity induced by γ irradiation in human blood lymphocytes. Cancer Biother. Radiopharm. 2011, 26, 325–329. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Martin, J.P.; Dailey, M.; Sugarman, E. Negative and positive assays of superoxidedismutase based on hematoxylin autoxidation. Arch. Biochem. Biophys. 1987, 255, 329–336. [Google Scholar] [CrossRef]
- Ho, Y.S.; Magnenat, J.L.; Bronsoni, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1966, 16, 359–364. [Google Scholar] [CrossRef]
- Gundersen, H.J.; Bendtsen, T.F.; Korbo, L.; Marcussen, N.; Møller, A.; Nielsen, K.; Nyengaard, J.R.; Pakkenberg, B.; Sørensen, F.B.; Vesterby, A.; et al. Some new, simple and efficient stereological methods and their use in pathological research and diagnosis. APMIS 1988, 96, 379–394. [Google Scholar] [CrossRef]
- Nyengaard, J.R. Stereologic methods and their application in kidney research. J. Am. Soc. Nephrol. 1999, 10, 1100–1123. [Google Scholar]
- Howard, C.V.; Reed, M. Unbiased Stereology: Three-Dimensional Measurementin Microscopy, 2nd ed.; Bios Scientific Publisher: Oxford, UK, 2005. [Google Scholar]
- Sterio, D.C. The unbiased estimation of number and sizes of arbitrary particles using the disector. J. Microsc. 1984, 134, 127–136. [Google Scholar] [CrossRef]
- Bjorndahl, L.; Soderlund, I.; Kvist, U. Evaluation of the OneStep Eosin-Nigrosin Staining Technique for Human Sperm Vitality Assessment. Hum. Reprod. 2003, 18, 813–816. [Google Scholar] [CrossRef]
- Khorsandi, L.; Mirhoseini, M.; Mohamadpour, M.; Orazizadeh, M.; Khaghani, S. Effect of curcumin on dexamethasone-induced testicular toxicity in mice. Pharm. Biol. 2013, 51, 206–212. [Google Scholar] [CrossRef]
- Navarro-Casado, L.; Juncos-Tobarra, M.A.; Chafer-Rudilla, M.; de Onzono, L.I.; Blazquez-Cabrera, J.A.; Miralles-Garcia, J.M. Effect of experimental diabetes and STZ on male fertility capacity. Study in rats. J. Androl. 2010, 31, 584–592. [Google Scholar] [CrossRef]
- Bahey, N.G.; Soliman, G.M.; El-Deeb, T.A.; El-Drieny, E.A. Influence of insulin and testosterone on diabetic rat ventral prostate: Histological, morphometric and immunohistochemical study. J. Microsc. Ultrasuct. 2014, 2, 151–160. [Google Scholar]
- Steger, R.W.; Rabe, M.B. The effect of diabetes mellitus on endocrine and reproductive function. Proc. Soc. Exp. Biol. Med. 1997, 214, 1–11. [Google Scholar] [CrossRef]
- Numila, R.; Gross, R.; Rchid, H.; Manteghetti, M.; Petit, P.; Tijane, M.; Ribes, G.; Sauvaire, Y. Insulin tropic effect of citrulluscolocynthis fruit extract. Planta Med. 2000, 66, 418–423. [Google Scholar] [CrossRef]
- Sharifi, A.M.; Mousavi, S.H.; Farhadi, M.; Larijani, B. Study of high glucose-induced apoptosis in PC12 cells: Role of bax protein. J. Pharmacol. Sci. 2007, 104, 258–262. [Google Scholar] [CrossRef]
- Donmez, Y.B.; Kizilay, G.; Topcu-Tarladacalisir, Y. MAPK immunoreactivity in streptozotocin-induced diabetic rat testis. Acta Cir. Bras. 2014, 29, 644–650. [Google Scholar] [CrossRef]
- Roy, S.; Metya, S.K.; Rahaman, N.; Sannigrahi, S.; Ahmed, F. Ferulic acid in the treatment of post-diabetes testicular damage: Relevance to the down regulation of apoptosis correlateswith antioxidant status via modulation of TGF-β1, IL-1β and Aktsignalling. Cell Biochem. Funct. 2014, 32, 115–124. [Google Scholar] [CrossRef]
- Khaki, A.; Fathiazad, F.; Nouri, M.; Khaki, A.; Maleki, N.A.; Khamnei, H.J.; Ahmadi, P. Beneficial effects of quercetin on sperm parameters in streptozotocin-induced diabetic male rats. Phytother. Res. 2010, 24, 1285–1291. [Google Scholar] [CrossRef]
- Navarrete, S.; Alarcon, M.; Palomo, I. Aqueous Extract of Tomato (Solanum lycopersicum L.) andFerulic Acid Reduce the Expression of TNF-α and IL-1β in LPS-Activated Macrophage. Molecules 2015, 20, 15319–15329. [Google Scholar] [CrossRef]
- Wang, B.; Trayhurn, P. Acute and prolonged effects of TNF-alpha on the expression andsecretion of inflammation-related adipokines by human adipocytes differentiated in culture. Pflug. Arch. 2006, 452, 418–427. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Peraldi, P.; Budavari, A.; Ellis, R.; White, M.F.; Spiegelman, B.M. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance. Science 1996, 271, 665–668. [Google Scholar] [CrossRef]
- Dinarello, C.A.; van der Meer, J.W. Treating inflammation by blocking interleukin-1 in humans. Semin. Immunol. 2013, 25, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Chuang, L.T.; Leonard, A.E.; Liu, J.W.; Mukerji, P.; Bray, T.M.; Huang, Y.S. Inhibitory effect of conjugated linoleic acid on linoleic acid elongation in transformed yeast with human elongase. Lipids 2001, 36, 1099–1103. [Google Scholar] [CrossRef]
- Watson, A.D. Thematic review series: Systems biology approaches to metabolic and cardiovascular disorders. Lipidomics: A global approach to lipid analysis in biological systems. JLR J. Lipid Res. 2006, 47, 2101–2111. [Google Scholar] [CrossRef]
- Van den Berg, J.J.M.; Cook, N.E.; Tribble, D.L. Reinvestigation of the antioxidant properties of conjugated linoleic acid. Lipids 1995, 30, 599–605. [Google Scholar] [CrossRef]
- Yurawecz, M.P.; Hood, J.K.; Mossoba, M.M.; Roach, J.A.G.; Ku, Y. Furan fatty acids determined as oxidation products of conjugated octadecadienoic acid. Lipids 1995, 30, 595–598. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Chan, P.T.; Kwan, K.Y.; Zhang, A. Reassessment of the antioxidant activity of conjugaed linoleic acids. J. Am. Oil Chem. Soc. 1997, 74, 749–753. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | RT (min) | Percentage of the Total Chromatogram Area |
|---|---|---|
| 2H-Oxireno [3,4] cyclopenta [1,2-c] furan-2-one, 1a,1b,4,4a,5,5a-hexahydro-4-(dimethoxymethyl)-, (1bR,1a-cis,4-trans,4a-cis,5a-cis)- | 6.057 | 0.305% |
| Nonanal dimethyl acetal | 6.123 | 0.264% |
| 9-Hexadecanoic acid, methyl ester, (Z)- | 10.999 | 0.301% |
| adecanoic acid, methyl ester | 11.179 | 18.509% |
| n-Hexadecanoic acid | 11.476 | 0.722% |
| 9,12-Octadecadienoic acid (Z,Z)-, methyl ester | 12.696 | 49.68% |
| 9-Octadecenoic acid (Z)-, methyl ester | 12.735 | 22.021% |
| Octadecanoic acid, methyl ester | 12.898 | 6.474% |
| 9,12-Octadecadienoic acid (Z,Z)- | 12.983 | 0.98% |
| Octadecanoic acid | 13.195 | 0.37% |
| Hexadecanoic acid, 15-methyl-, methyl ester | 14.558 | 0.374% |
| Total | 100 |
| Groups | Control | Diabetic | Diabetic + SL30 | Diabetic+SL90 | Diabetic+SL270 |
|---|---|---|---|---|---|
| Initial body weight (g) | 265 ± 12 | 252 ± 15 | 253 ± 22 | 244 ± 14 | 238 ± 18 |
| Final body weight (g) | 262 ± 51 a | 173 ± 5 b | 221 ± 40 a | 220 ± 19 a | 190 ± 3 b |
| Testis weight (g) | 1.4 ± 0.15 a | 0.85 ± 0.1 b | 0.97 ± 0.09 b | 1.42 ± 0.04 a | 1.56 ± 0.05 a |
| Groups | Control | Diabetic | Diabetic + SL30 | Diabetic+SL90 | Diabetic+SL270 |
|---|---|---|---|---|---|
| Testis volume | 1150.6 ± 110 a | 712.2 ± 34 b | 815.5 ± 12 b | 1108.4 ± 115 a | 1222 ± 108 a |
| Tubule volume | 948.3 ± 84 a | 605.1 ± 108 b | 732 ± 24 b | 918.5 ± 121 a | 1064.8 ± 119 a |
| Interstitial tissue volume | 126.2 ± 44 a | 98.8 ± 35 b | 81.2 ± 14 b | 117.9 ± 24 a | 131.8 ± 28 a |
| Epithelial height | 118.7 ± 12 a | 77.3 ± 9 b | 81.3 ± 24 b | 125.5 ± 32 a | 118.5 ± 16.5 a |
| Tubule diameter | 305.5 ± 31 a | 209.2 ± 12 b | 226.1 ± 8 b | 281.9 ± 34 a | 289 ± 41 a |
| Groups | Control | Diabetic | Diabetic + SL30 | Diabetic+SL90 | Diabetic+SL270 |
|---|---|---|---|---|---|
| Tubule length | 22.2 ± 3.1 a | 13.5 ± 0.8 b | 12.1 ± 1.5 b | 18.5 ± 7.1 a | 19.2 ± 4.7 a |
| Sperm tail length | 30.4 ± 4.8 a | 18.2 ± 7.8 b | 20.5 ± 4.2 b | 19.4 ± 5.8 b | 25.6 ± 2.9 a |
| Groups | Control | Diabetic | Diabetic + SL30 | Diabetic+SL90 | Diabetic+SL270 |
|---|---|---|---|---|---|
| Leydig cells | 6.2 ± 0.8 a | 4.2 ± 0.25 b | 4.8 ± 0.5 b | 4.6 ± 0.21 b | 5.9 ± 0.92 a |
| Sertoli cells | 13.7 ± 2.1 a | 7.5 ± 1.8 b | 8.2 ± 2.1 b | 7.6 ± 0.41 b | 15.3 ± 0.55 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kermani, J.; Goodarzi, N.; Bakhtiari, M. An Experimental Study to Evaluate the Protective Effects of Solanum lycopersicum Seed Essential Oil on Diabetes-Induced Testicular Injuries. Medicina 2019, 55, 499. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080499
Kermani J, Goodarzi N, Bakhtiari M. An Experimental Study to Evaluate the Protective Effects of Solanum lycopersicum Seed Essential Oil on Diabetes-Induced Testicular Injuries. Medicina. 2019; 55(8):499. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080499
Chicago/Turabian StyleKermani, Javid, Nader Goodarzi, and Mitra Bakhtiari. 2019. "An Experimental Study to Evaluate the Protective Effects of Solanum lycopersicum Seed Essential Oil on Diabetes-Induced Testicular Injuries" Medicina 55, no. 8: 499. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina55080499