
The Common Sunstar Crossaster papposus—A Neurotoxic Starfish

 ,
,
Abstract
:1. Introduction
2. Results
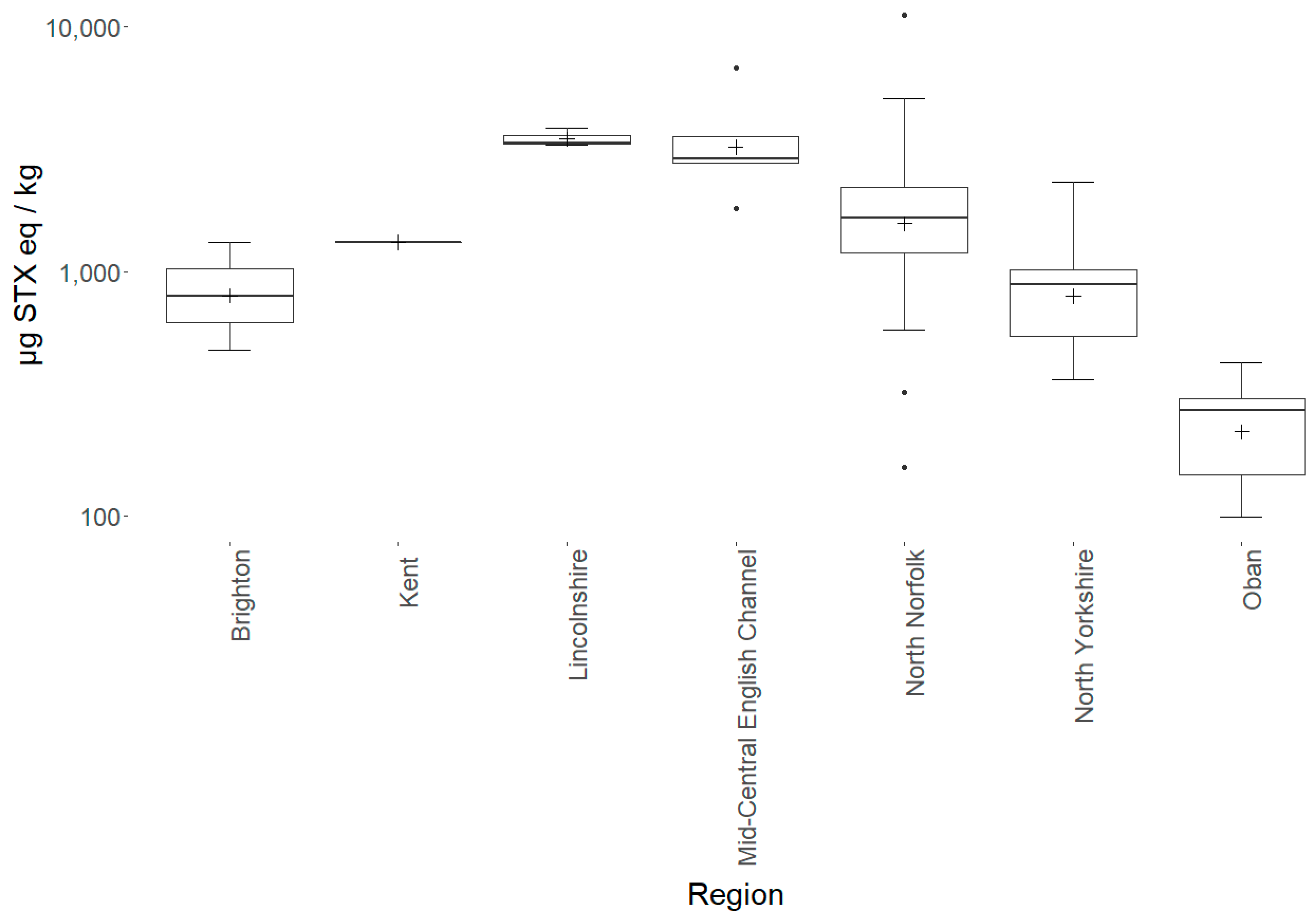
2.1. Starfish Toxicity
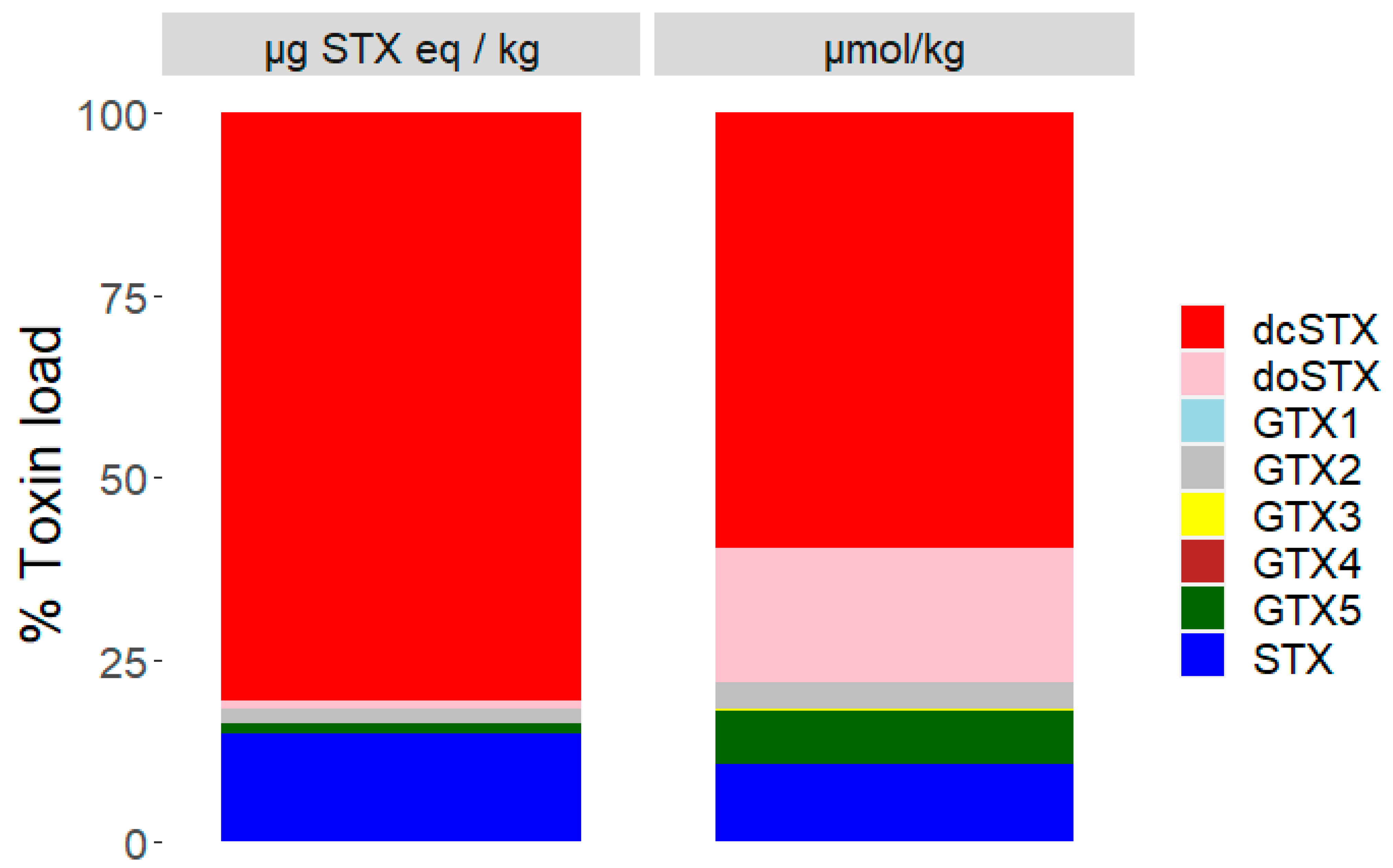
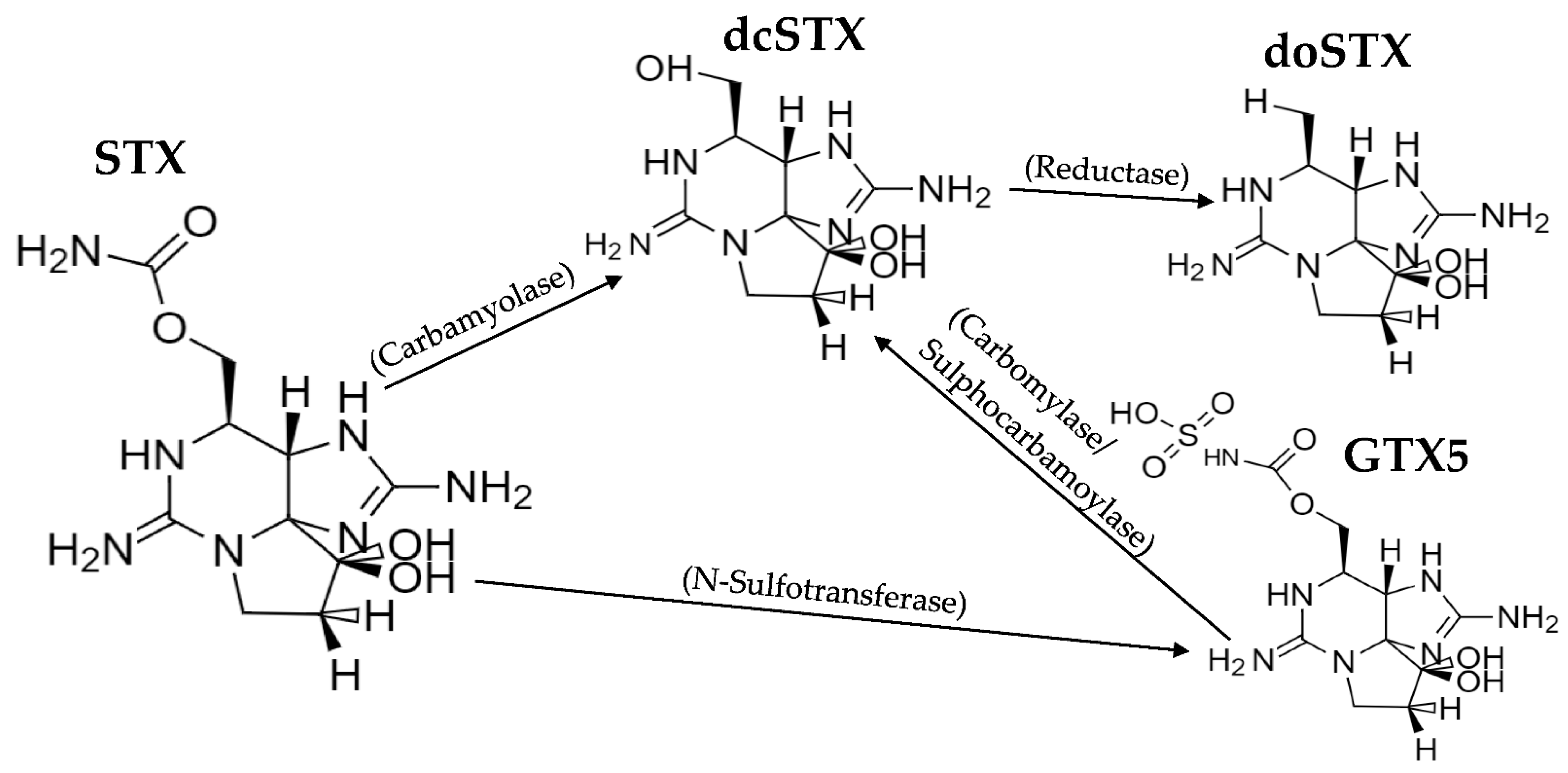
2.2. Toxin Profiles
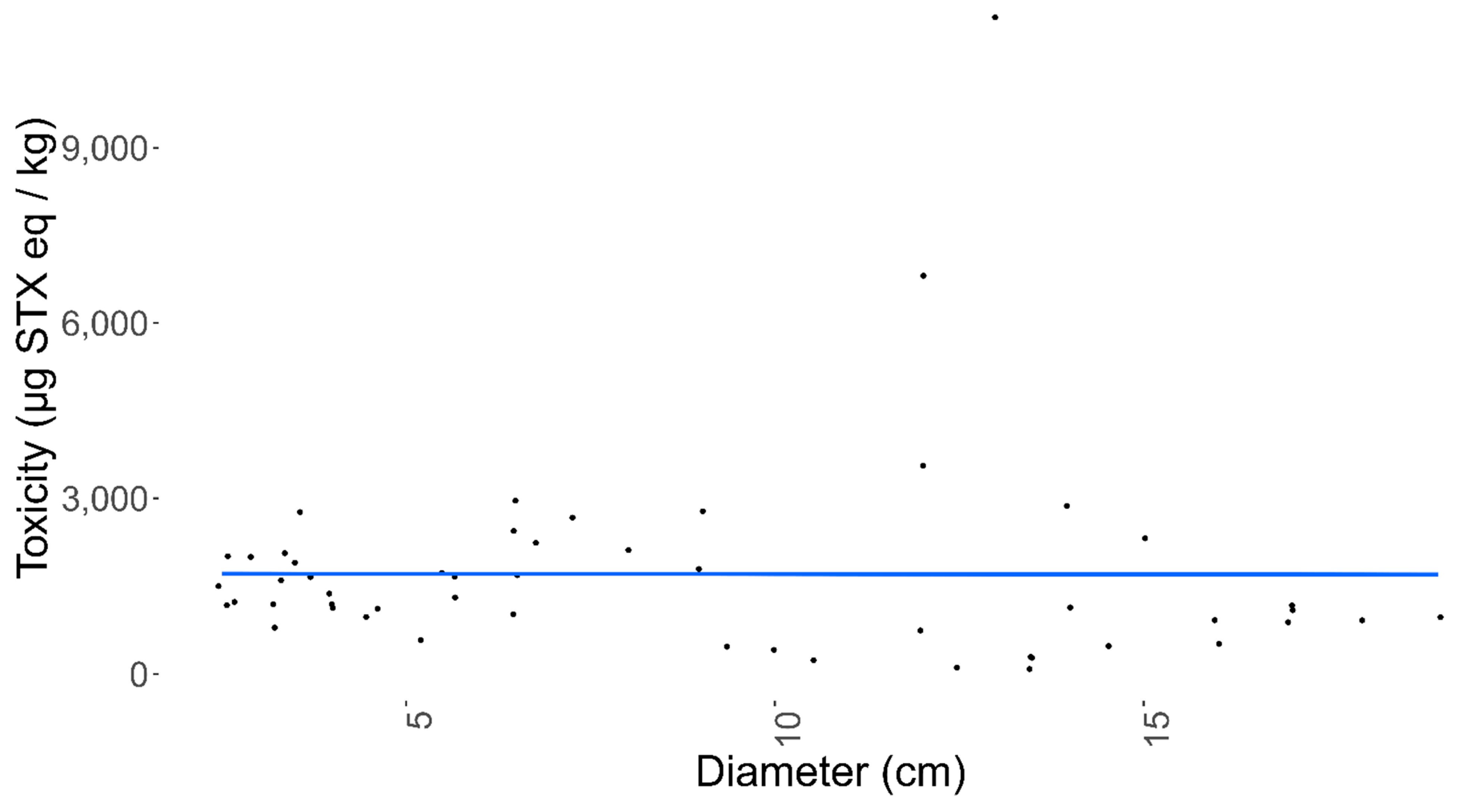
2.3. Sunstar Physiological Analysis
2.4. Comparison of Detection Techniques
3. Discussion
3.1. Starfish Toxicity
3.2. Toxin Profiles
3.3. Sunstar Physiological Analysis
4. Materials and Methods
4.1. Sample Collection
4.2. Reagents and Chemicals
4.3. Sample Preparation and Extraction for Toxin Analysis
4.4. Sample Analysis
4.4.1. Analysis of STXs by LC–FLD
4.4.2. Analysis of STXs by HILIC–MS/MS
4.5. LC–HRMS Qualitative Analysis of doSTX
4.6. Histological Processing and Analysis of Sunstars
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
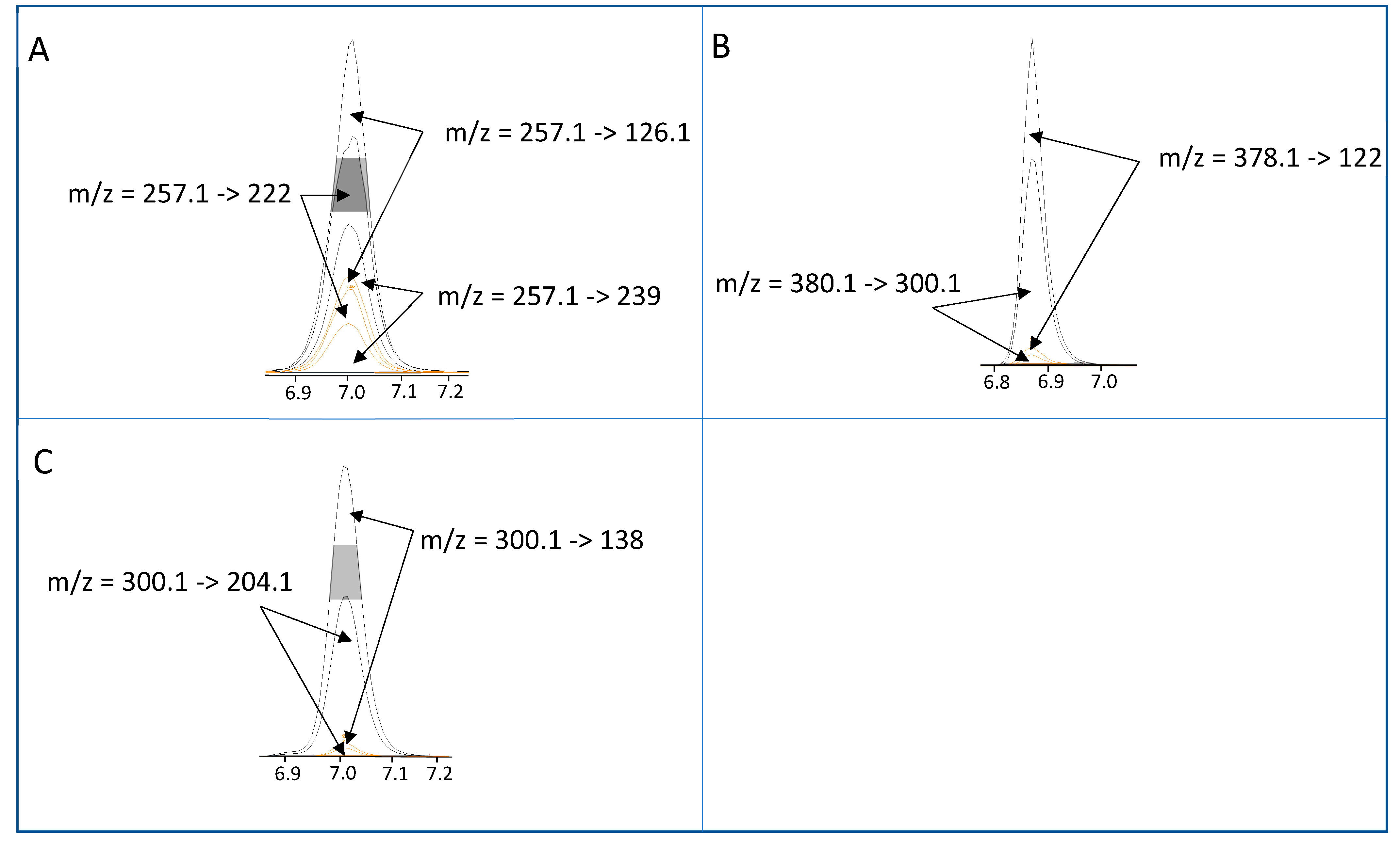{kind=link}
{kind=link}
{kind=link}
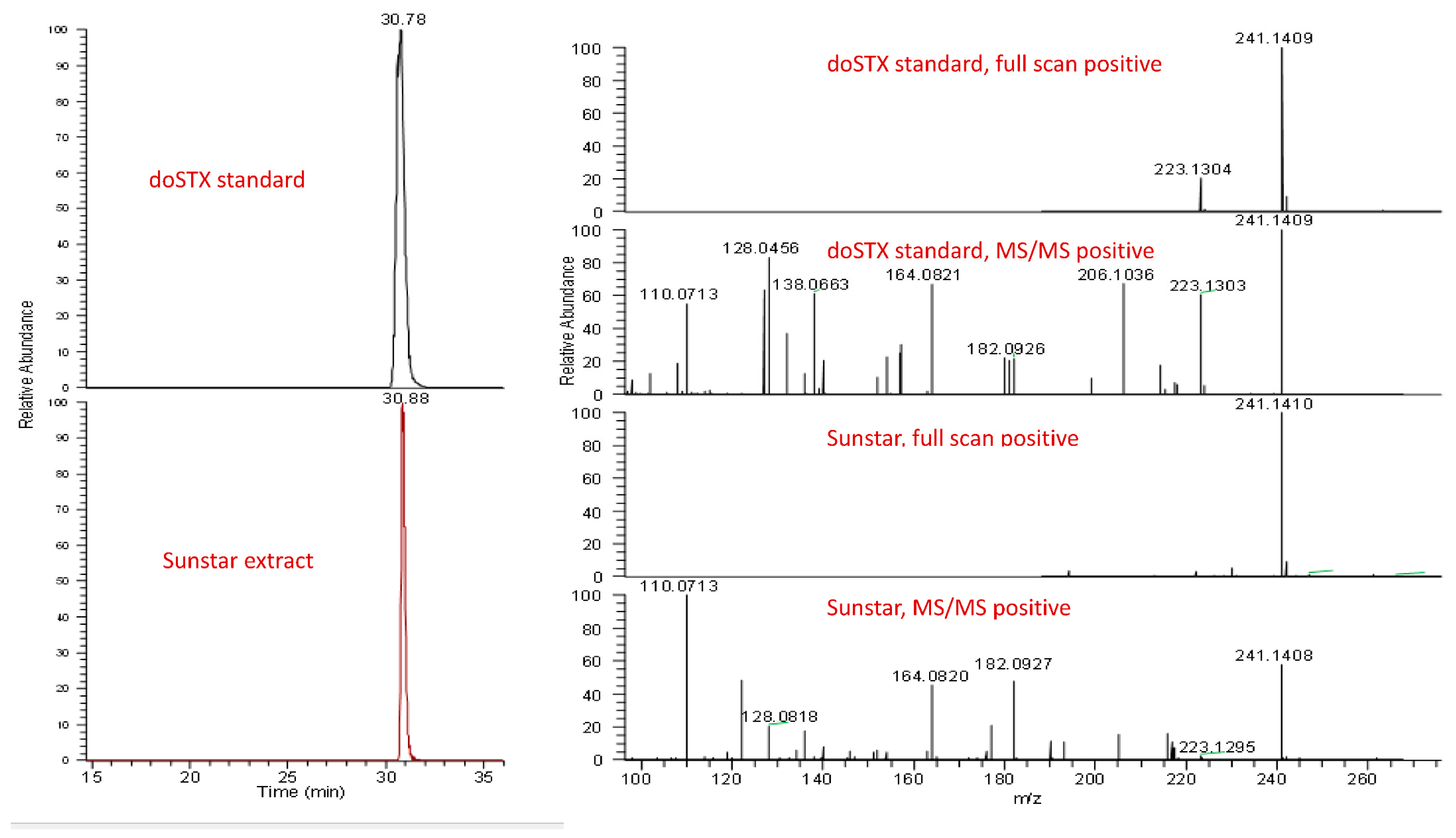{kind=link}
| Common Name | Scientific Name | Date Sampled | Location | Region | Diameter (cm) | HILIC–MS/MS Total (µg STX eq/kg) | LC–FLD Total (µg STX eq/kg) |
|---|---|---|---|---|---|---|---|
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | nt |
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | nt |
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | nt |
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | 1 |
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | 4 |
| Common starfish | Asterias rubens | 6 February 2018 | Felixstowe | Suffolk | nt | nd | nt |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | 4 |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | 1 |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 8 February 2018 | Lowestoft | East Suffolk | nt | nd | nd |
| Sunstar | Crossaster papposus | 16 February 2018 | Holkham Beach | North Norfolk | nt | 157 | 388 |
| Common starfish | Asterias rubens | 19 February 2018 | Lulworth Banks | Dorset | nt | nd | nd |
| Common starfish | Asterias rubens | 19 February 2018 | Lulworth Banks | Dorset | nt | nd | nd |
| Common starfish | Asterias rubens | 20 February 2018 | Aldeburgh Beach | East Suffolk | nt | nd | 2 |
| Common starfish | Asterias rubens | 20 February 2018 | Aldeburgh Beach | East Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 20 February 2018 | Aldeburgh Beach | East Suffolk | nt | nd | 167 |
| Common starfish | Asterias rubens | 3 March 2018 | Hunstanton Beach | North Norfolk | nt | nd | 10 |
| Common starfish | Asterias rubens | 3 March 2018 | Hunstanton Beach | North Norfolk | nt | nd | 5 |
| Common starfish | Asterias rubens | 3 March 2018 | Hunstanton Beach | North Norfolk | nt | nd | 10 |
| Common starfish | Asterias rubens | 5 March 2018 | Aldeburgh Beach | East Suffolk | nt | nd | 5 |
| Common starfish | Asterias rubens | 5 March 2018 | Aldeburgh Beach | East Suffolk | nt | nd | 3 |
| Common starfish | Asterias rubens | 5 March 2018 | Aldeburgh Beach | East Suffolk | nt | nd | 7 |
| Sunstar | Crossaster papposus | 5 March 2018 | Brancaster | North Norfolk | nt | 321 | 781 |
| Sunstar | Crossaster papposus | 5 March 2018 | Brancaster | North Norfolk | nt | 579 | 1357 |
| Sunstar | Crossaster papposus | 5 March 2018 | Wells | North Norfolk | nt | 1640 | 4980 |
| Sunstar | Crossaster papposus | 5 March 2018 | Hunstanton Beach | North Norfolk | nt | 2727 | 6679 |
| Sunstar | Crossaster papposus | 5 March 2018 | Hunstanton Beach | North Norfolk | nt | 2991 | 4778 |
| Sunstar | Crossaster papposus | 5 March 2018 | Wells | North Norfolk | nt | 3543 | 8489 |
| Sunstar | Crossaster papposus | 5 March 2018 | Wells | North Norfolk | nt | 5108 | 16,513 |
| Sunstar | Crossaster papposus | 5 March 2018 | Brancaster | North Norfolk | nt | nd | 13,237 |
| Common starfish | Asterias rubens | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | nd | 4 |
| Common starfish | Asterias rubens | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | nd | 4 |
| Common starfish | Asterias rubens | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | nd | 1 |
| Sunstar | Crossaster papposus | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | 3847 | 9058 |
| Sunstar | Crossaster papposus | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | 3292 | 10,993 |
| Sunstar | Crossaster papposus | 6 March 2018 | Lincolnshire coast | Lincolnshire | nt | 3343 | 5746 |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | 2 | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | nd | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | 3 | nt |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | 4 | nt |
| Sunstar | Crossaster papposus | 6 March 2018 | Ramsgate beach | Kent | nt | 1317 | 7061 |
| Common starfish | Asterias rubens | 6 March 2018 | Ramsgate beach | Kent | nt | 16 | 4 |
| Common starfish | Asterias rubens | 12 March 2018 | Felixstowe | Suffolk | nt | nd | nd |
| Common starfish | Asterias rubens | 12 March 2018 | Felixstowe | Suffolk | nt | nd | 7 |
| Common starfish | Asterias rubens | 12 March 2018 | Felixstowe | Suffolk | nt | nd | 5 |
| Spiny starfish | Marthasterias glacialis | 19 September 2019 | Cornwall—south of St.Austell | South Cornwall | nt | nd | nd |
| Common starfish | Asterias rubens | 14 October 2019 | Brixham | South Devon | nt | nd | nd |
| Common starfish | Asterias rubens | 24 October 2019 | South of Lyme Regis | Dorset | nt | nd | nd |
| Brittlestar | Ophiura ophiura | 25 October 2019 | South of Lyme Regis | Dorset | nt | nd | nd |
| Common starfish | Asterias rubens | 14 November 2019 | Brighton | Brighton | nt | nd | nd |
| Common starfish | Asterias rubens | 6 December 2019 | Plymouth | South Devon | nt | nd | nd |
| Common starfish | Asterias rubens | 17 December 2019 | Plymouth | South Devon | nt | nd | nd |
| Spiny starfish | Marthasterias glacialis | 17 December 2019 | Plymouth | South Devon | nt | 4 | 22 |
| Common starfish | Asterias rubens | 17 December 2019 | East of Whitby | North Yorkshire | nt | 89 | 48 |
| Common starfish | Asterias rubens | 17 December 2019 | East of Whitby | North Yorkshire | nt | 8 | 14 |
| Spiny starfish | Marthasterias glacialis | 17 December 2019 | East of Whitby | North Yorkshire | nt | 4 | 8 |
| Sunstar | Crossaster papposus | 17 December 2019 | East of Whitby | North Yorkshire | nt | 360 | 686 |
| Sunstar | Crossaster papposus | 17 December 2019 | East of Whitby | North Yorkshire | nt | 564 | 909 |
| Sunstar | Crossaster papposus | 17 December 2019 | East of Whitby | North Yorkshire | nt | 880 | 1447 |
| Sunstar | Crossaster papposus | 17 December 2019 | East of Whitby | North Yorkshire | nt | 548 | 779 |
| Sunstar | Crossaster papposus | 17 December 2019 | East of Whitby | North Yorkshire | nt | 472 | 625 |
| Sunstar | Crossaster papposus | 23 January 2020 | South of Brighton | Brighton | 9.4 | 479 | 795 |
| Sunstar | Crossaster papposus | 23 January 2020 | South of Brighton | Brighton | 5.7 | 1319 | 1929 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.5 | nt | 3109 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 5.2 | 590 | 837 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.2 | 805 | 1429 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 4.5 | 986 | 1420 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 6.5 | 1034 | 1667 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 4.6 | 1130 | 1609 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 4 | 1142 | 2115 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 2.6 | 1188 | 1803 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.2 | 1201 | 1610 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 4 | 1203 | 1548 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 2.7 | 1242 | 2146 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 4 | 1386 | 1726 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 2.5 | 1513 | 2632 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.3 | 1612 | 2864 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.7 | 1669 | 2460 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 5.7 | 1672 | 2651 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 6.5 | 1701 | 2545 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 5.5 | 1738 | 2633 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.5 | 1911 | 2682 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 2.9 | 2011 | 3500 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 2.6 | 2022 | 3131 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.4 | 2075 | 4104 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 8 | 2128 | 3982 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 6.8 | 2251 | 3825 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 6.5 | 2455 | 3910 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 7.3 | 2682 | 3648 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 3.6 | 2776 | 4131 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 6.5 | 2973 | 4745 |
| Sunstar | Crossaster papposus | 27 January 2020 | North Norfolk, near the Wash | North Norfolk | 13 | 11,245 | 16,244 |
| Common starfish | Asterias rubens | 29 January 2020 | Kings Lynn | North Norfolk | nt | nd | 7 |
| Goosefoot starfish | Anseropoda placenta | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | nt | nd | 23 |
| Common starfish | Asterias rubens | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | nt | nd | 8 |
| Brittlestar | Ophiura ophiura | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | nt | 121 | 112 |
| Sunstar | Crossaster papposus | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | 9 | 2790 | 4230 |
| Sunstar | Crossaster papposus | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | 12 | 6821 | 9510 |
| Sunstar | Crossaster papposus | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | 9 | 1804 | 3272 |
| Sunstar | Crossaster papposus | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | 12 | 3570 | 5500 |
| Sunstar | Crossaster papposus | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | 14 | 2881 | 8480 |
| Sand star | Astropecten irregularis | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | nt | nd | 36 |
| Seven-armed starfish | Luidia ciliaris | 6 February 2020 | Mid-central English Channel | Mid-central English Channel | nt | nd | nd |
| Common starfish | Asterias rubens | 3 March 2020 | South of Bridport | Dorset | nt | nd | nd |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 16 | 526 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 14 | 1148 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 19 | 984 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 17 | 896 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 16 | 933 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 14.5 | 492 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 17 | 1184 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 17 | 1104 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 18 | 930 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 15 | 2327 | nt |
| Sunstar | Crossaster papposus | 20 March 2020 | East of Whitby | North Yorkshire | 12 | 756 | nt |
| Sunstar | Crossaster papposus | 3 May 2020 | Hunstanton Beach | North Norfolk | nt | 1380 | 940 |
| Sunstar | Crossaster papposus | 3 May 2020 | Hunstanton Beach | North Norfolk | nt | 1607 | 1231 |
| Common starfish | Asterias rubens | 3 May 2020 | Cley shingle beach (from shoreline) | North Norfolk | nt | nd | nd |
| Goosefoot starfish | Anseropoda placenta | 21 July 2020 | Weymouth | Dorset | nt | 30 | nd |
| Sand star | Astropecten irregularis | 28 August 2020 | South of Dartmouth | South Devon | nt | 3 | nt |
| Sand star | Astropecten irregularis | 28 August 2020 | South of Dartmouth | South Devon | nt | 13 | nt |
| Sand star | Astropecten irregularis | 28 August 2020 | South of Dartmouth | South Devon | nt | 2 | nt |
| Common starfish | Asterias rubens | 28 August 2020 | South of Dartmouth | South Devon | nt | nd | nt |
| Goosefoot starfish | Anseropoda placenta | 28 August 2020 | South of Dartmouth | South Devon | nt | 9 | nt |
| Bloody Henry | Henrica oculata | 28 August 2020 | South of Dartmouth | South Devon | nt | 6 | nt |
| Brittlestar | Ophiura ophiura | 28 August 2020 | South of Dartmouth | South Devon | nt | 2 | nt |
| Brittlestar | Ophiura ophiura | 28 August 2020 | South of Dartmouth | South Devon | nt | 164 | nt |
| Brittlestar | Ophiura ophiura | 28 August 2020 | South of Dartmouth | South Devon | nt | nd | nt |
| Seven-armed starfish | Luidia ciliaris | 28 August 2020 | South of Dartmouth | South Devon | nt | 12 | nt |
| Goosefoot starfish | Anseropoda placenta | 28 August 2020 | South of Dartmouth | South Devon | nt | 4 | nt |
| Common starfish | Asterias rubens | 28 August 2020 | South of Dartmouth | South Devon | nt | 63 | nt |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 10.5 | 250 | 270 |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 13.5 | 99 | 125 |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 12.5 | 124 | 134 |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 10 | 425 | 452 |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 13.5 | 291 | 419 |
| Sunstar | Crossaster papposus | 11 March 2021 | Oban | Oban | 13.5 | 307 | 320 |
| Common Name | Scientific Name | Mean Toxicity | sd | Range | n | % > LOD |
|---|---|---|---|---|---|---|
| Bloody Henry | Henricia spp. | 6 | NA | NA | 1 | 100% |
| Brittlestar | Ophiura ophiura | 96 | 84 | nd–164 | 5 | 60% |
| Common starfish | Asterias rubens | 26 | 35 | nd–89 | 59 | 12% |
| Goosefoot starfish | Anserpoda placenta | 14 | 14 | nd–30 | 4 | 75% |
| Sand star | Astropecten irregularis | 6 | 6 | nd–13 | 4 | 75% |
| Seven-armed starfish | Luidia ciliaris | 12 | NA | nd–12 | 2 | 50% |
| Spiny starfish | Marthasterias glacialis | 4 | NA | nd–4 | 3 | 33% |
| Sunstar | Crossaster papposus | 1739 | 1669 | 99–11,245 | 71 | 100% |
| Region | Species | Mean Toxicity | sd | Range | n |
|---|---|---|---|---|---|
| Brighton | Common starfish | nd | N/A | N/A | 1 |
| Brighton | Sunstar | 899 | 594 | 479-1319 | 2 |
| Dorset | Brittlestar | nd | N/A | N/A | 1 |
| Dorset | Common starfish | nd | N/A | N/A | 3 |
| Dorset | Goosefoot starfish | 30 | N/A | N/A | 1 |
| East Suffolk | Common starfish | nd | N/A | N/A | 16 |
| Mid-central English Channel | Seven-armed starfish | nd | N/A | N/A | 1 |
| Mid-central English Channel | Brittlestar | 121 | N/A | N/A | 1 |
| Mid-central English Channel | Common starfish | nd | N/A | N/A | 1 |
| Mid-central English Channel | Goosefoot starfish | nd | N/A | N/A | 1 |
| Mid-central English Channel | Sand star | nd | N/A | N/A | 1 |
| Mid-central English Channel | Sunstar | 3573 | 1922 | 1804–6821 | 5 |
| Kent | Common starfish | 2 | 4 | nd-16 | 13 |
| Kent | Sunstar | 1317 | N/A | N/A | 1 |
| Lincolnshire | Common starfish | nd | N/A | N/A | 3 |
| Lincolnshire | Sunstar | 3494 | 307 | 3292–3847 | 3 |
| North Norfolk | Common starfish | nd | N/A | N/A | 5 |
| North Norfolk | Sunstar | 1910 | 1815 | 157–11,245 | 40 |
| North Yorkshire | Common starfish | 48 | 57 | 8–89 | 2 |
| North Yorkshire | Spiny starfish | 4 | N/A | N/A | 1 |
| North Yorkshire | Sunstar | 882 | 466 | 360–2326 | 16 |
| Oban | Sunstar | 249 | 122 | 99–425 | 6 |
| South Cornwall | Spiny starfish | nd | N/A | N/A | 1 |
| South Devon | Seven-armed starfish | 12 | N/A | N/A | 1 |
| South Devon | Bloody Henry | 6 | N/A | N/A | 1 |
| South Devon | Brittlestar | 55 | 94 | nd–164 | 3 |
| South Devon | Common starfish | 13 | 28 | nd–63 | 5 |
| South Devon | Goosefoot starfish | 6 | 4 | 4–9 | 2 |
| South Devon | Sand star | 6 | 6 | 2–12 | 3 |
| South Devon | Spiny starfish | 4 | N/A | N/A | 1 |
| Suffolk | Common starfish | nd | N/A | N/A | 9 |
| Organ | Sex | Mean Toxicity | sd | Range | n |
|---|---|---|---|---|---|
| Digestive gland | Female | 2871 | 2115 | 842–5666 | 7 |
| Digestive gland | Male | 4672 | 4232 | 700–10,339 | 6 |
| Gonad | Female | 14,234 | 16,952 | 669–45,766 | 7 |
| Gonad | Male | 2383 | 2167 | 234–5634 | 6 |
| Skin | Female | 1574 | 1035 | 128–3045 | 7 |
| Skin | Male | 3117 | 2685 | 766–7249 | 6 |


References
- Botana, L.M. Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, Chapter 2: Diversity of Marine and FreshwaterAlgal Toxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker Inc.: New York, NY, USA, 2000; pp. 20–24. ISBN 9780824789565. [Google Scholar]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Marine biotoxins in shellfish—Saxitoxin group. EFSA J. 2009, 7, 1019.
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. Harmful Algal Blooms: A Global Overview; UNESCO: Paris, France, 2004; ISBN 9231039482. [Google Scholar]
- Llewellyn, L.; Negri, A.; Robertson, A. Paralytic shellfish toxins in tropical oceans. Toxin Rev. 2006, 25, 159–196. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [Green Version]
- Anon Regulation (EC) No 854/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption. Off. J. Eur. Union 2004, L 139, 206–320.
- Anon Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. Union 2004, L 139, 55–205.
- Turner, A.D.; Rapkova, M.D.; Fong, S.Y.; Hungerford, J.; McNabb, P.S.; Boundy, M.J.; Harwood, D.T. Ultrahigh-Performance Hydrophilic Interaction Liquid Chromatography with Tandem Mass Spectrometry Method for the Determination of Paralytic Shellfish Toxins and Tetrodotoxin in Mussels, Oysters, Clams, Cockles, and Scallops: Collaborative Study. J. AOAC Int. 2020, 103, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Bresnan, E.; Graham, J.; Lacaze, J.; Turrell, E.; Collins, C.; Brown, L.; Bresnan, E.; Graham, J.; Lacaze, J.; et al. Distribution, diversity and toxin composition of the genus Alexandrium (Dinophyceae) in Scottish waters. Eur. J. Phycol. 2010, 45, 375–393. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, S.M.; Purdie, D.A.; Lilly, E.L.; Larsen, J.; Morris, S. Toxin profile, pigment composition, and large subunit rDNA phylogenetic analysis of an Alexandrium minutum (Dinophyceae) strain isolated from the fleet lagoon, United Kingdom. J. Phycol. 2005, 41, 343–353. [Google Scholar] [CrossRef]
- Lewis, A.M.; Coates, L.N.; Turner, A.D.; Percy, L.; Lewis, J. A review of the global distribution of Alexandrium minutum (Dinophyceae) and comments on ecology and associated paralytic shellfish toxin profiles, with a focus on northern Europe. J. Phycol. 2018, 54, 581–598. [Google Scholar] [CrossRef]
- Turner, A.D.; Stubbs, B.; Coates, L.; Dhanji-Rapkova, M.; Hatfield, R.G.; Lewis, A.M.; Rowland-Pilgrim, S.; O’Neil, A.; Stubbs, P.; Ross, S.; et al. Variability of paralytic shellfish toxin occurrence and profiles in bivalve molluscs from Great Britain from official control monitoring as determined by pre-column oxidation liquid chromatography and implications for applying immunochemical tests. Harmful Algae 2014, 31, 87–99. [Google Scholar] [CrossRef]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, Y.; Bolch, C.J.; Hallegraeff, G.M. Toxin composition of resting cysts of Alexandrium tamerense (Dinophycae). Toxicon 1992, 30, 1539–1544. [Google Scholar] [CrossRef]
- Persson, A.; Smith, B.C.; Wikfors, G.H.; Quilliam, M. Grazing on toxic Alexandrium fundyense resting cysts and vegetative cells by the eastern oyster (Crassostrea virginica). Harmful Algae 2006, 5, 678–684. [Google Scholar] [CrossRef]
- Hunt, O.D. The Food of the Bottom Fauna of the Plymouth Fishing Grounds. J. Mar. Biol. Assoc. UK 1925, 13, 560–599. [Google Scholar] [CrossRef] [Green Version]
- Quiblier, C.; Wood, S.; Echenique Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Baker, T.R.; Doucette, G.J.; Powell, C.L.; Boyer, G.L.; Plumley, F.G. GTX4imposters: Characterization of fluorescent compounds synthesized by Pseudomonas stutzeri SF/PS and Pseudomonas/Alteromonas PTB-1, symbionts of saxitoxin-producing Alexandrium spp. Toxicon 2003, 41, 339–347. [Google Scholar] [CrossRef]
- Gallacher, S.; Smith, E.A. Bacteria and paralytic shellfish toxins. Protist 1999, 150, 245–255. [Google Scholar] [CrossRef]
- Martins, C.A.; Alvito, P.; Tavares, M.J.; Pereira, P.; Doucette, G.; Franca, S. Reevaluation of production of paralytic shellfish toxin by bacteria associated with dinoflagellates of the Portuguese Coast. Appl. Environ. Microbiol. 2003, 69, 5693–5698. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Chai, T.; Hwang, D. Isolation of bacteria from toxic dinoflagellate Alexandrium minutum and their effects on algae toxicity. J. Nat. Toxins 2000, 9, 409–417. [Google Scholar]
- Bane, V.; Lehane, M.; Dikshit, M.; Riordan, A.O.; Furey, A. Tetrodotoxin: Chemistry, Toxicity, Source, Distribution and Detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, H.; Matsubara, S.; Miwa, N.; Akahane, S.; Murakami, M.; Goto, T.; Nara, M.; Noguchi, T.; Saito, T.; Shida, Y.; et al. Vibrio alginolyticus, a TTX-producing Bacterium Isolated from the Starfish Astropecten polyacanthus. Nippon Suisan Gakkaishi 1987, 53, 617–621. [Google Scholar] [CrossRef]
- Akbar, M.A.; Yusof, N.Y.M.; Tahir, N.I.; Ahmad, A.; Usup, G.; Sahrani, F.K.; Bunawan, H. Biosynthesis of saxitoxin in marine dinoflagellates: An omics perspective. Mar. Drugs 2020, 18, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The genetic basis of toxin biosynthesis in dinoflagellates. Microorganisms 2019, 7, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, moleculartargets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [Green Version]
- Raposo, M.I.C.; Gomes, M.T.S.R.; Jo, M.; Rudnitskaya, A. Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review. Toxins 2020, 12, 344. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.M.; Wang, W.X.; Lam, P.K.S.; Yu, P.K.N. The uptake, distribution and elimination of paralytic shellfish toxins in mussels and fish exposed to toxic dinoflagellates. Aquat. Toxicol. 2006, 80, 82–91. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: Occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Taleb, H.; Vale, P.; Jaime, E.; Blaghen, M. Study of paralytic shellfish poisoning toxin profile in shellfish from the Mediterranean shore of Morocco. Toxicon 2001, 39, 1855–1861. [Google Scholar] [CrossRef]
- Bazzoni, A.M.; Mudadu, A.G.; Lorenzoni, G.; Arras, I.; Lugliè, A.; Vivaldi, B.; Cicotelli, V.; Sanna, G.; Tedde, G.; Ledda, S.; et al. Occurrence of harmful algal species and shellfish toxicity in Sardinia (Italy). Ital. J. Food Saf. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, C.; Mardones, P.; Sfeir, A.; Lagos, N. Simultaneous presence of Paralytic and Diarrheic Shellfish Poisoning toxins in Mytilus chilensis samples collected in the Chiloe Island, Austral Chilean fjords. Biol. Res. 2004, 37, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; O’Connor, W.A.; Alvin, A.; Mihali, T.K.; Kalaitzis, J.; Neilan, B.A. Differential accumulation of paralytic shellfish toxins from Alexandrium minutum in the pearl oyster, Pinctada imbricata. Toxicon 2009, 54, 217–223. [Google Scholar] [CrossRef]
- Shumway, S.E. A Review of the Effects of Algal Blooms on Shellfish and Aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Wong, C.K.; Hung, P.; Ng, H.C.C.; Lee, S.Y.; Kam, K.M. Cluster analysis of toxins profile pattern as a tool for tracing shellfish contaminated with PSP-toxins. Environ. Res. 2011, 111, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Reyero, M.I.; Franco, J. Kinetics of accumulation and transformation of paralytic shellfish toxins in the blue mussel Mytilus galloprovincialis. Toxicon 2003, 42, 777–784. [Google Scholar] [CrossRef]
- DeGrasse, S.; Vanegas, C.; Conrad, S. Paralytic shellfish toxins in the sea scallop Placopecten magellanicus on Georges Bank: Implications for an offshore roe-on and whole scallop fishery. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 301–307. [Google Scholar] [CrossRef]
- Montoya, N.; Akselman, R.; Franco, J.; Carreto, J.I. Paralytic shellfish toxins and mackerel (Scomber japonicus) mortality in the Argentine Sea. Harmful Toxic Algal Bloom. 1996, 417–420. [Google Scholar]
- Ben-Gigirey, B.; Rossignoli, A.E.; Riobo, P.; Rodriguez, F. First Report of Paralytic Shellfish Toxins in Marine Invertebrates and Fish in Spain. Toxins 2020, 12, 723. [Google Scholar] [CrossRef]
- Reis Costa, P. Impact and effects of paralytic shellfish poisoning toxins derived from harmful algal blooms to marine fish. Fish Fish. 2016, 17, 226–248. [Google Scholar] [CrossRef]
- Abbott, J.P.; Flewelling, L.J.; Landsberg, J.H. Saxitoxin monitoring in three species of Florida puffer fish. Harmful Algae 2009, 8, 343–348. [Google Scholar] [CrossRef]
- Hernandez, M.; Robinson, I.; Aguilar, A.; Gonzalez, L.M.; Lopez-Jurado, L.F.; Reyero, M.; Cacho, E. Did algal toxins cause monk seal mortality? Nature 1998, 393, 28–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.; Lacaze, J.; Hermann, G.; Kershaw, J.; Brownlow, A.; Turner, A.; Hall, A. Toxicon Detection and effects of harmful algal toxins in Scottish harbour seals and potential links to population decline. Toxicon 2015, 97, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kvitek, R.G.; DeGange, A.R.; Beitler, M.K. Paralytic shellfish poisoning toxins mediate feeding behavior of sea otters. Limnol. Ocean. 1991, 36, 393–404. [Google Scholar] [CrossRef]
- Durbin, E.; Teegarden, G.; Campbell, R.; Cembella, A.; Baumgartner, M.F.; Mate, B.R. North Atlantic right whales, Eubalaena glacialis, exposed to paralytic shellfish poisoning (PSP) toxins via a zooplankton vector, Calanus finmarchicus. Harmful Algae 2002, 1, 243–251. [Google Scholar] [CrossRef]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Botana, A.; Botana, L.M.; Vasconcelos, V. Paralytic Shellfish Toxins Occurrence in Non-Traditional Invertebrate Vectors from North AtlanticWaters (Azores, Madeira, and Morocco). Toxins 2018, 10, 362. [Google Scholar] [CrossRef] [Green Version]
- García, C.; Pérez, F.; Contreras, C.; Figueroa, D.; Barriga, A.; López-Rivera, A.; Araneda, O.F.; Contreras, H.R. Saxitoxins and okadaic acid group: Accumulation and distribution in invertebrate marine vectors from Southern Chile. Food Addit. Contam.—Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 984–1002. [Google Scholar] [CrossRef]
- Silva, M.; Barreiro, A.; Rodriguez, P.; Otero, P.; Azevedo, J.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New invertebrate vectors for PST, spirolides and okadaic acid in the North Atlantic. Mar. Drugs 2013, 11, 1936–1960. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Dhanji-Rapkova, M.; Dean, K.; Milligan, S.; Hamilton, M.; Thomas, J.; Poole, C.; Haycock, J.; Spelman-Marriott, J.; Watson, A.; et al. Fatal canine intoxications linked to the presence of saxitoxins in stranded marine organisms following winter storm activity. Toxins 2018, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Cefas Harmful Algal Blooms (HABS) Surveillance Programmes and Monitoring. Available online: https://www.cefas.co.uk/data-and-publications/habs/ (accessed on 1 June 2021).
- Lin, H.; Cho, Y.; Yashiro, H.; Yamada, T.; Oshima, Y. Purification and characterization of paralytic shellfish toxin transforming enzyme from Mactra chinensis. Toxicon 2004, 44, 657–668. [Google Scholar] [CrossRef]
- Cho, Y.; Ogawa, N.; Takahashi, M.; Lin, H.; Oshima, Y. Purification and characterization of paralytic shellfish toxin-transforming enzyme, sulfocarbamoylase I, from the Japanese bivalve Peronidia venulosa. Biochim. Biophys. Acta 2008, 1784, 1277–1285. [Google Scholar] [CrossRef]
- Dean, K.J.; Hatfield, R.G.; Lee, V.; Alexander, R.P.; Lewis, A.M.; Maskrey, B.H.; Alves, M.T.; Hatton, B.; Coates, L.N.; Capuzzo, E.; et al. Multiple New Paralytic Shellfish Toxin Vectors in Off shore North Sea Benthos, a Deep Secret Exposed. Mar. Drugs 2020, 18, 400. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, M.; Nishimura, F.; Miyazawa, K.; Noguchi, T. Occurance of paralytic shellfish poison in the starfish Asteria amurensis in Kure Bay, Hiroshima prefecture, Japan. Toxicon 1997, 37, 1081–1087. [Google Scholar] [CrossRef]
- Ito, K.; Asakawa, M.; Sida, Y.; Miyazawa, K. Occurrence of paralytic shellfish poison (PSP) in the starfish Asterina pectinifera collected from the Kure Bay, Hiroshima Prefecture, Japan. Toxicon 2003, 41, 291–295. [Google Scholar] [CrossRef]
- Lin, S.-J.; Tsai, Y.-H.; Lin, H.; Hwang, D. Paralytic toxins in Taiwanese starfish Astrpecten scoparius. Toxicon 1998, 36, 799–803. [Google Scholar] [CrossRef]
- Ferrer, R.P.; Lunsford, E.T.; Candido, C.M.; Strawn, M.L.; Pierce, K.M. Saxitoxin and the ochre sea star: Molecule of keystone significance and a classic keystone species. Integr. Comp. Biol. 2015, 55, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, J.R.; Rogers, S.I. The distribution, relative abundance and diversity of echinoderms in the eastern English Channel, Bristol Channel, and Irish Sea. J. Mar. Biol. Assoc. UK 2000, 80, 127–138. [Google Scholar] [CrossRef]
- Ringvold, H.; Moum, T. On the genus Crossaster (Echinodermata: Asteroidea) and its distribution. PLoS ONE 2020, 15, e0227223. [Google Scholar]
- Birkeland, C. Interactions between a Sea Pen and Seven of Its Predators. Ecol. Monogr. 1974, 44, 211–232. [Google Scholar] [CrossRef]
- Carlson, H.R.; Pfister, C.A. A seventeen-year study of the rose star Crossaster papposus population in a coastal bay in southeast Alaska. Mar. Biol. 1999, 133, 223–230. [Google Scholar] [CrossRef]
- Hancock, D.A. The feeding behaviour of starfish on Essex oyster beds. J. Mar. Biol. Assoc. UK 1955, 34, 313–331. [Google Scholar] [CrossRef] [Green Version]
- Sloan, N.A. Starfish encounters: An experimental study of its advantages. Experientia 1979, 35, 1314–1315. [Google Scholar] [CrossRef]
- Himmelman, J.H.; Dutil, C. Distribution, population structure and feeding of subtidal seastars in the northern Gulf of St Lawrence. Mar. Ecol. Prog. Ser. 1991, 76, 61–72. [Google Scholar] [CrossRef]
- Hancock, D.A. Some aspects of the biology of the sunstar Crossaster papposus (L.). Ophelia 1974, 13, 1–30. [Google Scholar] [CrossRef]
- Sloan, N.A.; Northway, S.M. Chemoreception by the asteroid Crossaster papposus (L.). J. Exp. Mar. Bio. Ecol. 1982, 61, 85–98. [Google Scholar] [CrossRef]
- Feder, M.; Christianson, A. Aspects of Asteroid biology. In Physiology of Echinodermata; Inderscience Publishers: Hoboken, NJ, USA, 1966; pp. 88–127. [Google Scholar]
- Andersson, L.; Bohlin, L.; Iorizzi, M.; Riccio, R.; Minale, L.; Moreno-Lopez, W. Biological Activity of Saponins and Saponin-like Compounds from Starfish and Brittle-stars. Toxicon 1989, 27, 179–188. [Google Scholar] [CrossRef]
- Choi, M.C.; Yu, P.K.N.; Hsieh, D.P.H.; Lam, P.K.S. Trophic transfer of paralytic shellfish toxins from clams (Ruditapes philippinarum) to gastropods (Nassarius festivus). Chemosphere 2006, 64, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Villagrán, P.V.; Navarro, J.M.; Aliste, S.; Chaparro, O.R.; Ortíz, A. Trophic transfer of paralytic shellfish toxin (PST): Physiological and reproductive effects in the carnivorous gastropod Acanthina monodon (Pallas, 1774). Aquat. Toxicol. 2019, 212, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.J.; Wang, D.Z.; Niu, T.; Xu, Y.X. Trophic transfer of paralytic shellfish toxins from the cladoceran (Moina mongolica) to larvae of the fish (Sciaenops ocellatus). Toxicon 2007, 50, 639–645. [Google Scholar] [CrossRef]
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2010, 10, 113–390. [Google Scholar] [CrossRef]
- White, A.W. Sensitivity of marine fishes to toxins from the red-tide dinoflagellate Gonyaulax excavata and implications for fish kills. Mar. Biol. 1981, 65, 255–260. [Google Scholar] [CrossRef]
- Li, S.; Wang, W.; Hsieh, D.P.H. Effects of toxic dinoflagellate Alexandrium tamarense on the energy budgets and growth of two marine bivalves. Mar. Environ. Res. 2002, 53, 145–160. [Google Scholar] [CrossRef]
- Cisternas, B.; Lo, J.A.; Navarro, J.M.; Gonza, K.; Segura, C.J.; Co, M. Contrasting Physiological Responses of Two Populations of the Razor Clam Tagelus dombeii with Different Histories of Exposure to Paralytic Shellfish Poisoning. PLoS ONE 2014, 9, 1–13. [Google Scholar]
- Bernardi Bif, M.; Yunes, J.S.; Resgalla, C. Evaluation of mysids and sea urchins exposed to saxitoxins. Environ. Toxicol. Pharmacol. 2013, 36, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Fraser, S.M. Seabirds Caused By Paralytic Shellfish Poison. Br. Birds 1968, 61, 1–404. [Google Scholar]
- Shumway, S.E.; Allen, S.M.; Boersma, P.D. Marine birds and harmful algal blooms: Sporadic victims or under-reported events? Harmful Algae 2003, 2, 1–17. [Google Scholar] [CrossRef]
- Van Hemert, C.; Schoen, S.K.; Litaker, R.W.; Smith, M.M.; Arimitsu, M.L.; Piatt, J.F.; Holland, W.C.; Ransom Hardison, D.; Pearce, J.M. Algal toxins in Alaskan seabirds: Evaluating the role of saxitoxin and domoic acid in a large-scale die-off of Common Murres. Harmful Algae 2020, 92, 101730. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gigirey, B.; Soliño, L.; Bravo, I.; Rodríguez, F.; Casero, M.V.M. Paralytic and amnesic shellfish toxins impacts on seabirds, analyses and management. Toxins 2021, 13, 454. [Google Scholar] [CrossRef] [PubMed]
- Dean, K.J.; Hatfield, R.G.; Turner, A.D. Performance Characteristics of refined LC-FLD and HILIC-MS/MS methods for the Determination of Paralytic Shellfish Toxins in Shrimp, Whelk and Crab. J. AOAC Int. 2021, 104, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Harwood, D.T.; Selwood, A.I.; Van Ginkel, R.V.; Waugh, C.; McNabb, P.S.; Munday, R.; Hay, B.; Thomas, K.; Quilliam, M.A.; Malhi, N.; et al. Paralytic shellfish toxins, including deoxydecarbamoyl-STX, in wild-caught tasmanian abalone (Haliotis rubra). Toxicon 2014, 90, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Quayle, D. Paralytic shellfish poisoning in British Columbia. Fish. Res. Board Can. 1969, 168, 1–68. [Google Scholar]
- Sekiguchi, K.; Sato, S.; Ogata, T.; Kaga, S.; Kodama, M. Accumulation and depuration kinetics of paralytic shellfish toxins in the scallop Patinopecten yessoensis fed Alexandrium tamarense. Mar. Ecol. Prog. Ser. 2001, 220, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Baron, R.; Couedel, M.; Joret, C.; Garen, P.; Truquet, P.; Masselin, P.; Bardouil, M.; Lassus, P. Continuous fluorescence recording as a way to improve Pacific oyster (Crassostrea gigas) models of paralytic shellfish toxin accumulation. Aquat. Living Resour. 2006, 19, 77–84. [Google Scholar] [CrossRef] [Green Version]
- White, A.W.; Shumway, S.E.; Nassif, J.; Whittaker, D. Variation in levels of paralytic shellfish toxins among individual shellfish. Toxic Phytoplankt. Bloom. Sea 1993, 11, 209. [Google Scholar]
- Natural Scotland Scotland’s Aquaculture. Available online: http://aquaculture.scotland.gov.uk/data/data.aspx (accessed on 1 June 2021).
- Hancock, D.A. Notes on starfish on an essex oyster bed. J. Mar. Biol. Assoc. UK 1958, 37, 565–589. [Google Scholar] [CrossRef] [Green Version]
- Etheridge, S.M.; Pitcher, G.C.; Roesler, C.S. Depuration and transformation of PSP toxins in the South African abalone Haliotis midae. Harmful Algae 2002, 98–101. [Google Scholar]
- Medina-Elizalde, J.; García-Mendoza, E.; Turner, A.D.; Sánchez-Bravo, Y.A.; Murillo-Martínez, R. Transformation and depuration of paralytic shellfish toxins in the geoduck clam Panopea globosa from the Northern Gulf of California. Front. Mar. Sci. 2018, 5, 1–13. [Google Scholar] [CrossRef]
- Graneli, E.; Sundstrom, B.; Edler, L.; Anderson, D.M. Uptake and distribution of PSP toxins in butter clams. In Toxic Marine Phytoplankton; Elsevier: New York, NY, USA, 1990; pp. 257–262. [Google Scholar]
- Martin, J.L.; LeGresley, M.M.; Hanke, A.R. Thirty years—Alexandrium fundyense cyst, bloom dynamics and shellfish toxicity in the Bay of Fundy, eastern Canada. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2014, 103, 27–39. [Google Scholar] [CrossRef]
- Anderson, D.M.; Stock, C.A.; Keafer, B.A.; Bronzino, A.; Thompson, B.; Mcgillicuddy, D.J.; Keller, M.; Matrai, P.A.; Martin, J. Alexandrium fundyense cyst dynamics in the Gulf of Maine. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2522–2542. [Google Scholar] [CrossRef]
- Sacilotto Detoni, A.M.; Fonseca Costa, L.D.; Pacheco, L.A.; Yunes, J.S. Toxic Trichodesmium bloom occurrence in the southwestern South Atlantic Ocean. Toxicon 2016, 110, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Shunmugam, S.; Gayathri, M.; Prasannabalaji, N.; Thajuddin, N.; Muralitharan, G. Unraveling the presence of multi-class toxins from Trichodesmium bloom in the Gulf of Mannar region of the Bay of Bengal. Toxicon 2017, 135, 43–50. [Google Scholar] [CrossRef]
- Lewis, J.; Higman, W.; Kuenstner, S. Occurrence of Alexandrium sp. Cysts in Sediments from the North East Coast of Britain. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard-Le Denn, E., Gentien, P., Macrcaillou-Le Baut, C., Eds.; Lavoisier: Paris, France, 1995; pp. 175–180. [Google Scholar]
- Brown, J.; Fernand, L.; Horsburgh, K.J.; Hill, A.E.; Read, J.W. Paralytic shellfish poisoning on the east coast of the UK in relation to seasonal density-driven circulation. J. Plankton Res. 2001, 23, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Joint, I.; Lewis, J.; Aiken, J.; Proctor, R.; Moore, G.; Higman, W.; Donald, M. Interannual variability of PSP outbreaks on the north east UK coast. J. Plankton Res. 1997, 19, 937–956. [Google Scholar] [CrossRef] [Green Version]
- Lambert, P. Sea Stars of British Columbia, Southeast Alaska and Puget Sound; UBC Press: Vancouver, BC, Canada, 2000. [Google Scholar]
- Painefilú, J.C.; Bianchi, V.A.; Krock, B.; De Anna, J.S.; Kristoff, G.; Luquet, C.M. Ecotoxicology and Environmental Safety Effects of paralytic shellfish toxins on the middle intestine of Oncorhynchus mykiss: Glutathione metabolism, oxidative status, lysosomal function and ATP-binding cassette class C (ABCC) proteins activity. Ecotoxicol. Environ. Saf. 2020, 204, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rangel, L.M.; Silva, L.H.S.; Faassen, E.J.; Lürling, M.; Ger, K.A. Copepod prey selection and grazing efficiency mediated by chemical and morphological defensive traits of cyanobacteria. Toxins 2020, 12, 465. [Google Scholar] [CrossRef]
- Abdulhussain, A.H.; Cook, K.B.; Turner, A.D.; Lewis, A.M.; Bibby, T.S.; Mayor, D.J. The Influence of the Toxin-Producing Dinoflagellate, Alexandrium catenella (1119/27), on the Survival and Reproduction of the Marine Copepod, Acartia tonsa, During Prolonged Exposure. Front. Mar. Sci. 2021, 8, 1–10. [Google Scholar] [CrossRef]
- Friess, S.L. Mode of Action of Marine Saponins on Neuromuscluar Tissues. Fed. Proc. 1972, 31, 1146–1149. [Google Scholar]
- Itoi, S.; Yoshikawa, S.; Tatsuno, R.; Suzuki, M.; Asahina, K.; Yamamoto, S.; Takanashi, S.; Takatani, T.; Arakawa, O.; Sakakura, Y.; et al. Difference in the localization of tetrodotoxin between the female and male pufferfish Takifugu niphobles, during spawning. Toxicon 2012, 60, 1000–1004. [Google Scholar] [CrossRef]
- Itoi, S.; Yoshikawa, S.; Asahina, K.; Suzuki, M.; Ishizuka, K.; Takimoto, N.; Mitsuoka, R.; Yokoyama, N.; Detake, A.; Takayanagi, C.; et al. Larval pufferfish protected by maternal tetrodotoxin. Toxicon 2014, 78, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.A.; Taylor, D.I.; McNabb, P.; Walker, J.; Adamson, J.; Cary, S.C. Tetrodotoxin concentrations in Pleurobranchaea maculata: Temporal, spatial and individual variability from New Zealand Populations. Mar. Drugs 2012, 10, 163–176. [Google Scholar] [CrossRef]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, L.A.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculata, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 56, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Mayo, P. Ecological, Behavioural and Biochemical Studies of Aviodance Responses in Sea-Stars; University of Aberdeen: Aberdeen, UK, 1975. [Google Scholar]
- Freitas, R.; Marques, F.; De Marchi, L.; Vale, C.; Botelho, M.J. Biochemical performance of mussels, cockles and razor shells contaminated by paralytic shellfish toxins. Environ. Res. 2020, 188, 109846. [Google Scholar] [CrossRef] [PubMed]
- Kvitek, R.; Bretz, C. Harmful algal bloom toxins protect bivalve populations from sea otter predation. Mar. Ecol. Prog. Ser. 2004, 271, 233–243. [Google Scholar] [CrossRef]
- Kvitek, R.G. Paralytic shellfish toxins sequestered by bivalves as a defense against siphon-nipping fish. Mar. Biol. 1991, 111, 369–374. [Google Scholar] [CrossRef]
- AOAC. AOAC Official Method 2005.06 Paralytic Shellfish Poisoning Toxins in Shellfish Prechromatographic Oxidation and Liquid Chromatography with Fluorescence Detection. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-laboratory validation of a multitoxin ultra- performance LC-hydrophilic interaction LC-MS/MS method for quantitation of paralytic shellfish toxins in bivalve shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Dhanji-Rapkova, M.; Baker, C.; Algoet, M. Assessment of a Semiquantitative Liquid Chromatography- Fluorescence Detection Method for the Determination of Paralytic Shellfish Poisoning Toxin Levels in Bivalve Molluscs from Great Britain. J. AOAC Int. 2014, 97, 492–497. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Punn, R.; Algoet, M.; Turner, A.D. A rapid method for the analysis of paralytic shellfish toxins utilizing standard pressure HPLC: Refinement of AOAC 2005.06. J. AOAC Int. 2016, 99, 475–480. [Google Scholar] [CrossRef]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; Mcnabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography—Mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef]
- Thomas, K.M.; Beach, D.G.; Reeves, K.L.; Gibbs, R.S.; Kerrin, E.S.; McCarron, P.; Quilliam, M.A. Hydrophilic interaction liquid chromatography-tandem mass spectrometry for quantitation of paralytic shellfish toxins: Validation and application to reference materials. Anal. Bioanal. Chem. 2017, 409, 5675–7687. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, N. Distribution Expansion and Historical Population Outbreak Patterns of Crown-of-Thorns Starfish, Acanthaster planci sensu lato, in Japan from 1912 to 2015. In Coral Reef Studies of Japan 2018; Springer: Singapore, 2018; pp. 125–148. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team nlme: Linear and Nonlinear Mixed Effects Models; R Foundation: Vienna, Austria, 2018. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavent, M.; Kuentz-Simonet, V.; Labenne, A.; Saracco, J. Multivariate Analysis of Mixed Data: The R Package PCAmixdata. arXiv 2014, arXiv:1411.4911. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dean, K.J.; Alexander, R.P.; Hatfield, R.G.; Lewis, A.M.; Coates, L.N.; Collin, T.; Teixeira Alves, M.; Lee, V.; Daumich, C.; Hicks, R.; et al. The Common Sunstar Crossaster papposus—A Neurotoxic Starfish. Mar. Drugs 2021, 19, 695. https://0-doi-org.brum.beds.ac.uk/10.3390/md19120695
Dean KJ, Alexander RP, Hatfield RG, Lewis AM, Coates LN, Collin T, Teixeira Alves M, Lee V, Daumich C, Hicks R, et al. The Common Sunstar Crossaster papposus—A Neurotoxic Starfish. Marine Drugs. 2021; 19(12):695. https://0-doi-org.brum.beds.ac.uk/10.3390/md19120695
Chicago/Turabian StyleDean, Karl J., Ryan P. Alexander, Robert G. Hatfield, Adam M. Lewis, Lewis N. Coates, Tom Collin, Mickael Teixeira Alves, Vanessa Lee, Caroline Daumich, Ruth Hicks, and et al. 2021. "The Common Sunstar Crossaster papposus—A Neurotoxic Starfish" Marine Drugs 19, no. 12: 695. https://0-doi-org.brum.beds.ac.uk/10.3390/md19120695