New Benthic Cyanobacteria from Guadeloupe Mangroves as Producers of Antimicrobials

 , , , and
, , , and
Abstract
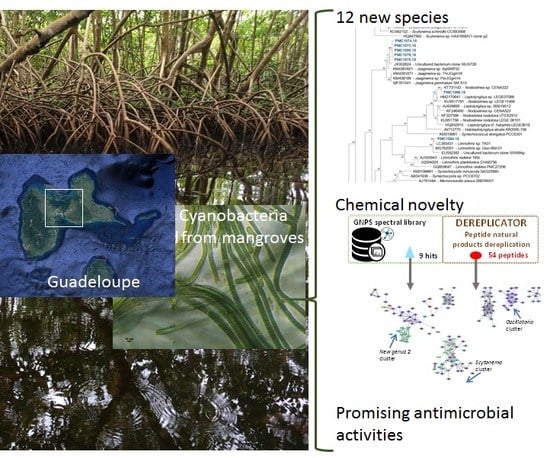
:
1. Introduction
2. Results
2.1. Phylogenetic and 16S rRNA Dissimilarity-Based Identification of Strains
2.2. MS/MS Analysis and Annotation of Cyanobacterial Specialized Metabolites
2.3. Evaluation of the Antimicrobial Activity
3. Discussion and Conclusions
3.1. Mangroves of Guadeloupe are a Source of Novel Cyanobacteria
3.2. The Benthic Cyanobacterial Strains Reveal a High Level of Chemical Novelty
3.3. Certain Cyanobacterial Strains Reveal Promising Antimicrobial Activities
4. Materials and Methods
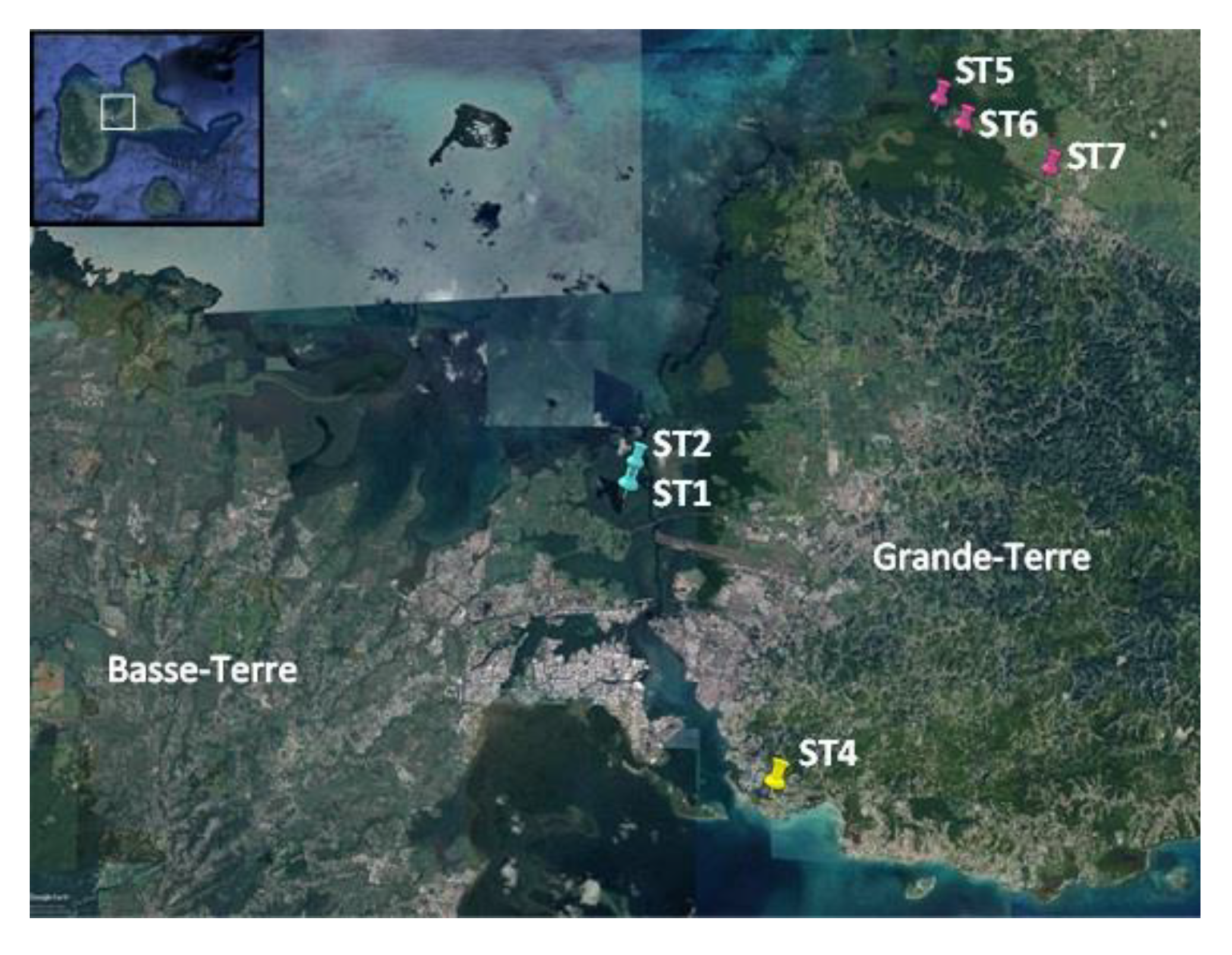
4.1. Sampling and Strain Isolation
4.2. Strain Cultivation and Biomass Production
4.3. Strain Identification
4.4. Preparation of Cyanobacteria Chemical Extracts
4.5. LC-MS2 Analyzes of Extracts
4.5.1. MS/MS Data Pretreatment
4.5.2. Molecular Networks Generation
4.6. Evaluation of the Antibacterial Activity of Cyanobacterial Strains
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shah, S.A.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.-W. Structural diversity, biological properties and applications of natural products from Cyanobacteria. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, D.O.; Rigonato, J.; Branco, L.H.Z.; Fiore, M.F. Cyanobacteria in mangrove ecosystems. Biodivers. Conserv. 2015, 24, 799–817. [Google Scholar] [CrossRef]
- Lovelock, C.; Grinham, A.; Adame, F.; Penros, H. Elemental composition and productivity of cyanobacterial mats in an arid zone estuary in north Western Australia. Wetl. Ecol. Manag. 2010, 18, 37–47. [Google Scholar] [CrossRef]
- Kleigrewe, K.; Almaliti, J.; Tian, I.Y.; Kinnel, R.B.; Korobeynikov, A.; Monroe, E.A.; Duggan, B.M.; Di Marzo, V.; Sherman, D.H.; Dorrestein, P.C.; et al. Combining Mass Spectrometric Metabolic Profiling with Genomic Analysis: A Powerful Approach for Discovering Natural Products from Cyanobacteria. J. Nat. Prod. 2015, 78, 1672–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [Green Version]
- Mi, Y.; Zhang, J.; He, S.; Yan, X. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef] [Green Version]
- Komarek, J.; Kastovsky, J.; Mares, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Guidi-Rontani, C.; Jean, M.R.N.; Gonzalez-Rizzo, S.; Bolte-Kluge, S.; Gros, O. Description of new filamentous toxic Cyanobacteria (Oscillatoriales) colonizing the sulfidic periphyton mat in marine mangroves. FEMS Microbiol. Lett. 2014, 359, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Fox Ramos, A.E.; Evanno, L.; Poupon, E.; Champy, P.; Beniddir, M.A. Natural products targeting strategies involving molecular networking: Different manners, one goal. Nat. Prod. Rep. 2019, 36, 960–980. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Alvarenga, D.O.; Rigonata, J.; Branco, L.E.; Mele, I.S.; Fiore, M.F. Phyllonema aviceniicola gen. nov., sp nov and Foliisarcina bertiogensis gen. nov., sp nov., epiphyllic cyanobacteria associated with Avicennia schaueriana leaves. Int. J. Syst. Evol. Microbiol. 2016, 66, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohimani, H.; Gurevich, A.; Mikheenko, A.; Garg, N.; Nothias, L.-F.; Ninomiya, A.; Takada, K.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of peptidic natural products through database search of mass spectra. Nat. Chem. Biol. 2016, 13, 30–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, H.C.; Ibá#xF1;ez, R.; Nothias, L.-F.; Boya, P.C.A.; Reinert, L.K.; Rollins-Smith, L.A.; Dorrestein, P.C.; Gutiérrez, M. Viscosin-like lipopeptides from frog skin bacteria inhibit Aspergillus fumigatus and Batrachochytrium dendrobatidis detected by imaging mass spectrometry and molecular networking. Sci. Rep. 2019, 9, 3019. [Google Scholar] [CrossRef]
- Saurav, K.; Macho, M.; Kust, A.; Delawská, K.; Hájek, J.; Hrouzek, P. Antimicrobial activity and bioactive profiling of heterocytous cyanobacterial strains using MS/MS-based molecular networking. Folia Microbiol. 2019, 64, 645–654. [Google Scholar] [CrossRef]
- Paulo, B.S.; Sigrist, R.; Angolini, C.F.F.; De Oliveira, L.G. New Cyclodepsipeptide Derivatives Revealed by Genome Mining and Molecular Networking. Chem. Sel. 2019, 4, 7785–7790. [Google Scholar] [CrossRef]
- Olivon, F.; Allard, P.-M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.-F.; Cachet, X.; et al. Bioactive Natural Products Prioritization Using Massive Multi-informational Molecular Networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef]
- Cellamare, M.; Duval, C.; Drelin, Y.; Djediat, C.; Touibi, N.; Agogué, H.; Leboulanger, C.; Ader, M.; Bernard, C. Characterization of phototrophic microorganisms and description of new cyanobacteria isolated from the saline-alkaline crater-lake Dziani Dzaha (Mayotte, Indian Ocean). FEMS Microbiol. Ecol. 2018, 94, 108. [Google Scholar] [CrossRef]
- Rigonato, J.; Alvarenga, D.O.; Andreote, F.; Dias, A.C.; Melo, I.S.; Kent, A.; Fiore, M.F. Cyanobacterial diversity in the phyllosphere of a mangrove forest. FEMS Microbiol. Ecol. 2012, 80, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Zubia, M.; Turquet, J.; Golubic, S. Benthic cyanobacterial diversity of Iles Eparses (Scattered Islands) in the Mozambique Channel. Acta Oecol. 2016, 72, 21–32. [Google Scholar] [CrossRef]
- Zubia, M.; Vieira, C.; Palinska, K.A.; Roue, M.; Gaertner, J.C.; Zloch, I.; Grellier, M.; Golubic, S. Benthic cyanobacteria on coral reefs of Moorea Island (French Polynesia): Diversity response to habitat quality. Hybrobiologia 2019, 843, 61–78. [Google Scholar] [CrossRef]
- Beck, C.; Knoop, H.; Steuer, R. Modules of co-occurrence in the cyanobacterial pan-genome reveal functional associations between groups of ortholog genes. PLoS Genet. 2018, 14, e1007239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, D.O.; Fiore, M.F.; Varani, A.M. A metagenomic approach to cyanobacterial genomics. Front. Microbiol. 2017, 8, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontharet, S.; Crémière, A.; Blanc-Valleron, M.M.; Sébilo, M.; Gros, O.; Laverman, A.M.; Dessailly, D. Sediment characteristics and microbial mats in a marine mangrove, Manche-à-eau lagoon (Guadeloupe). J. Soils Sediments 2017, 17, 1999–2010. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and purification of cyanobacteria. Meth. Enzym. 1988, 167, 3–27. [Google Scholar]
- Lane, D.L. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackenbrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Gugger, M.; Lyra, C.; Henriksen, P.; Couté, A.; Humbert, J.-F.; Sivonen, K. Phylogenetic comparison of the cyanobacterial genera Anabaena and Aphanizomenon. Int. J. Syst. Evol. Microbiol. 2002, 52, 1867–1880. [Google Scholar]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of RNA alignments. Bioinformatics 2009, 10, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Order | Affiliation | Sampling Site | Coordinates | Isolation Source | Best BLAST Hit and % 16S rRNA Sequence Similarity |
|---|---|---|---|---|---|---|
| PMC 1050.18 | Oscillatoriales | Oscillatoria n. sp. 3 | Manche-à-Eau, ST1 | 16°16′32″ N/61°33′18″ W | Dense filamentous brown mat | Oscillatoria/Kamptonema formosum BDU 92022 (KU958133)/99% |
| PMC 1051.18 | Oscillatoriales | Oscillatoria n. sp. 3 | Dense filamentous brown mat | Oscillatoria/Kamptonema formosum BDU 92022 (KU958133)/99% | ||
| PMC 1052.18 | Synechococcales | Gen. Nov. 3, n. sp. 1 | Benthic mat with Cyanobacteria and Beggiatoa-like morphotypes | Synechocystis sp. (KU951820)/92% | ||
| PMC 1056.18 | Oscillatoriales | Arthrospira, n. sp. 1 | Manche-à-Eau, ST2 | 16°16′49″ N/61°33′13″ W | Benthic blue-green mat | Uncultured bacterium clone TV002_28 (JX521753)/98% |
| PMC 1057.18 | Oscillatoriales | Arthrospira, n. sp. 1 | Periphytic biofilm covering immersed roots of Rhizophora mangle | Uncultured bacterium clone TV002_28 (JX521753)/97% | ||
| PMC 1064.18 | Synechococcales | Limnothrix n. sp. 1 | Marina Bas du Fort, ST4 | 16°13′13″ N/61°31′24″ W | Dense green benthic biofilm containing Oscillatoriales, Spirulina and Beggiatoa-like morphotypes | Limnothrix sp. TK01 (LC383431)/99% |
| PMC 1066.18 | Synechococcales | Nodosilinea n. sp. 1 | Canal des Rotours, ST5 | 16°21′8.4″ N/61°29′20″ W | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Nodosilinea sp. CENA322 (KT731143)/99% |
| PMC 1068.18 | Oscillatoriales | Oscillatoria n. sp. 2 | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Oscillatoria/Kamptonema formosum BDU 92022 (KU958133)/98% | ||
| PMC 1069.18 | Nostocales | Scytonema n. sp. 1 | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Scytonema cf. mirabile ER0515.01 (MG970546)/96% | ||
| PMC 1070.18 | Nostocales | Scytonema n. sp. 1 | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Scytonema cf. mirabile ER0515.01 (MG970546)/96% | ||
| PMC 1071.18 | Oscillatoriales | Gen. Nov. 1, n. sp. 1 | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Symploca sp. NAC 12/21/08-3 (JQ388601)/92% | ||
| PMC 1072.18 | Oscillatoriales | Gen. Nov. 2, n. sp. 1 | Dense periphytic biofilm covering immersed roots of Rhizophora mangle | Uncultured bacterium clone DM1-166 (KC329581)/94% | ||
| PMC 1073.18 | Synechococcales | Jaaginema n. sp. 1 | Canal des Rotours, ST 6 | 16°20′50″ N/61°29′03″ W | Dense periphytic blue-green biofilm covering immersed roots of Rhizophora nigra | Jaaginema sp. PsrJGgm14 (KM438189)/98% |
| PMC 1074.18 | Synechococcales | Jaaginema n. sp. 1 | Dense periphytic blue-green biofilm covering immersed roots of Rhizophora nigra | Jaaginema sp. PsrJGgm14 (KM438189)/91% | ||
| PMC 1075.18 | Oscillatoriales | Oscillatoria n. sp. 1 | Dense periphytic blue-green biofilm covering immersed roots of Rhizophora nigra | Oscillatoria/Kamptonema formosum BDU 92022 (KU958133)/94% | ||
| PMC 1076.18 | Oscillatoriales | Oscillatoria n. sp. 2 | Canal des Rotours, ST7 | 16°20′19″ N/61°27′55″ W | Dense periphytic biofilm covering immersed branch fragment | Oscillatoria/Kamptonema formosum BDU 92022 (KU958133)/99% |
| PMC 1078.18 | Synechococcales | Jaaginema n. sp. 1 | Dense periphytic biofilm covering immersed branch fragment | Jaaginema sp. PsrJGgm14 (KM438189)/98% | ||
| PMC 1079.18 | Synechococcales | Jaaginema n. sp. 1 | Dense periphytic biofilm covering immersed branch fragment | Jaaginema sp. PsrJGgm14 (KM438189)/98% | ||
| PMC 1080.18 | Synechococcales | Jaaginema n. sp. 1 | Dense periphytic biofilm covering immersed branch fragment | Jaaginema sp. PsrJGgm14 (KM438189)/97% | ||
| PMC 1092.19 | Oscillatoriales | Lyngbya, n. sp. 1 | Dense periphytic biofilm covering immersed branch fragment | Lyngbya sp. ALCB114379 (KY824052)/98% |
| Strain ID | Gram-Negative Pathogenic Bacteria | |
|---|---|---|
| Human E. coli | Environmental P. atlantica | |
| Oscillatoria spp. | ||
| PMC 1051.18 | 100 ± 4.17 | n.a. |
| PMC 1076.18 | n.a. | 50.87 ± 1.37 |
| Jaaginema sp. | ||
| PMC 1078.18 | n.a. | 54.07 ± 8.50 |
| PMC 1079.18 | n.a. | 60.95 ± 5.52 |
| PMC 1080.18 | n.a. | 53.01 ± 4.42 |
| Variant PNP | Producer | Detected in | Score | P-Value | Variant PNP Mass | Peptide Mass * |
|---|---|---|---|---|---|---|
| Viequeamide A | Marine Button Cyanobacterium (unidentified) | Oscillatoria sp., Scytonema sp., Arthrospira sp. | 12 | 2.8 10−17 | 803.50 | 892.55 |
| Nostophycin | Nostoc 152 | Lyngbya sp. | 11 | 2.1 10−15 | 888.48 | 943.55 |
| Wewakamide A | Lyngbya semiplena Lyngbya majuscula | Oscillatoria sp., Scytonema sp., Arthrospira sp. | 12 | 4.4 10−15 | 994.65 | 891.59 |
| Majusculamide C_Demethoxy | Lyngbya majuscula | Oscillatoria sp., Scytonema sp., Arthrospira sp. | 11 | 4.6 10−13 | 954.58 | 892.49 |
| Wewakazole | Lyngbya majuscula | Lyngbya sp. | 12 | 6.7 10−12 | 1140.54 | 1058.54 |
| Anacyclamide A10 | Anabaena sp. 90 | Lyngbya sp. | 10 | 9.6 10−11 | 1052.53 | 1009.49 |
| Aerucyclamide C | Microcystis aeruginosa | Lyngbya sp. | 7 | 9.5 10−9 | 516.22 | 645.28 |
| Microcystin LR | Microcystis bloom | Lyngbya sp. | 7 | 1.5 10−8 | 980.53 | 846.91 |
| Pitipeptolide A | Lyngbya majuscula | Oscillatoria sp., Scytonema sp., Arthrospira sp. | 7 | 1.6 10−8 | 807.48 | 795.51 |
| Microcystin RA | Microcystis | Lyngbya sp. | 7 | 1.7 10−8 | 952.50 | 825.38 |
| Raocyclamide B | Oscillatoria raoi | Arthrospira sp. | 6 | 6.5 10−8 | 568.21 | 581.17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duperron, S.; Beniddir, M.A.; Durand, S.; Longeon, A.; Duval, C.; Gros, O.; Bernard, C.; Bourguet-Kondracki, M.-L. New Benthic Cyanobacteria from Guadeloupe Mangroves as Producers of Antimicrobials. Mar. Drugs 2020, 18, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/md18010016
Duperron S, Beniddir MA, Durand S, Longeon A, Duval C, Gros O, Bernard C, Bourguet-Kondracki M-L. New Benthic Cyanobacteria from Guadeloupe Mangroves as Producers of Antimicrobials. Marine Drugs. 2020; 18(1):16. https://0-doi-org.brum.beds.ac.uk/10.3390/md18010016
Chicago/Turabian StyleDuperron, Sébastien, Mehdi A. Beniddir, Sylvain Durand, Arlette Longeon, Charlotte Duval, Olivier Gros, Cécile Bernard, and Marie-Lise Bourguet-Kondracki. 2020. "New Benthic Cyanobacteria from Guadeloupe Mangroves as Producers of Antimicrobials" Marine Drugs 18, no. 1: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/md18010016