
The Effect of Cholesterol on the Long-Range Network of Interactions Established among Sea Anemone Sticholysin II Residues at the Water-Membrane Interface

Abstract
:1. Introduction

2. Results
2.1. Protein Purification and Characterization

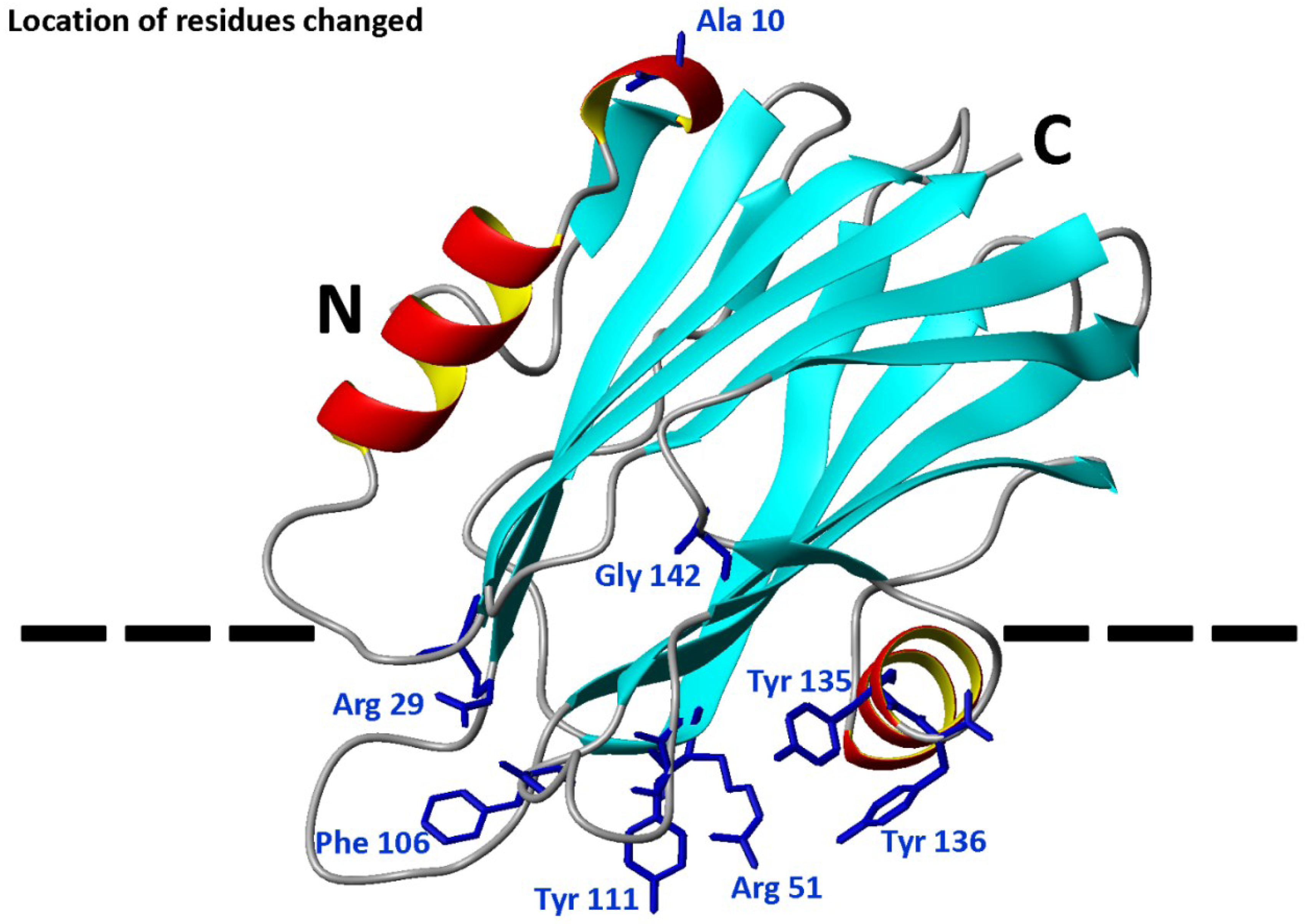{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
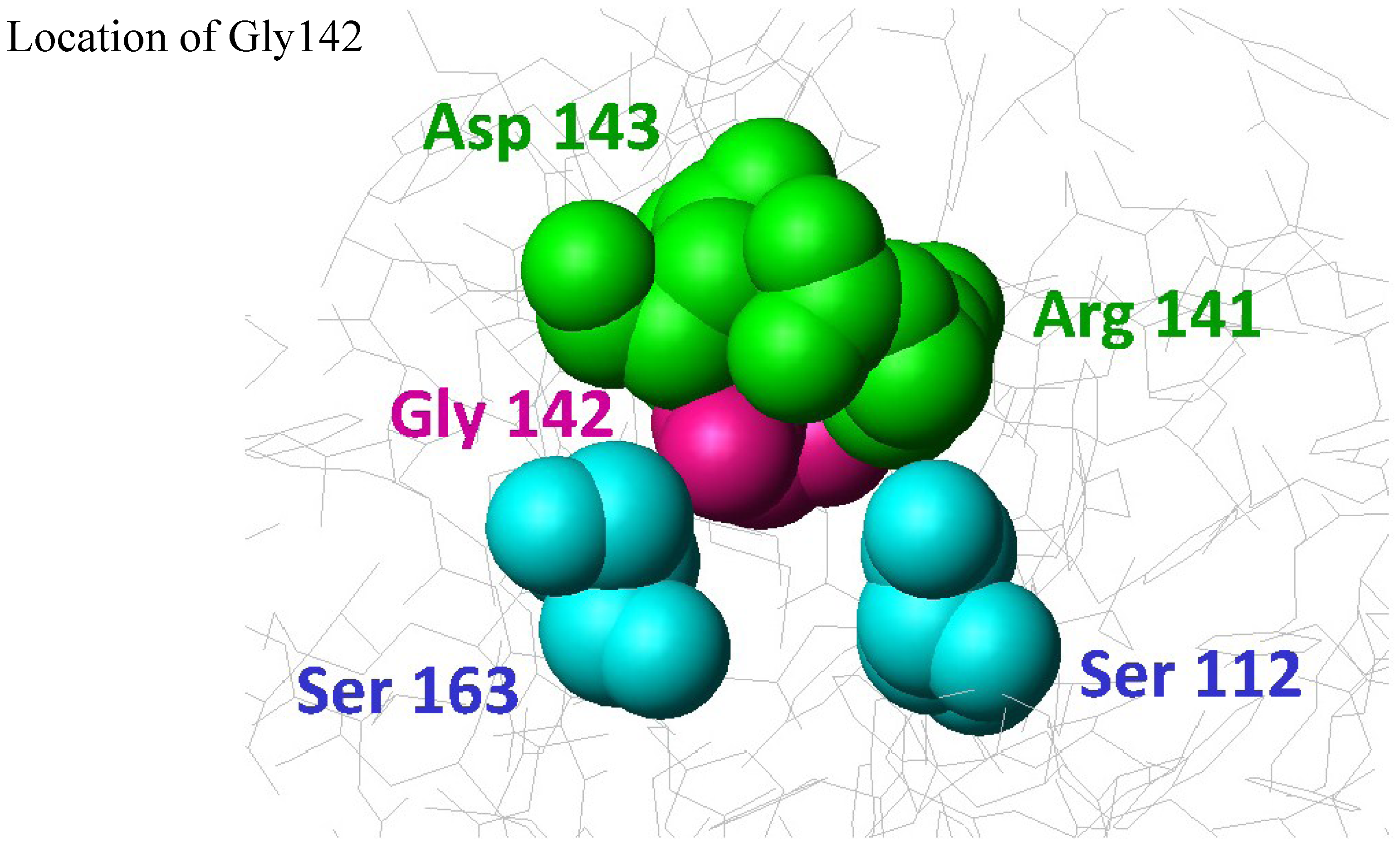{kind=link}
| StnII Variant | E0.1% (280 nm, 1 cm) | Tm (°C) | Relative Hemolytic Activity c |
|---|---|---|---|
| Wild-type | 2.54 a | 67 a | 1.00 |
| A10P | 2.69 a | 66 a | 0.26 a |
| R29Q | 2.54 a | 60 a | <0.10 a |
| R51Q | 2.38 | 67 | 0.30 |
| F106L | 2.62 a | 66 a | 0.39 a |
| Y111N | 2.58 a | 70 a | <0.10 a |
| Y135F | 2.47 | 66 | 0.26 |
| Y136F | 2.66 | 69 | 0.26 |
| G142A | 2.30 b | 61 b | 0.13 b |
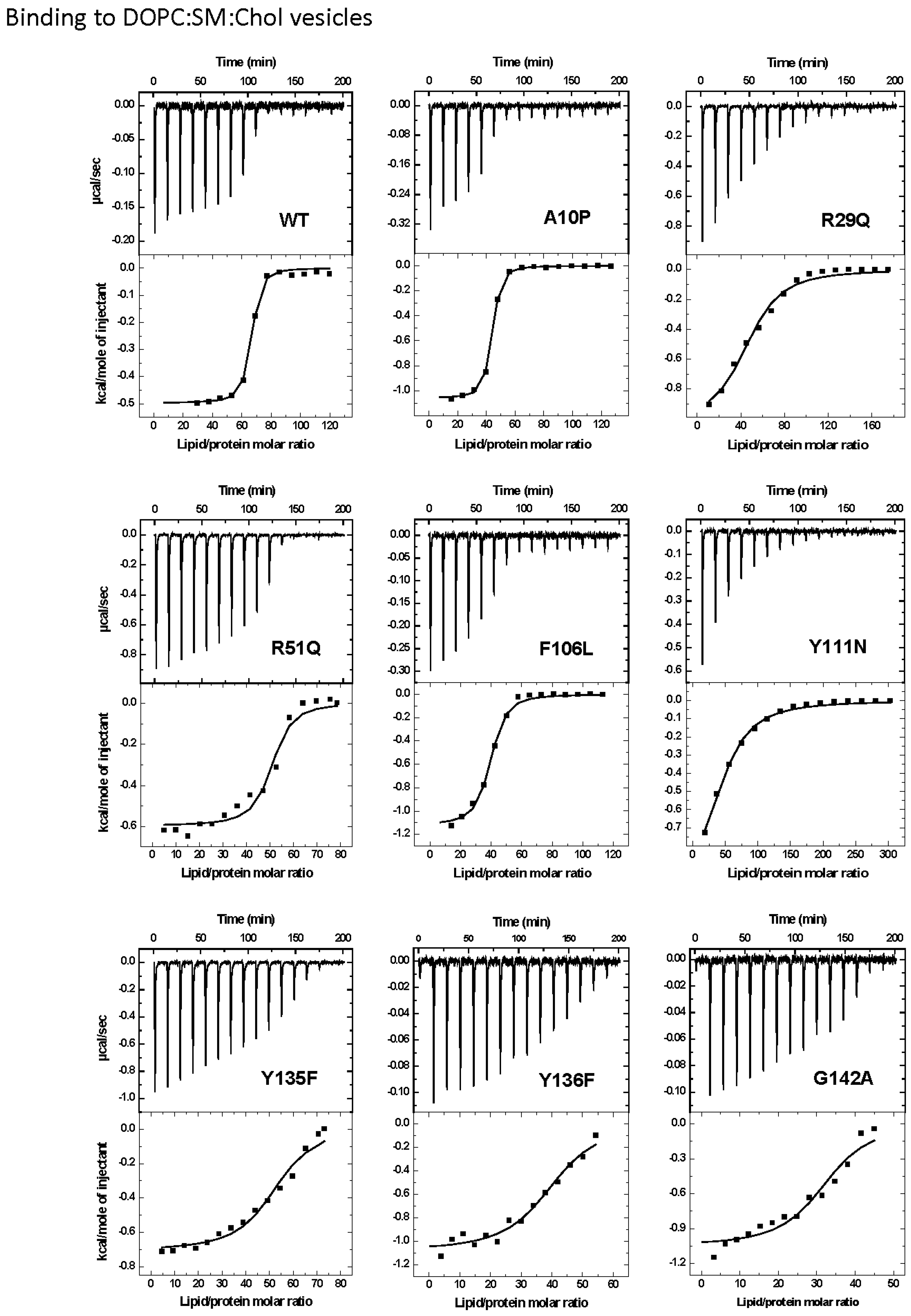
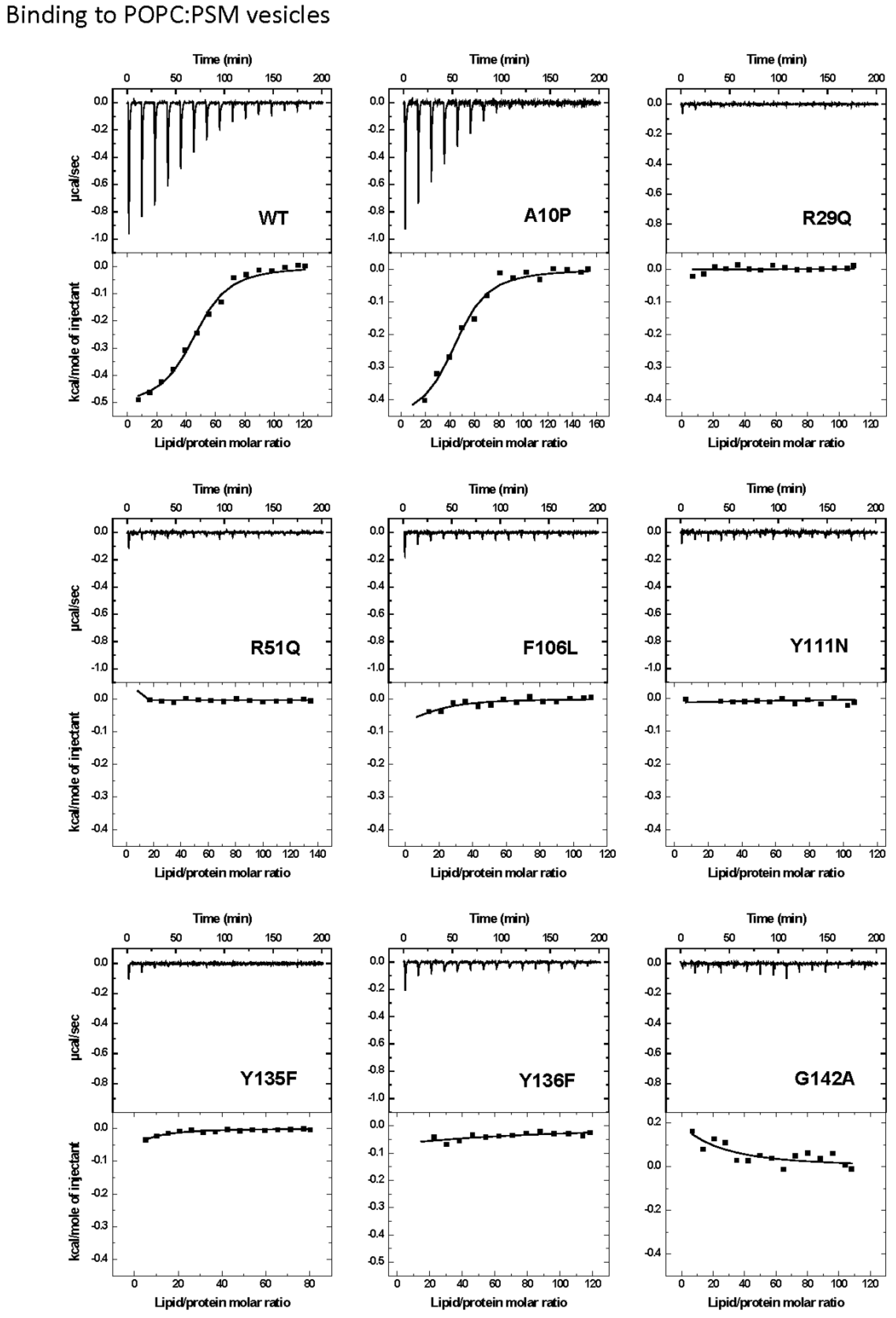
2.2. Protein Binding to the Two Different Lipid Model Vesicles Employed


| DOPC:SM:Chol (1:1:1) | ||||||
|---|---|---|---|---|---|---|
| StnII Variant | n | K × 10−8 (M−1) | ΔG (kcal/mol) | ΔH (kcal/mol) | ΔS (cal·mol−1·K−1) | Relative Membranebinding a |
| WT | 39 ± 4 b | 1.700 ± 0.900 b | −9.1 ± 0.5 b | −44.0 ± 3.0 b | −115.0 ± 9.0 b | 1.000 b |
| A10P | 37 ± 4 b | 1.900 ± 0.900 b | −9.3 ± 0.4 b | −39.0 ± 6.0 b | −99.0 ± 20.0 b | 1.180 b |
| R29Q | 51 ± 8 b | 0.031 ± 0.002 b | −6.7 ± 0.2 b | −45.0 ± 3.0 b | −129.0 ± 8.0 b | 0.014 b |
| R51Q | 49 ± 2 | 0.140 ± 0.070 | −7.6 ± 0.2 | −29.0 ± 1.0 | −72.0 ± 4.0 | 0.082 |
| F106L | 36 ± 3 b | 0.600 ± 0.100 b | −8.6 ± 0.1 b | −37.0 ± 7.0 b | −94.0 ± 24.0 b | 0.380 b |
| Y111N | 46 ± 7 b | 0.025 ± 0.004 b | −6.6 ± 0.1 b | −47.0 ± 2.0 b | −134.0 ± 4.0 b | 0.012 b |
| Y135F | 51 ± 2 | 0.039 ± 0.013 | −6.8 ± 0.2 | −36.0 ± 2.0 | −99.0 ± 6.0 | 0.023 |
| Y136F | 40 ± 1 | 0.270 ± 0.080 | −8.1 ± 0.1 | −43.0 ± 1.0 | −117.0 ± 4.0 | 0.160 |
| G142A | 32 ± 1 | 0.250 ± 0.100 | −8.2 ± 0.1 | −33.0 ± 1.0 | −83.0 ± 4.0 | 0.150 |
| POPC:PSM (4:1) | ||||||
| WT | 45 ± 2 | 1.5 ± 0.3 | −6.3 ± 0.1 | −23.0 ± 1.0 | −57.0 ± 3.0 | 1.000 |
| A10P | 44 ± 3 | 1.2 ± 0.4 | −6.2 ± 0.2 | −20.0 ± 2.0 | −48.0 ± 6.0 | 0.800 |
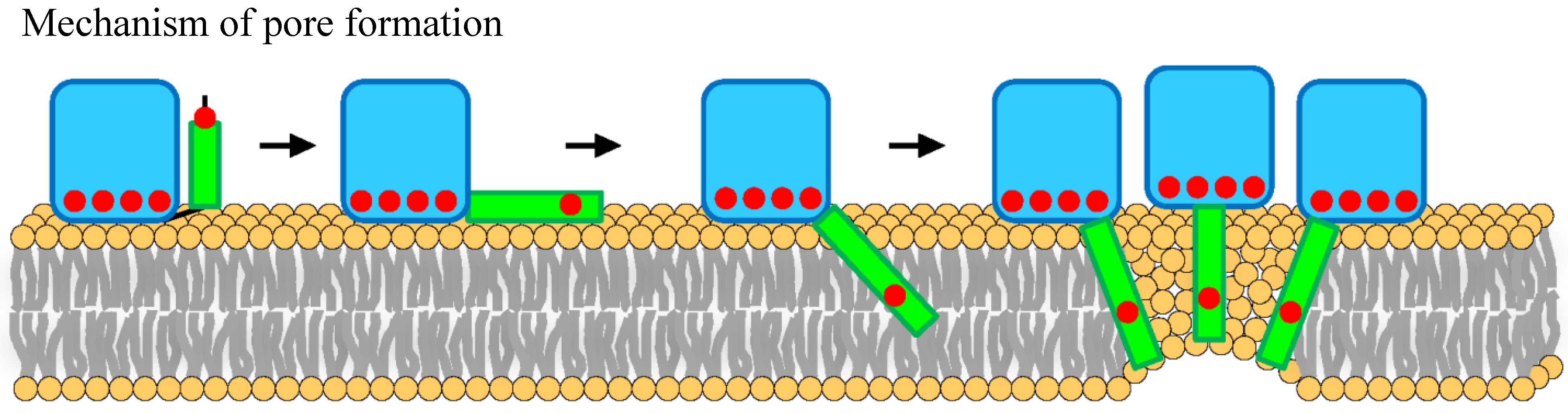
2.3. Pore Formation

3. Discussion

3.1. StnII Mutants Affecting the 30 First Residues Sequence Stretch
3.2. StnII Mutants Affecting the Aromatic Cluster
3.3. StnII Mutants Affecting the POC-Binding Site
3.4. StnII Mutant Affecting the Pore-Competent State of Protein Oligomerization

4. Experimental Section
4.1. Materials
4.2. Spectroscopic Characterization
4.3. Hemolysis Assay
4.4. Binding of StnII to Bilayer Membranes
4.5. Calcein Leakage Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Suput, D. In vivo effects of cnidarian toxins and venoms. Toxicon 2009, 54, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Macek, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, G.; Pederzolli, C.; Macek, P.; Menestrina, G. Pore formation by the sea anemone cytolysin equinatoxin-II in red blood cells and model lipid membranes. J. Membr. Biol. 1993, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Macek, P.; Belmonte, G.; Pederzolli, C.; Menestrina, G. Mechanism of action of equinatoxin II, a cytolysin from the sea anemone Actinia equina l. Belonging to the family of actinoporins. Toxicology 1994, 87, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Serra, M.D.; Ferreras, M.; Lanio, M.E.; Menestrina, G. Mechanism of membrane permeabilization by sticholysin I, a cytolysin isolated from the venom of the sea anemone Stichodactyla helianthus. Biochemistry 1996, 35, 14947–14957. [Google Scholar] [CrossRef] [PubMed]
- Basulto, A.; Pérez, V.M.; Noa, Y.; Varela, C.; Otero, A.J.; Pico, M.C. Immunohistochemical targeting of sea anemone cytolysins on tentacles, mesenteric filaments and isolated nematocysts of Stichodactyla helianthus. J. Exp. Zoolog. A Comp. Exp. Biol. 2006, 305, 253–258. [Google Scholar] [CrossRef]
- Tejuca, M.; Anderluh, G.; Macek, P.; Marcet, R.; Torres, D.; Sarracent, J.; Alvarez, C.; Lanio, M.E.; Dalla Serra, M.; Menestrina, G. Antiparasite activity of sea-anemone cytolysins on Giardia duodenalis and specific targeting with anti-Giardia antibodies. Int. J. Parasitol. 1999, 29, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Krizaj, I.; Strukelj, B.; Gubensek, F.; Macek, P.; Pungercar, J. Equinatoxins, pore-forming proteins from the sea anemone Actinia equina, belong to a multigene family. Toxicon 1999, 37, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- De los Ríos, V.; Oñaderra, M.; Martínez-Ruiz, A.; Lacadena, J.; Mancheño, J.M.; Martínez-del-Pozo, A.; Gavilanes, J.G. Overproduction in Escherichia coli and purification of the hemolytic protein sticholysin II from the sea anemone Stichodactyla helianthus. Protein Expr. Purif. 2000, 18, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Cebollada, J.; Oñaderra, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Sea anemone actinoporins: The transition from a folded soluble state to a functionally active membrane-bound oligomeric pore. Curr. Protein Pept. Sci. 2007, 8, 558–572. [Google Scholar] [CrossRef] [PubMed]
- García-Ortega, L.; Alegre-Cebollada, J.; García-Linares, S.; Bruix, M.; Martínez-del-Pozo, A.; Gavilanes, J.G. The behavior of sea anemone actinoporins at the water-membrane interface. Biochim. Biophys. Acta 2011, 1808, 2275–2288. [Google Scholar] [CrossRef] [PubMed]
- Bellomio, A.; Morante, K.; Barlic, A.; Gutiérrez-Aguirre, I.; Viguera, A.R.; Gonzalez-Mañas, J.M. Purification, cloning and characterization of fragaceatoxin c, a novel actinoporin from the sea anemone Actinia fragacea. Toxicon 2009, 54, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.; Moritz, R.L.; Simpson, R.J.; Norton, R.S. Tenebrosin-A, a new cardiostimulant protein from the australian sea anemone Actinia tenebrosa. Biochem. Int. 1987, 15, 711–718. [Google Scholar] [PubMed]
- Norton, R.S.; Bobek, G.; Ivanov, J.O.; Thomson, M.; Fiala-Beer, E.; Moritz, R.L.; Simpson, R.J. Purification and characterisation of proteins with cardiac stimulatory and haemolytic activity from the anemone Actinia tenebrosa. Toxicon 1990, 28, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Reid, G.E.; Moritz, R.L.; Morton, C.; Norton, R.S. Complete amino acid sequence of tenebrosin-C, a cardiac stimulatory and haemolytic protein from the sea anemone Actinia tenebrosa. Eur. J. Biochem. 1990, 190, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Monastyrnaya, M.M.; Zykova, T.A.; Apalikova, O.V.; Shwets, T.V.; Kozlovskaya, E.P. Biologically active polypeptides from the tropical sea anemone Radianthus macrodactylus. Toxicon 2002, 40, 1197–1217. [Google Scholar] [CrossRef] [PubMed]
- Antonini, V.; Perez-Barzaga, V.; Bampi, S.; Penton, D.; Martinez, D.; Dalla Serra, M.; Tejuca, M. Functional characterization of sticholysin I and W111C mutant reveals the sequence of the actinoporin’s pore assembly. PLoS ONE 2014, 9, e110824. [Google Scholar] [CrossRef] [PubMed]
- Penton, D.; Perez-Barzaga, V.; Diaz, I.; Reytor, M.L.; Campos, J.; Fando, R.; Calvo, L.; Cilli, E.M.; Morera, V.; Castellanos-Serra, L.R.; et al. Validation of a mutant of the pore-forming toxin sticholysin-I for the construction of proteinase-activated immunotoxins. Protein Eng. Des. Sel. 2011, 24, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Bakrac, B.; Anderluh, G. Molecular mechanism of sphingomyelin-specific membrane binding and pore formation by actinoporins. Adv. Exp. Med. Biol. 2009, 677, 106–115. [Google Scholar]
- Bernheimer, A.W.; Avigad, L.S. Properties of a toxin from the sea anemone Stoichactis helianthus, including specific binding to sphingomyelin. Proc. Natl. Acad. Sci. USA 1976, 73, 467–471. [Google Scholar] [CrossRef] [PubMed]
- De los Ríos, V.; Mancheño, J.M.; Lanio, M.E.; Oñaderra, M.; Gavilanes, J.G. Mechanism of the leakage induced on lipid model membranes by the hemolytic protein sticholysin II from the sea anemone Stichodactyla helianthus. Eur. J. Biochem. 1998, 252, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Barlic, A.; Gutiérrez-Aguirre, I.; Caaveiro, J.M.; Cruz, A.; Ruiz-Argüello, M.B.; Pérez-Gil, J.; González-Mañas, J.M. Lipid phase coexistence favors membrane insertion of equinatoxin-II, a pore-forming toxin from Actinia equina. J. Biol. Chem. 2004, 279, 34209–34216. [Google Scholar] [CrossRef] [PubMed]
- Bakrac, B.; Gutierrez-Aguirre, I.; Podlesek, Z.; Sonnen, A.F.; Gilbert, R.J.; Macek, P.; Lakey, J.H.; Anderluh, G. Molecular determinants of sphingomyelin specificity of a eukaryotic pore-forming toxin. J. Biol. Chem. 2008, 283, 18665–18677. [Google Scholar] [CrossRef]
- Maula, T.; Isaksson, Y.J.; García-Linares, S.; Niinivehmas, S.; Pentikainen, O.T.; Kurita, M.; Yamaguchi, S.; Yamamoto, T.; Katsumura, S.; Gavilanes, J.G.; et al. 2NH and 3OH are crucial structural requirements in sphingomyelin for sticholysin II binding and pore formation in bilayer membranes. Biochim. Biophys. Acta 2013, 1828, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Schön, P.; Garcia-Saez, A.J.; Malovrh, P.; Bacia, K.; Anderluh, G.; Schwille, P. Equinatoxin II permeabilizing activity depends on the presence of sphingomyelin and lipid phase coexistence. Biophys. J. 2008, 95, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Varanda, W.; Finkelstein, A. Ion and nonelectrolyte permeability properties of channels formed in planar lipid bilayer membranes by the cytolytic toxin from the sea anemone, Stoichactis helianthus. J. Membr. Biol. 1980, 55, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Alm, I.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Cholesterol stimulate and ceramide inhibit sticholysin II-induced pore formation in complex bilayer membranes. Biochim. Biophys. Acta Biomembr. 2014, 1848, 925–931. [Google Scholar] [CrossRef]
- Martínez, D.; Otero, A.; Alvarez, C.; Pazos, F.; Tejuca, M.; Lanio, M.E.; Gutierrez-Aguirre, I.; Barlic, A.; Iloro, I.; Arrondo, J.L.; et al. Effect of sphingomyelin and cholesterol on the interaction of St II with lipidic interfaces. Toxicon 2007, 49, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Cebollada, J.; Cunietti, M.; Herrero-Galán, E.; Gavilanes, J.G.; Martínez-del-Pozo, A. Calorimetric scrutiny of lipid binding by sticholysin II toxin mutants. J. Mol. Biol. 2008, 382, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Barenholz, Y.; Thompson, T.E. Sphingomyelins in bilayers and biological membranes. Biochim. Biophys. Acta 1980, 604, 129–158. [Google Scholar] [PubMed]
- Alegre-Cebollada, J.; Rodríguez-Crespo, I.; Gavilanes, J.G.; Martínez-del-Pozo, A. Detergent-resistant membranes are platforms for actinoporin pore-forming activity on intact cells. FEBS J. 2006, 273, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Patton, S. Correlative relationship of cholesterol and sphingomyelin in cell membranes. J. Theor. Biol. 1970, 29, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimaki, P.; Slotte, J.P. Cholesterol interactions with phospholipids in membranes. Prog. Lipid Res. 2002, 41, 66–97. [Google Scholar] [CrossRef] [PubMed]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadis, A.; Anderluh, G.; Macek, P.; Turk, D. Crystal structure of the soluble form of equinatoxin II, a pore-forming toxin from the sea anemone Actinia equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Malovrh, P.; Viero, G.; Serra, M.D.; Podlesek, Z.; Lakey, J.H.; Macek, P.; Menestrina, G.; Anderluh, G. A novel mechanism of pore formation: Membrane penetration by the N-terminal amphipathic region of equinatoxin. J. Biol. Chem. 2003, 278, 22678–22685. [Google Scholar] [CrossRef] [PubMed]
- Rojko, N.; Kristan, K.C.; Viero, G.; Zerovnik, E.; Macek, P.; Dalla Serra, M.; Anderluh, G. Membrane damage by an alpha-helical pore-forming protein, equinatoxin II, proceeds through a succession of ordered steps. J. Biol. Chem. 2013, 288, 23704–23715. [Google Scholar] [CrossRef] [PubMed]
- Rojko, N.; Cronin, B.; Danial, J.S.; Baker, M.A.; Anderluh, G.; Wallace, M.I. Imaging the lipid-phase-dependent pore formation of equinatoxin II in droplet interface bilayers. Biophys. J. 2014, 106, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Rojko, N.; Cronin, B.; Anderluh, G.; Wallace, M.I. Photobleaching reveals heterogeneous stoichiometry for equinatoxin II oligomers. Chembiochem 2014, 15, 2139–2145. [Google Scholar] [CrossRef] [PubMed]
- Subburaj, Y.; Ros, U.; Hermann, E.; Tong, R.; García-Sáez, A.J. Toxicity of an α-pore-forming toxin depends on the assembly mechanism on the target membrane as revealed by single-molecule imaging. J. Biol. Chem. 2015, 290, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Mechaly, A.E.; Bellomio, A.; Gil-Carton, D.; Morante, K.; Valle, M.; Gonzalez-Mañas, J.M.; Guerin, D.M. Structural insights into the oligomerization and architecture of eukaryotic membrane pore-forming toxins. Structure 2011, 19, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Koradi, R.; Billeter, M.; Wüthrich, K. MolMol: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 29–32, 51–55. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Lacadena, V.; Oñaderra, M.; Mancheño, J.M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Phenotypic selection and characterization of randomly produced non-haemolytic mutants of the toxic sea anemone protein sticholysin II. FEBS Lett. 2004, 575, 14–18. [Google Scholar] [CrossRef] [PubMed]
- García-Linares, S.; Richmond, R.; García-Mayoral, M.F.; Bustamante, N.; Bruix, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. The sea anemone actinoporin (Arg-Gly-Asp) conserved motif is involved in maintaining the competent oligomerization state of these pore-forming toxins. FEBS J. 2014, 281, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, T.K.; Lindroos, D.; Westerlund, B.; Slotte, J.P. Construction of a dopc/psm/cholesterol phase diagram based on the fluorescence properties of trans-parinaric acid. Langmuir 2011, 27, 8339–8350. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, R.F.; Fedorov, A.; Prieto, M. Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: Boundaries and composition of lipid rafts. Biophys. J. 2003, 85, 2406–2416. [Google Scholar] [CrossRef] [PubMed]
- Veatch, S.L.; Keller, S.L. Miscibility phase diagrams of giant vesicles containing sphingomyelin. Phys. Rev. Lett. 2005, 94, 148101. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Cea, M.A.; Castrillo, I.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Bruix, M. Intrinsic local disorder and a network of charge-charge interactions are key to actinoporin membrane disruption and cytotoxicity. FEBS J. 2011, 278, 2080–2089. [Google Scholar] [CrossRef] [PubMed]
- García-Linares, S.; Castrillo, I.; Bruix, M.; Menéndez, M.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G. Three-dimensional structure of the actinoporin sticholysin I. Influence of long-distance effects on protein function. Arch. Biochem. Biophys. 2013, 532, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Goormaghtigh, E. Infrared spectroscopy study on the conformational changes leading to pore formation of the toxin sticholysin II. Biophys. J. 2007, 93, 3191–3201. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.J.; Elliott, J.L.; Collier, R.J. Anthrax protective antigen: Prepore-to-pore conversion. Biochemistry 1999, 38, 10432–10441. [Google Scholar] [CrossRef] [PubMed]
- Heuck, A.P.; Tweten, R.K.; Johnson, A.E. Assembly and topography of the prepore complex in cholesterol-dependent cytolysins. J. Biol. Chem. 2003, 278, 31218–31225. [Google Scholar] [CrossRef] [PubMed]
- Kristan, K.; Podlesek, Z.; Hojnik, V.; Gutiérrez-Aguirre, I.; Guncar, G.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; Macek, P.; Anderluh, G. Pore formation by equinatoxin, a eukaryotic pore-forming toxin, requires a flexible N-terminal region and a stable β-sandwich. J. Biol. Chem. 2004, 279, 46509–46517. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Gutiérrez-Aguirre, I.; Barlic, A.; Malovrh, P.; Kristan, K.; Podlesek, Z.; Macek, P.; Turk, D.; Gonzalez-Mañas, J.M.; Lakey, J.H.; et al. Two-step membrane binding by equinatoxin II, a pore-forming toxin from the sea anemone, involves an exposed aromatic cluster and a flexible helix. J. Biol. Chem. 2002, 277, 41916–41924. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, I.; Araujo, N.A.; Alegre-Cebollada, J.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Bruix, M. Specific interactions of sticholysin I with model membranes: An nmr study. Proteins 2010, 78, 1959–1970. [Google Scholar] [PubMed]
- Hinds, M.G.; Zhang, W.; Anderluh, G.; Hansen, P.E.; Norton, R.S. Solution structure of the eukaryotic pore-forming cytolysin equinatoxin II: Implications for pore formation. J. Mol. Biol. 2002, 315, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Cea, M.A.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Bruix, M. 1h, 13C, and 15NNMR assignments of stnII-Y111N, a highly impaired mutant of the sea anemone actinoporin sticholysin ii. Biomol. NMR Assign. 2010, 4, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Jaikishan, S.; Björkbom, A.; Slotte, J.P. Sphingomyelin analogs with branched n-acyl chains: The position of branching dramatically affects acyl chain order and sterol interactions in bilayer membranes. Biochim. Biophys. Acta 2010, 1798, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Terová, B.; Heczko, R.; Slotte, J.P. On the importance of the phosphocholine methyl groups for sphingomyelin/cholesterol interactions in membranes: A study with ceramide phosphoethanolamine. Biophys. J. 2005, 88, 2661–2669. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Alegre-Cebollada, J.; Clementi, G.; Cunietti, M.; Porres, C.; Oñaderra, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Silent mutations at the 5'-end of the cDNA of actinoporins from the sea anemone Stichodactyla helianthus allow their heterologous overproduction in Escherichia coli. J. Biotechnol. 2007, 127, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Mancheño, J.M.; de los Ríos, V.; Martínez-del-Pozo, A.; Lanio, M.E.; Oñaderra, M.; Gavilanes, J.G. Partially folded states of the cytolytic protein sticholysin II. Biochim. Biophys. Acta 2001, 1545, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-García, E.; Martínez-del-Pozo, A.; Gavilanes, J.G. Role of the basic character of α-sarcin’s NH2-terminal β-hairpin in ribosome recognition and phospholipid interaction. Arch. Biochem. Biophys. 2009, 481, 37–44. [Google Scholar] [CrossRef] [PubMed]
- De Antonio, C.; Martínez-del-Pozo, A.; Mancheño, J.M.; Oñaderra, M.; Lacadena, J.; Martínez-Ruiz, A.; Pérez-Cañadillas, J.M.; Bruix, M.; Gavilanes, J.G. Assignment of the contribution of the tryptophan residues to the spectroscopic and functional properties of the ribotoxin α-sarcin. Proteins 2000, 41, 350–361. [Google Scholar] [CrossRef] [PubMed]
- De los Ríos, V.; Mancheño, J.M.; Martínez-del-Pozo, A.; Alfonso, C.; Rivas, G.; Oñaderra, M.; Gavilanes, J.G. Sticholysin II, a cytolysin from the sea anemone Stichodactyla helianthus, is a monomer-tetramer associating protein. FEBS Lett. 1999, 455, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Menestrina, G.; Cabiaux, V.; Tejuca, M. Secondary structure of sea anemone cytolysins in soluble and membrane bound form by infrared spectroscopy. Biochem. Biophys. Res. Commun. 1999, 254, 174–180. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Linares, S.; Alm, I.; Maula, T.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, Á. The Effect of Cholesterol on the Long-Range Network of Interactions Established among Sea Anemone Sticholysin II Residues at the Water-Membrane Interface. Mar. Drugs 2015, 13, 1647-1665. https://0-doi-org.brum.beds.ac.uk/10.3390/md13041647
García-Linares S, Alm I, Maula T, Gavilanes JG, Slotte JP, Martínez-del-Pozo Á. The Effect of Cholesterol on the Long-Range Network of Interactions Established among Sea Anemone Sticholysin II Residues at the Water-Membrane Interface. Marine Drugs. 2015; 13(4):1647-1665. https://0-doi-org.brum.beds.ac.uk/10.3390/md13041647
Chicago/Turabian StyleGarcía-Linares, Sara, Ida Alm, Terhi Maula, José G. Gavilanes, Johan Peter Slotte, and Álvaro Martínez-del-Pozo. 2015. "The Effect of Cholesterol on the Long-Range Network of Interactions Established among Sea Anemone Sticholysin II Residues at the Water-Membrane Interface" Marine Drugs 13, no. 4: 1647-1665. https://0-doi-org.brum.beds.ac.uk/10.3390/md13041647