
Expression and Characterization of a Novel Cold-Adapted Chitosanase from Marine Renibacterium sp. Suitable for Chitooligosaccharides Preparation

Abstract
:1. Introduction
2. Results
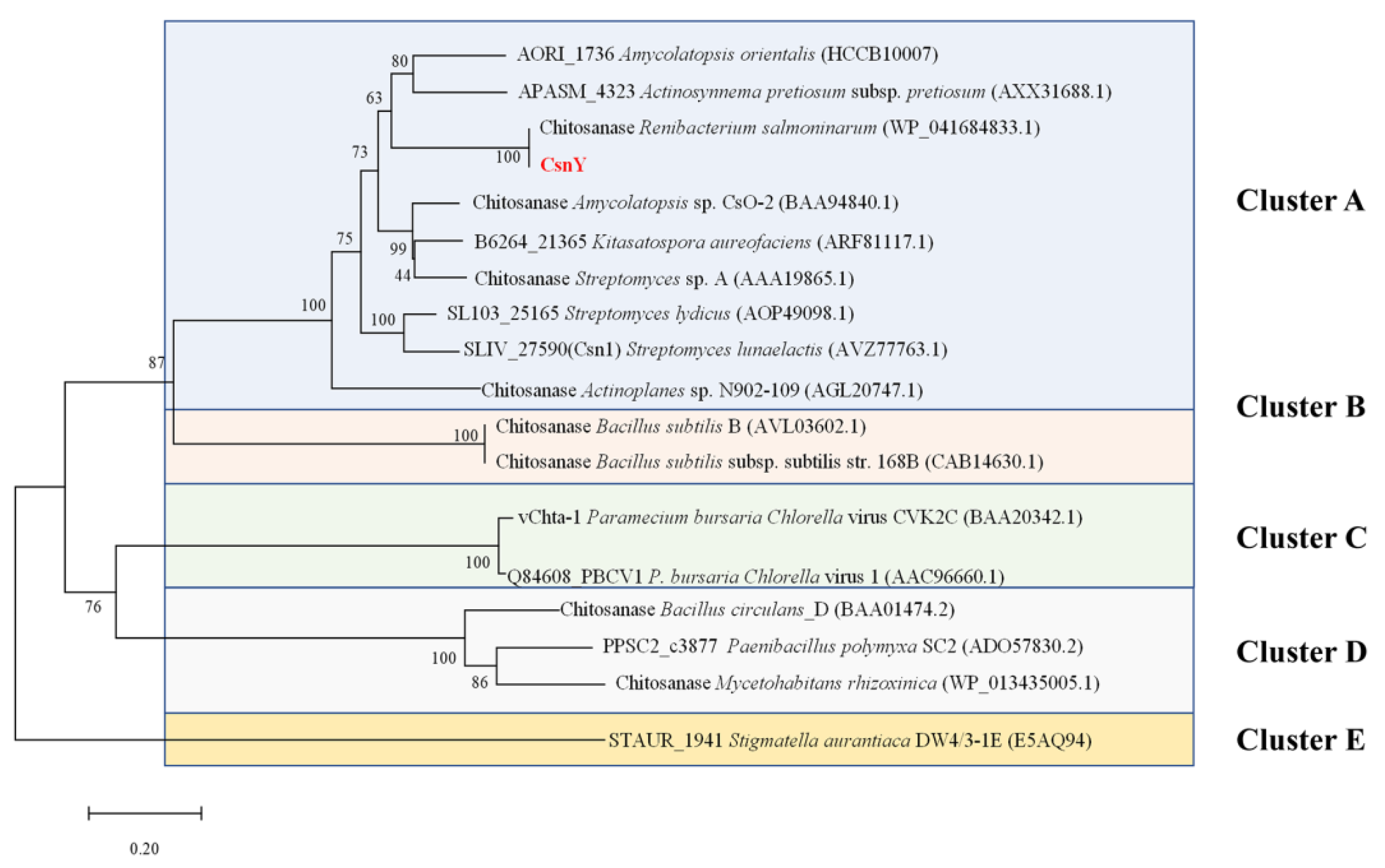
2.1. CsnY Sequence Analysis
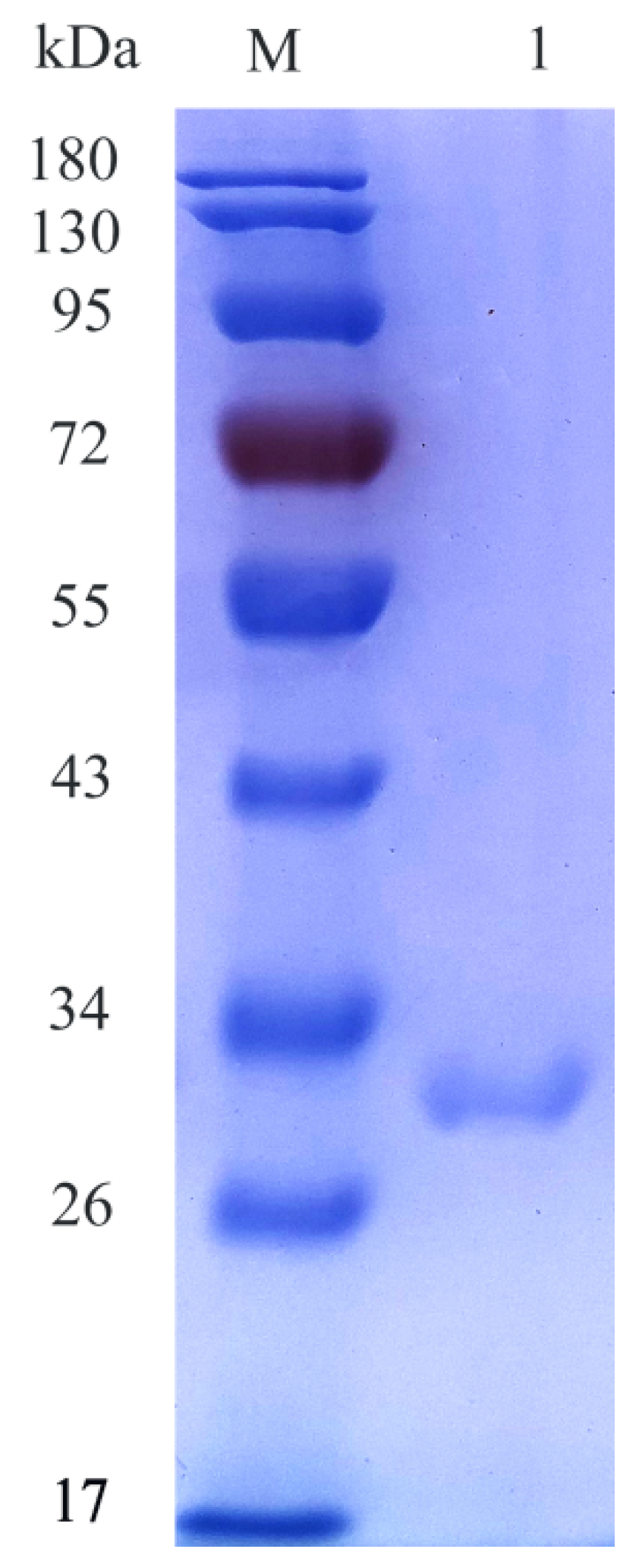
2.2. Expression and Purification of CsnY
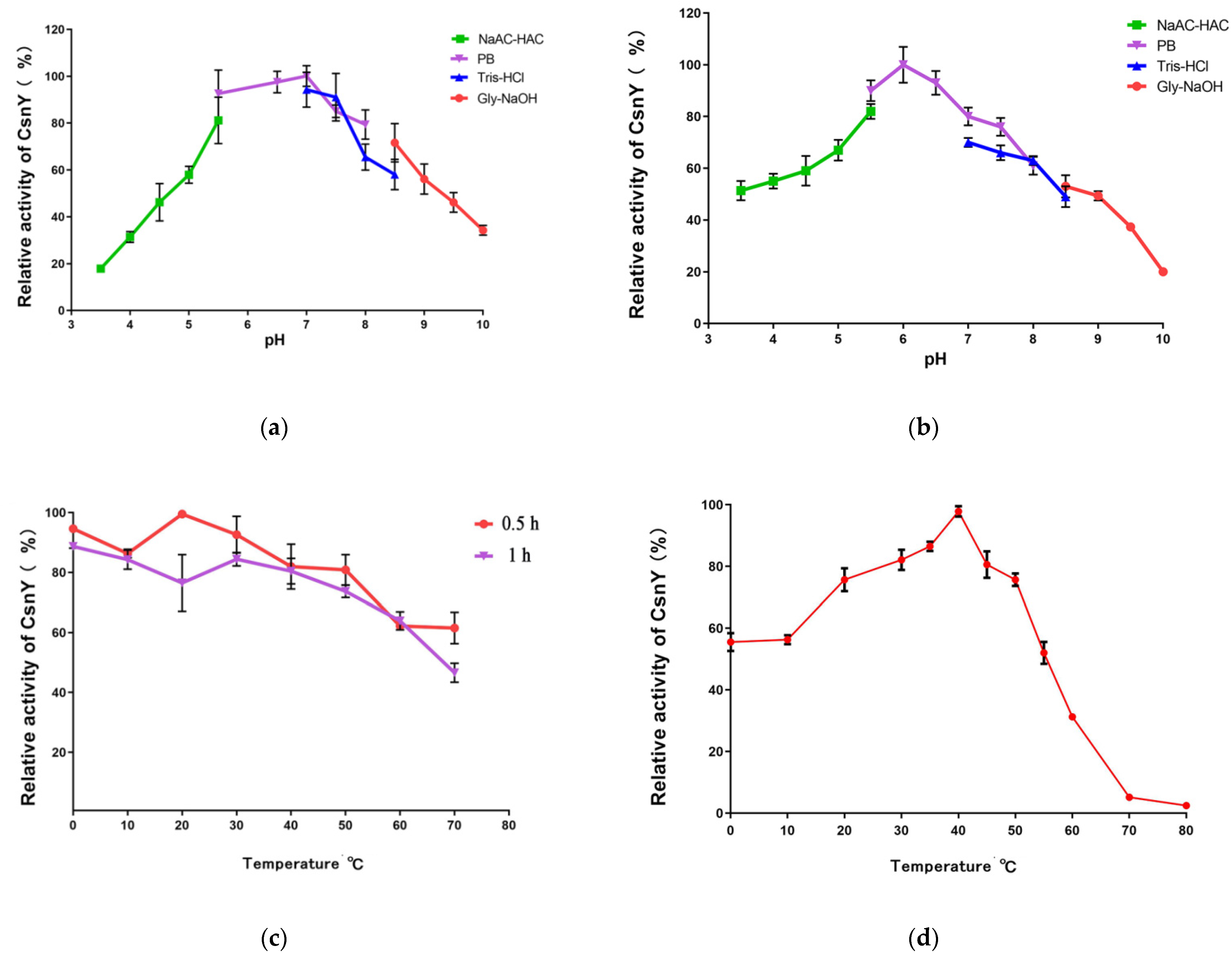
2.3. Effects of Temperature and pH on CsnY Activity and Stability
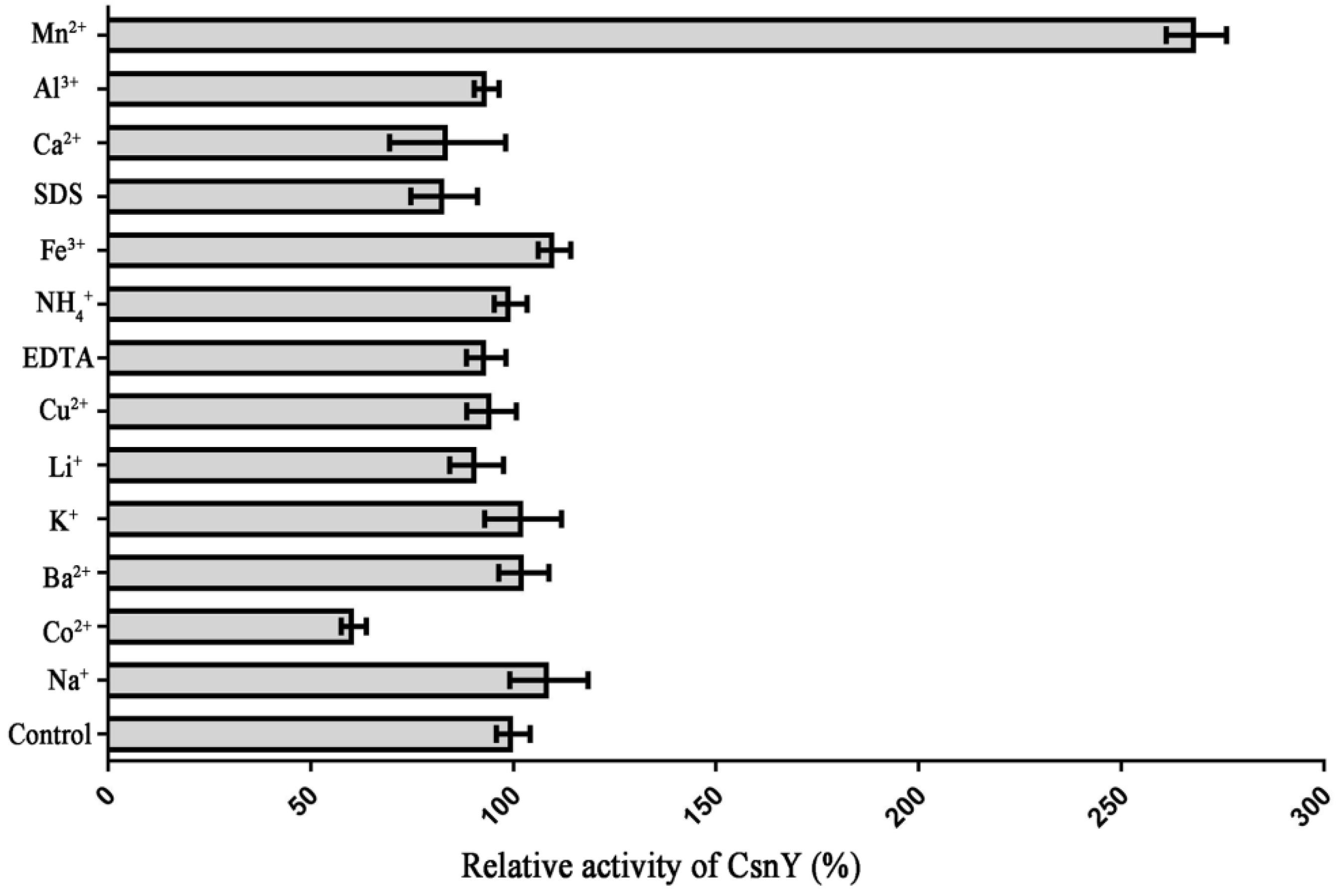
2.4. The Effects of Various Metal Ions or Chemicals on CsnY
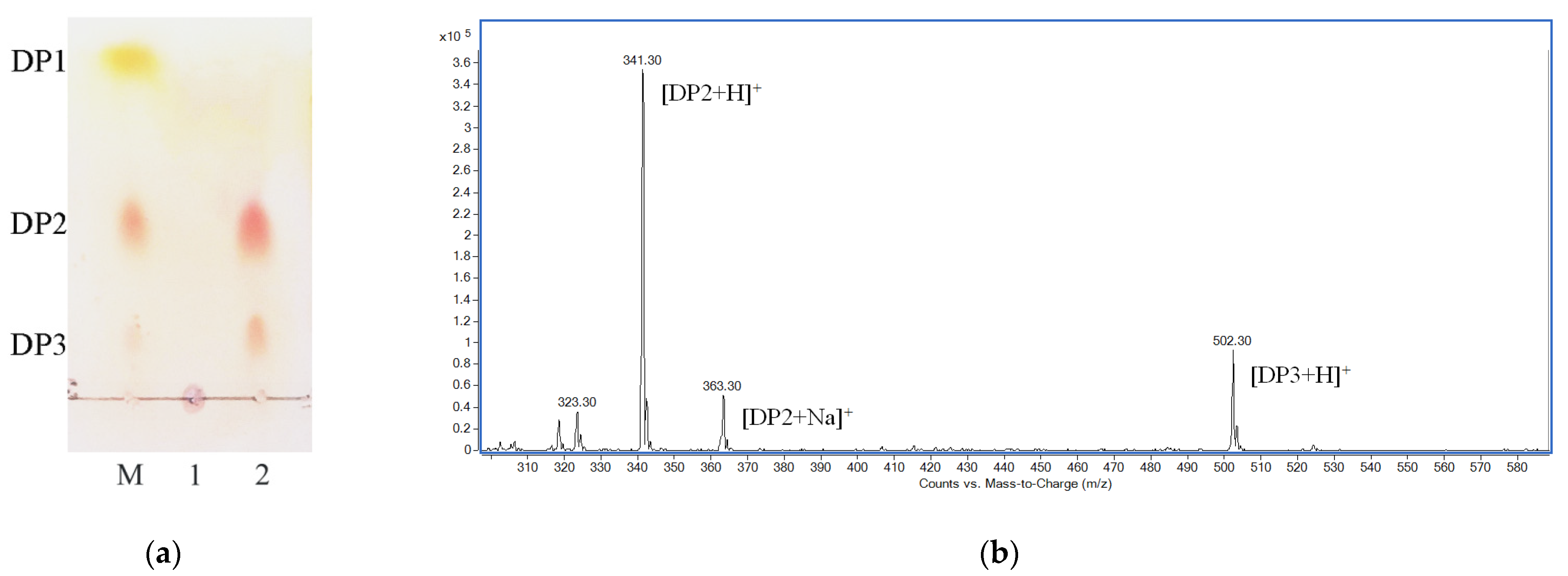
2.5. Final Degradation Product Analysis
3. Discussion
4. Materials and Methods
4.1. Materials, Strains, Plasmids, and Media
4.2. Sequence Analysis of CsnY
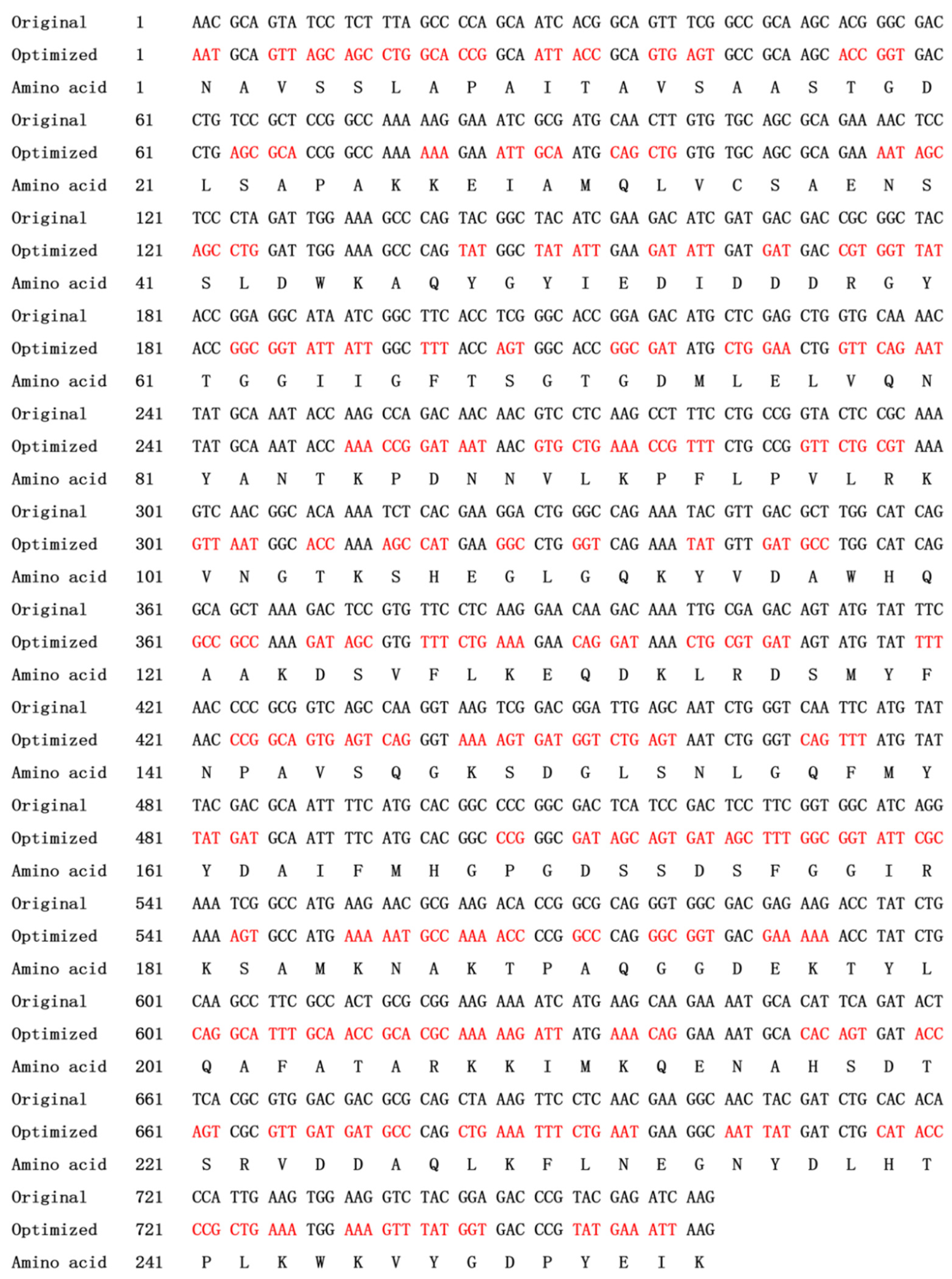
4.3. Codon Optimization and Construction of Expression Vector
4.4. Expression and Purification of CsnY
4.5. Measurement of CsnY Activity
4.6. Effects of Temperature and pH on CsnY Activity and Stability
4.7. The Effects of Various Metal Ions or Chemicals on CsnY Activity
4.8. Analysis of Degradation Products
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Production of a thermostable chitosanase from shrimp heads via Paenibacillus Mucilaginosus TKU032 conversion and its application in the preparation of bioactive chitosan oligosaccharides. Mar. Drugs 2019, 17, 217. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.; Mengibar, M.; Fernandez, M.; Alemany, S.; Heras, A.; Acosta, N. Influence of preparation methods of chitooligosaccharides on their physicochemical properties and their anti-inflammatory effects in mice and in RAW 264.7 macrophages. Mar. Drugs 2018, 16, 430. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.X.; Chen, H.X.; Zhang, J.; Zhang, X.D.; Liang, Y.X. Protective effect of chitooligosaccharides against Cyclophosphamide-induced immunosuppression in mice. Int. J. Biol. Macromol. 2013, 62, 330–335. [Google Scholar] [CrossRef]
- Lin, S.M.; Mao, S.H.; Guan, Y.; Lin, X.; Luo, L. Dietary administration of chitooligosaccharides to enhance growth, innate immune response and disease resistance of Trachinotus ovatus. Fish Shellfish Immun. 2012, 32, 909–913. [Google Scholar] [CrossRef]
- Inmaculada, A.; Niuris, A.; Concepción, C.; Begoña, E.; Javier, M.; Carolina, C.; María, G.; Angeles, C. Cosmetics and cosmeceutical applications of chitin, chitosan and their derivatives. Polymers 2018, 10, 213–238. [Google Scholar]
- Malerba, M.; Cerana, R. Recent advances of chitosan applications in plants. Polymers 2018, 10, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, C.; Auza, L.G.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of chitin to defined chitosan oligomers: Current status and future prospects. Mar. Drugs 2019, 17, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, W.J.; Park, R.D. Bioproduction of chitooligosaccharides: Present and perspectives. Mar. Drugs 2014, 12, 5328–5356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.K.; Je, J.Y.; Lee, S.J.; Kim, Y.S.; Hwang, J.W.; Sung, S.H.; Moon, S.H.; Jeon, B.T.; Kim, S.K.; Jeon, Y.J.; et al. Chitooligosaccharides induce apoptosis in human myeloid leukemia Hl-60 cells. Bioorg. Med. Chem. Lett. 2012, 22, 6136–6138. [Google Scholar] [CrossRef] [PubMed]
- Viens, P.; Lacombe-Harvey, M.E.; Brzezinski, R. Chitosanases from family 46 of glycoside hydrolases: From proteins to phenotypes. Mar. Drugs 2015, 13, 6566–6587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kim, S.K. Application of marine microbial enzymes in the food and pharmaceutical industries. Adv. Food. Nutr. Res. 2012, 65, 423–435. [Google Scholar] [PubMed]
- Xing, P.C.; Liu, D.; Yu, W.G.; Lu, X.Z. Molecular characterization of an endo-type chitosanase from the fish pathogen Renibacterium sp. QD1. J. Mar. Biol. Assoc. UK 2014, 94, 681–686. [Google Scholar] [CrossRef]
- Guan, F.; Han, Y.; Yan, K.; Zhang, Y.; Zhang, Z.; Wu, N.; Tian, J. Highly efficient production of chitooligosaccharides by enzymes mined directly from the marine metagenome. Carbohydr. Polym. 2020, 234, 115909. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Mohd Omar, S.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from Psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zheng, Z.; Xiao, Y.; Zhang, J.; Zhou, Y.; Li, X.; Li, S.; Yu, H. Cloning and characterization of a cold-adapted chitosanase from marine bacterium Bacillus sp. BY01. Molecules 2019, 24, 3915. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, X.H.; Li, X.; Han, Y.T.; Wang, Y.A.; Yao, R.Y.; Li, S.Y. Purification and characterization of a new cold-adapted and thermo-tolerant chitosanase from marine bacterium Pseudoalteromonas sp. SY39. Molecules 2019, 24, 183. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, K.S.; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends. Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Åqvist, J.; Isaksen, G.V.; Brandsdal, B.O. Computation of enzyme cold adaptation. Nat. Rev. Chem. 2017, 1, 0051. [Google Scholar] [CrossRef]
- Shinya, S.; Fukamizo, T. Interaction between chitosan and its related enzymes: A review. Int. J. Biol. Macromol. 2017, 104, 1422–1435. [Google Scholar] [CrossRef]
- Fukamizo, T.; Juffer, A.H.; Vogel, H.J.; Honda, Y.; Tremblay, H.; Boucher, I.; Neugebauer, W.A.; Brzezinski, R. Theoretical calculation of pKa reveals an important role of Arg205 in the activity and stability of Streptomyces sp. N174 chitosanase. J. Biol. Chem. 2000, 275, 25633–25640. [Google Scholar] [CrossRef] [Green Version]
- Lacombe-Harvey, M.; Fortin, M.; Ohnuma, T.; Fukamizo, T.; Letzel, T.; Brzezinski, R. A highly conserved arginine residue of the chitosanase from Streptomyces sp. N174 is involved both in catalysis and substrate binding. BMC Biochem. 2013, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.; Sun, J.A.; Wang, W.; Gao, L.; Mao, X.Z. Cloning, expression and characterization of a novel chitosanase from Streptomyces albolongus ATCC 27414. Food. Chem. 2019, 286, 696–702. [Google Scholar] [CrossRef]
- Ma, C.R.; Li, X.; Yang, K.; Li, S.Y. Characterization of a new chitosanase from a marine Bacillus sp. and the anti-oxidant activity of its hydrolysate. Mar. Drugs 2020, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Bhuvanachandra, B.; Sivaramakrishna, D.; Alim, S.; Preethiba, G.; Rambabu, S.; Swamy, M.J.; Podile, A.R. New class of chitosanase from Bacillus amyloliquefaciens for the generation of chitooligosaccharides. J. Agric. Food. Chem. 2021, 69, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Qin, Z.; Chen, Q.M.; Fan, L.Q.; Jiang, L.H.; Zhao, L.M. High level production of a Bacillus amlyoliquefaciens chitosanase in Pichia Pastoris suitable for chitooligosaccharides preparation. Int. J. Biol. Macromol. 2020, 149, 1034–1041. [Google Scholar] [CrossRef]
- Sun, H.H.; Cao, R.; Li, L.H.; Zhao, L.; Qiu, L. Cloning, purification and characterization of a novel GH46 family chitosanase, Csn-CAP, from Staphylococcus capitis. Process. Biochem. 2018, 75, 146–151. [Google Scholar] [CrossRef]
- Wang, Y.N.; Qin, Z.; Fan, L.Q.; Zhao, L.M. Structure–function analysis of Gynuella sunshinyii chitosanase uncovers the mechanism of substrate binding in GH family 46 members. Int. J. Biol. Macromol. 2020, 165, 2038–2048. [Google Scholar] [CrossRef]
- Zheng, Q.; Meng, X.; Cheng, M.; Li, Y.; Liu, Y.; Chen, X. Cloning and Characterization of a New Chitosanase From a Deep-Sea Bacterium Serratia sp. QD07. Front. Microbiol. 2021, 12, 360. [Google Scholar] [CrossRef]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell. Fact. 2010, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, M.B.; Struszczyk-Swita, K.; Li, X.; Szczesna-Antczak, M.; Daroch, M. Enzymatic modifications of chitin, chitosan, and chitooligosaccharides. Front. Bioeng. Biotechnol. 2019, 7, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Cao, H.; Li, S.; Zhao, Y.; Wang, W.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new family 75 chitosanase from Aspergillus sp. W-2. Int. J. Biol. Macromol. 2015, 81, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Tan, H.Q.; Zhu, C.; Liao, L.; Zhang, X.Q.; Wu, M. Cloning and overexpression of a new chitosanase gene from Penicillium sp. D-1. AMB Express 2012, 2, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goo, B.G.; Park, J.K. Characterization of an alkalophilic extracellular chitosanase from Bacillus cereus GU-02. J. Biosci. Bioeng. 2014, 117, 684–689. [Google Scholar] [CrossRef]
- Gao, X.A.; Jung, W.J.; Kuk, J.H.; Park, R.D. Reaction pattern of Bacillus cereus D-11 chitosanase on chitooligosaccharide alcohols. J. Microbiol. Biotechnol. 2009, 19, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Li, J.; Wang, Y.L.; Liu, S.; Wang, Z.P.; Yu, X.J. Integrated approaches to reveal genes crucial for tannin degradation in Aureobasidium melanogenum T9. Biomolecules 2019, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.L.; Wu, Y.N.; Shi, K.; Xiao, X.F.; Gao, Y.; Li, L.; Qiao, Y.L. Study on the degradation performance and kinetics of immobilized cells in straw-alginate beads in marine environment. Bioresour. Technol. 2019, 280, 88–94. [Google Scholar] [CrossRef]
- Zhou, H.X.; Xu, S.S.; Yin, X.J.; Wang, F.L.; Li, Y. Characterization of a new bifunctional and cold-adapted polysaccharide lyase (PL) family 7 alginate lyase from Flavobacterium sp. Mar. Drugs 2020, 18, 388. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Shao, Y.; Jiao, C.; Yang, Q.M.; Weng, H.F.; Xiao, A.F. Characterization and application of an alginate lyase, Aly1281 from marine bacterium Pseudoalteromonas carrageenovora ASY5. Mar. Drugs 2020, 18, 95. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name/Source | Mw (kDa) | Optimum pH/Temperature (°C) | Specific Activity (U/mg) | Relative Activity at 10/20/30 °C | Thermal Stability | Final Products | Reference |
|---|---|---|---|---|---|---|---|
| CsnY/Renibacterium sp. Y82 | 27.8 | 6.0/40 | 330.67 | 60%/80%/>80% | 80% activity remained after 1 h at 50 °C | DP2,3 | This study |
| CsnB/Bacillus sp. BY01 | 30.89 | 5.0/35 | 329.3 | 40.4%/76.8%/>80% | 10% activity retained after 1 h at 40 °C | DP2,3 | [15] |
| CsnM/Pseudoalteromonas sp. SY39 | 28 | 5.9/40 | 393.2 | 30.6%/>50%/80% | only 25.4% and 15.8% activity retained at 30 and 40 °C for 1 h | DP2,3 | [16] |
| Csn-CAP/Staphylococcus capitis | 35 | 7.0/30 | 89.2 | N.D./90%/90% | 50% activity retained at 55 °C for 1 h | DP2,3 | [27] |
| GsCsn46A/Gynuella sunshinyii | 29.7 | 5.5/30 | 260.39 | 70%/>80%/>80% | 80% activity remained at 30 °C for 1 h | DP2,3 | [28] |
| CsnS/Serratia sp. QD07 | 27.1 | 5.8/60 | 412.6 | 42.6%/40%/>40% | 80% activity retained at 30 °C for 2 h | DP2,3 | [29] |
| N.D./Janthinobacterium sp. 4239 | 29 | 5.0/45 | 1500 | 30%/60%/70% | almost all the activity retained after 30 min at 50 °C | DP1–3 | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.-L.; Jiang, X.-H.; Xiao, X.-F.; Zhang, W.-X.; Shi, Y.-Q.; Wang, Z.-P.; Zhou, H.-X. Expression and Characterization of a Novel Cold-Adapted Chitosanase from Marine Renibacterium sp. Suitable for Chitooligosaccharides Preparation. Mar. Drugs 2021, 19, 596. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110596
Zhang L-L, Jiang X-H, Xiao X-F, Zhang W-X, Shi Y-Q, Wang Z-P, Zhou H-X. Expression and Characterization of a Novel Cold-Adapted Chitosanase from Marine Renibacterium sp. Suitable for Chitooligosaccharides Preparation. Marine Drugs. 2021; 19(11):596. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110596
Chicago/Turabian StyleZhang, Lin-Lin, Xiao-Hua Jiang, Xin-Feng Xiao, Wen-Xiu Zhang, Yi-Qian Shi, Zhi-Peng Wang, and Hai-Xiang Zhou. 2021. "Expression and Characterization of a Novel Cold-Adapted Chitosanase from Marine Renibacterium sp. Suitable for Chitooligosaccharides Preparation" Marine Drugs 19, no. 11: 596. https://0-doi-org.brum.beds.ac.uk/10.3390/md19110596