
Highlighting the Biotechnological Potential of Deep Oceanic Crust Fungi through the Prism of Their Antimicrobial Activity

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
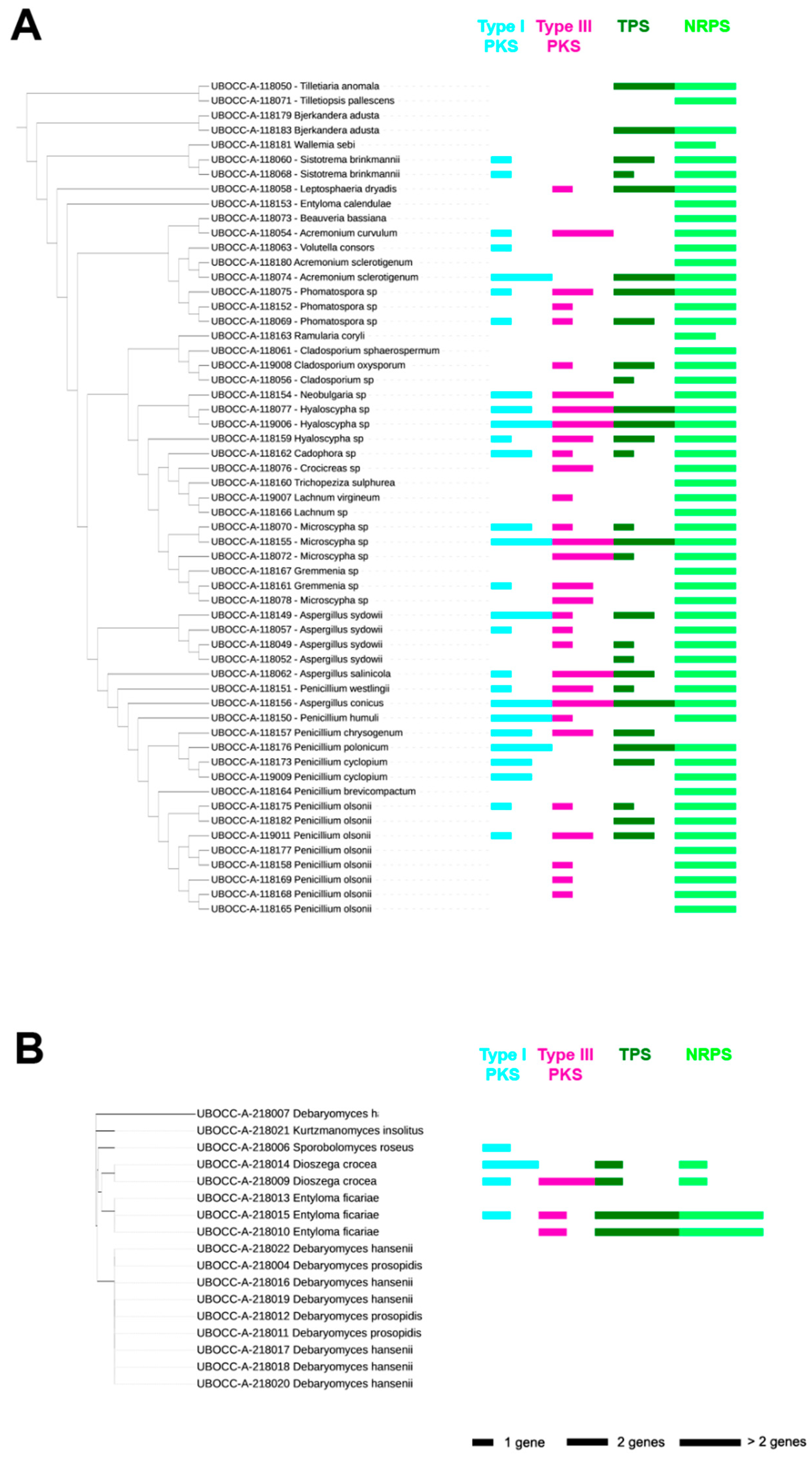
2.1. Presence of Genes Involved in the Production of Secondary Metabolites
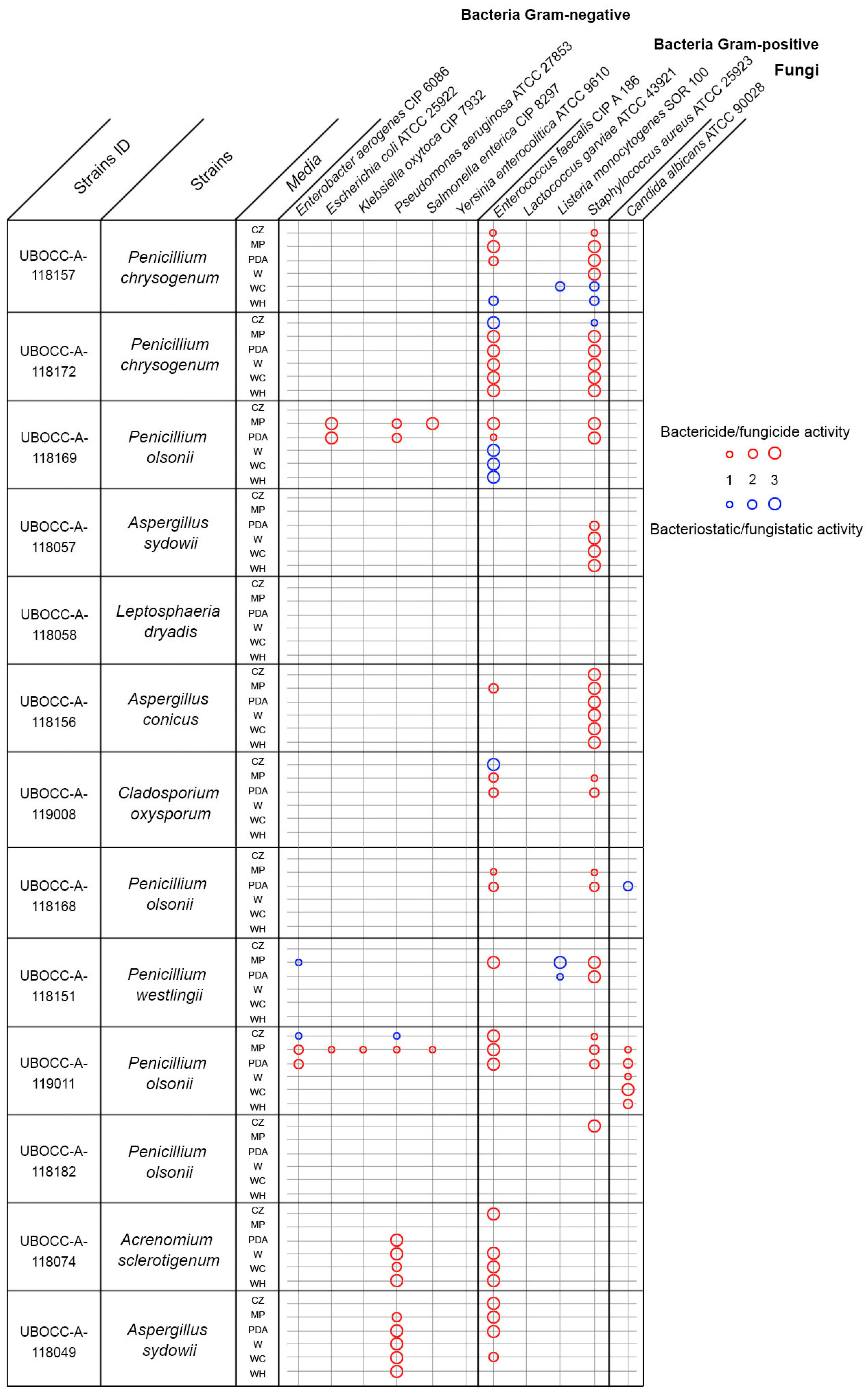
2.2. Antimicrobial Activities
2.3. Antimicrobial Activity against Multidrug-Resistant Bacteria
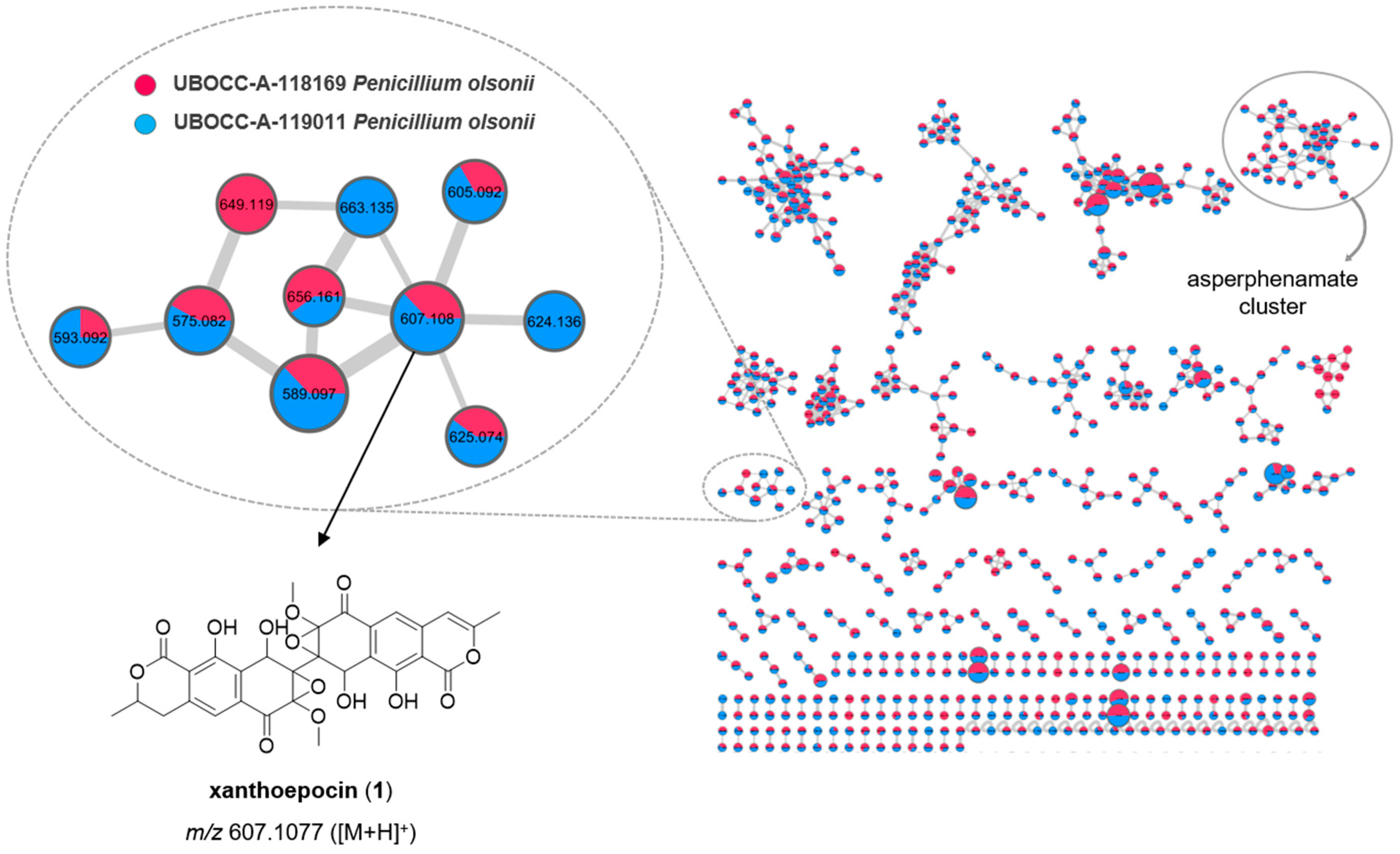
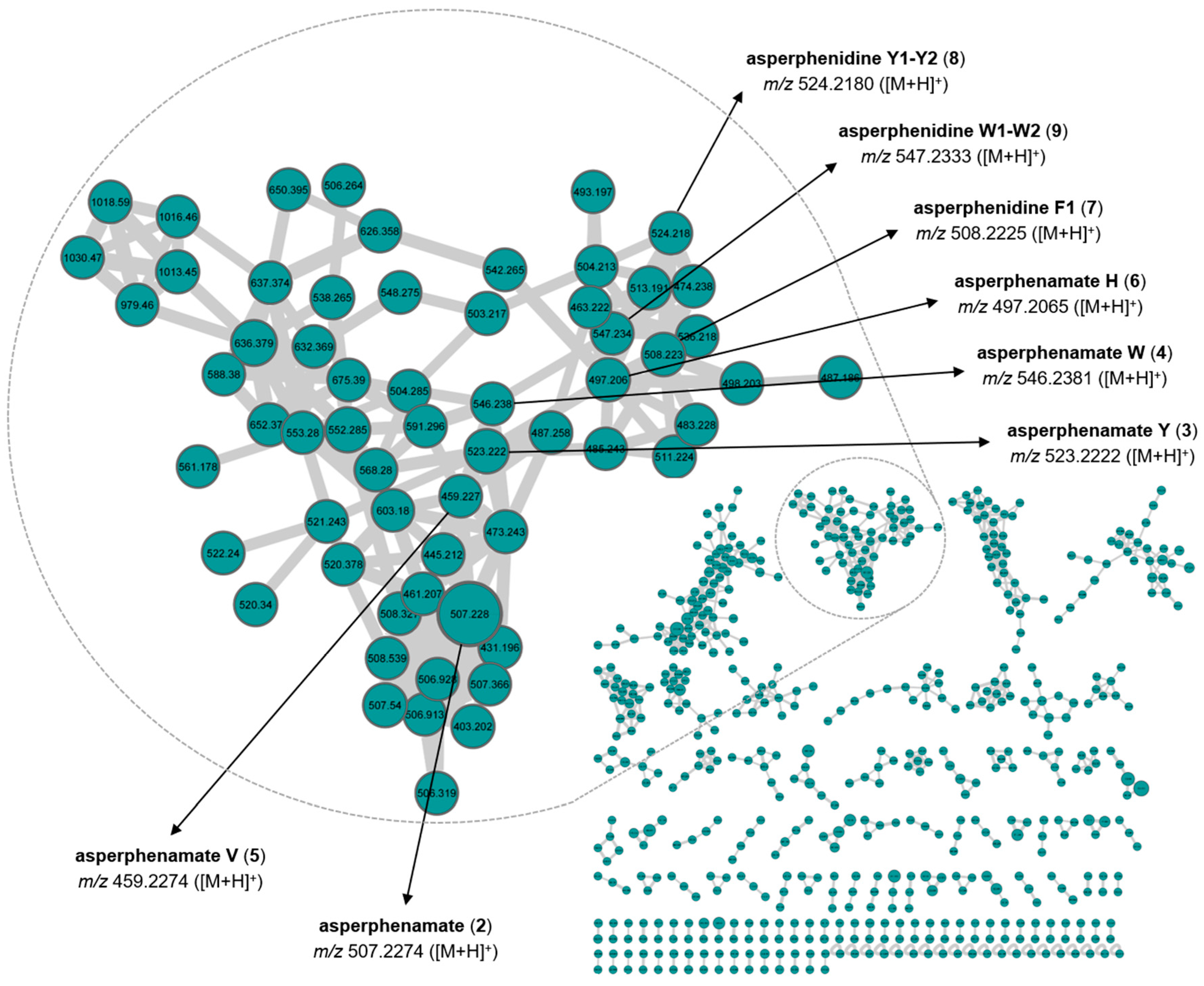
2.4. Molecular Networking
3. Materials and Methods
3.1. Culture Collection
3.2. Occurrence of Genes Involved in Secondary Metabolite Anabolism
3.3. First Screening—Agar Plug Diffusion Assay
3.4. Secondary Screening—Metabolite Extraction, Molecular Networking and Determination of the Minimum Inhibitory Concentration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The Amazing Potential of Fungi: 50 Ways We Can Exploit Fungi Industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Bills, G.F.; Gloer, J.B. Biologically Active Secondary Metabolites from the Fungi. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Calvez, T.L. Diversité et Fonctions Ecologiques des Champignons en Ecosystème Hydrothermal Marin Profond. Ph.D. Thesis, Université de Rennes 1, Rennes, France, 2009. [Google Scholar]
- Burgaud, G.; Calvez, T.; Arzur, D.; Vandenkoornhuyse, P.; Barbier, G. Diversity of Culturable Marine Filamentous Fungi from Deep-Sea Hydrothermal Vents. Environ. Microbiol. 2009, 11, 1588–1600. [Google Scholar] [CrossRef] [PubMed]
- Edgcomb, V.P.; Beaudoin, D.; Gast, R.; Biddle, J.F.; Teske, A. Marine Subsurface Eukaryotes: The Fungal Majority. Environ. Microbiol. 2011, 13, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Orsi, W.D.; Edgcomb, V.P.; Christman, G.D.; Biddle, J.F. Gene Expression in the Deep Biosphere. Nature 2013, 499, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Rédou, V.; Ciobanu, M.C.; Pachiadaki, M.G.; Edgcomb, V.; Alain, K.; Barbier, G.; Burgaud, G. In-Depth Analyses of Deep Subsurface Sediments Using 454-Pyrosequencing Reveals a Reservoir of Buried Fungal Communities at Record-Breaking Depths. FEMS Microbiol. Ecol. 2014, 90, 908–921. [Google Scholar] [CrossRef] [Green Version]
- Rédou, V.; Navarri, M.; Meslet-Cladière, L.; Barbier, G.; Burgaud, G. Species Richness and Adaptation of Marine Fungi from Deep-Subseafloor Sediments. Appl. Environ. Microbiol. 2015, 81, 3571–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachiadaki, M.G.; Rédou, V.; Beaudoin, D.J.; Burgaud, G.; Edgcomb, V.P. Fungal and Prokaryotic Activities in the Marine Subsurface Biosphere at Peru Margin and Canterbury Basin Inferred from RNA-Based Analyses and Microscopy. Front. Microbiol. 2016, 7, 846. [Google Scholar] [CrossRef]
- Li, J.; Mara, P.; Schubotz, F.; Sylvan, J.B.; Burgaud, G.; Klein, F.; Beaudoin, D.; Wee, S.Y.; Dick, H.J.B.; Lott, S.; et al. Recycling and Metabolic Flexibility Dictate Life in the Lower Oceanic Crust. Nature 2020, 579, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Quemener, M.; Mara, P.; Schubotz, F.; Beaudoin, D.; Li, W.; Pachiadaki, M.; Sehein, T.R.; Sylvan, J.B.; Li, J.; Barbier, G.; et al. Meta-Omics Highlights the Diversity, Activity and Adaptations of Fungi in Deep Oceanic Crust. Environ. Microbiol. 2020, 22, 3950–3967. [Google Scholar] [CrossRef] [PubMed]
- Nagano, Y.; Miura, T.; Nishi, S.; Lima, A.O.; Nakayama, C.; Pellizari, V.H.; Fujikura, K. Fungal Diversity in Deep-Sea Sediments Associated with Asphalt Seeps at the Sao Paulo Plateau. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 146, 59–67. [Google Scholar] [CrossRef]
- Orsi, W.D. Ecology and Evolution of Seafloor and Subseafloor Microbial Communities. Nat. Rev. Microbiol. 2018, 16, 671–683. [Google Scholar] [CrossRef]
- Karagiosis, S.A.; Baker, S.E. Fungal Cell Factories. In Food Bioproducts Processing; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 205–219. ISBN 978-1-119-94608-3. [Google Scholar]
- Petersen, L.-E.; Kellermann, M.Y.; Schupp, P.J. Secondary Metabolites of Marine Microbes: From Natural Products Chemistry to Chemical Ecology. In YOUMARES 9—The Oceans: Our Research, Our Future: Proceedings of the 2018 Conference for Young Marine Researcher in Oldenburg, Germany; Jungblut, S., Liebich, V., Bode-Dalby, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 159–180. ISBN 978-3-030-20389-4. [Google Scholar]
- Daletos, G.; Ebrahim, W.; Ancheeva, E.; El-Neketi, M.; Song, W.; Lin, W.; Proksch, P. Natural Products from Deep-Sea-Derived Fungi A New Source of Novel Bioactive Compounds? Curr. Med. Chem. 2018, 25, 186–207. [Google Scholar] [CrossRef]
- Hasan, S.; Ansari, M.I.; Ahmad, A.; Mishra, M. Major Bioactive Metabolites from Marine Fungi: A Review. Bioinformation 2015, 11, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Legido-Quigley, H.; Hsu, L.Y. Antimicrobial Resistance in One Health. In Global Health Security: Recognizing Vulnerabilities, Creating Opportunities; Masys, A.J., Izurieta, R., Reina Ortiz, M., Eds.; Advanced Sciences and Technologies for Security Applications; Springer International Publishing: Cham, Switzerland, 2020; pp. 209–229. ISBN 978-3-030-23491-1. [Google Scholar]
- van Hengel, A.J.; Marin, L. Research, Innovation, and Policy: An Alliance Combating Antimicrobial Resistance. Trends Microbiol 2019, 27, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life v2: Online Annotation and Display of Phylogenetic Trees Made Easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.M.; Kim, M.J.; Li, H.; Zhang, P.; Bao, B.; Lee, K.J.; Jung, J.H. Marine-Derived Aspergillus Species as a Source of Bioactive Secondary Metabolites. Mar. Biotechnol. 2013, 15, 499–519. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-G.; Liu, Q.; Zhu, G.-L.; Liu, H.-S.; Zhu, W.-M. Marine Natural Products Sourced from Marine-Derived Penicillium Fungi. J. Asian Nat. Prod. Res. 2016, 18, 92–115. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, H.; Zhu, W. New natural products from the marine-derived Aspergillus fungi—A review. Wei Sheng Wu Xue Bao 2016, 56, 331–362. [Google Scholar]
- Farha, A.K.; Hatha, A.M. Bioprospecting Potential and Secondary Metabolite Profile of a Novel Sediment-Derived Fungus Penicillium Sp. ArCSPf from Continental Slope of Eastern Arabian Sea. Mycology 2019, 10, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tang, G.; Xu, X.; Nong, X.; Qi, S. Insights into Deep-Sea Sediment Fungal Communities from the East Indian Ocean Using Targeted Environmental Sequencing Combined with Traditional Cultivation. PLoS ONE 2014, 9, e109118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boettger, D.; Hertweck, C. Molecular Diversity Sculpted by Fungal PKS-NRPS Hybrids. Chembiochem 2013, 14, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Navarri, M.; Jégou, C.; Meslet-Cladière, L.; Brillet, B.; Barbier, G.; Burgaud, G.; Fleury, Y. Deep Subseafloor Fungi as an Untapped Reservoir of Amphipathic Antimicrobial Compounds. Mar. Drugs 2016, 14, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, N.; Turner, G.; Bennett, J. Fungal Secondary Metabolism—From Biochemistry to Genomics. Nat. Rev. Microbiol. 2006, 3, 937–947. [Google Scholar] [CrossRef]
- Duarte, K.; Rocha-Santos, T.A.P.; Freitas, A.C.; Duarte, A.C. Analytical Techniques for Discovery of Bioactive Compounds from Marine Fungi. TrAC Trends Anal. Chem. 2012, 34, 97–110. [Google Scholar] [CrossRef]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and Role of Fungal Secondary Metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal Secondary Metabolism: Regulation, Function and Drug Discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. AntiSMASH 3.0—A Comprehensive Resource for the Genome Mining of Biosynthetic Gene Clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Duran, H.G.S.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. AntiSMASH 4.0-Improvements in Chemistry Prediction and Gene Cluster Boundary Identification. Nucleic Acids Res. 2017, 45, 36–41. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the Secondary Metabolite Genome Mining Pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Rédou, V.; Kumar, A.; Hainaut, M.; Henrissat, B.; Record, E.; Barbier, G.; Burgaud, G. Draft Genome Sequence of the Deep-Sea Basidiomycetous Yeast Cryptococcus Sp. Strain Mo29 Reveals Its Biotechnological Potential. Genome Announc. 2016, 4, e00461. [Google Scholar] [CrossRef] [Green Version]
- Roullier, C.; Bertrand, S.; Blanchet, E.; Peigné, M.; Robiou du Pont, T.; Guitton, Y.; Pouchus, Y.F.; Grovel, O. Time Dependency of Chemodiversity and Biosynthetic Pathways: An LC-MS Metabolomic Study of Marine-Sourced Penicillium. Mar. Drugs 2016, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Lin, Z.; Li, D.; Gu, Q.; Zhu, T. OSMAC (one strain many compounds) approach in the research of microbial metabolites—A review. Wei Sheng Wu Xue Bao 2010, 50, 701–709. [Google Scholar]
- Hewage, R.T.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. One Strain-Many Compounds (OSMAC) Method for Production of Polyketides, Azaphilones, and an Isochromanone Using the Endophytic Fungus Dothideomycete sp. Phytochemistry 2014, 108, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, B.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Influence of OSMAC-Based Cultivation in Metabolome and Anticancer Activity of Fungi Associated with the Brown Alga Fucus Vesiculosus. Mar. Drugs 2019, 17, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagestad, O.C.; Andersen, J.H.; Altermark, B.; Hansen, E.; Rämä, T. Cultivable Marine Fungi from the Arctic Archipelago of Svalbard and Their Antibacterial Activity. Mycology 2019, 11, 230–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Sun, Y.; Bao, J.; He, F.; Xu, X.; Qi, S. Phylogenetic Survey and Antimicrobial Activity of Culturable Microorganisms Associated with the South China Sea Black Coral Antipathes Dichotoma. FEMS Microbiol. Lett. 2012, 336, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, L.A.; Tanaka, N.; Kalinovskaya, N.I.; Mikhailov, V.V. Antimicrobial Potential of Deep Surface Sediment Associated Bacteria from the Sea of Japan. World J. Microbiol. Biotechnol. 2013, 29, 1169–1177. [Google Scholar] [CrossRef]
- León-Palmero, E.; Joglar, V.; Álvarez, P.A.; Martín-Platero, A.; Llamas, I.; Reche, I. Diversity and Antimicrobial Potential in Sea Anemone and Holothurian Microbiomes. PLoS ONE 2018, 13, e0196178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.M.; Hufford, C.D.; Robertson, L.W. Two Metabolites from Aspergillus Flavipes [Fungi]. Lloydia 1977, 40, 146–151. [Google Scholar] [PubMed]
- Igarashi, Y.; Kuwamori, Y.; Takagi, K.; Ando, T.; Fudou, R.; Furumai, T.; Oki, T. Xanthoepocin, a New Antibiotic from Penicillium Simplicissimum IFO5762. J. Antibiot. 2000, 53, 928–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnaweera, P.B.; Williams, D.E.; Dilip de Silva, E.; Andersen, R.J. Antibacterial Metabolites from the Sri Lankan Demosponge-Derived Fungus, Aspergillus Flavipes. Curr. Sci. 2016, 111, 1473. [Google Scholar] [CrossRef]
- Subko, K.; Wang, X.; Nielsen, F.H.; Isbrandt, T.; Gotfredsen, C.H.; Ramos, M.C.; Mackenzie, T.; Vicente, F.; Genilloud, O.; Frisvad, J.C.; et al. Mass Spectrometry Guided Discovery and Design of Novel Asperphenamate Analogs from Penicillium astrolabium Reveals an Extraordinary NRPS Flexibility. Front. Microbiol. 2021, 11, 618–730. [Google Scholar] [CrossRef]
- Yuan, L.; Li, Y.; Zou, C.; Wang, C.; Gao, J.; Miao, C.; Ma, E.; Sun, T. Synthesis and in Vitro Antitumor Activity of Asperphenamate Derivatives as Autophagy Inducer. Bioorg. Med. Chem. Lett. 2012, 22, 2216–2220. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Sheng, L.; Zou, C.; Sun, H.; Zhang, C.; Liu, Y.; Shi, J.; Ma, E.; Yuan, L. Design, Synthesis and Biological Evaluation of Novel Asperphenamate Derivatives. Eur. J. Med. Chem. 2016, 110, 76–86. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Shao, C.-L.; Wu, L.-Y.; Chen, M.; Wang, K.-L.; Zhao, D.-L.; Sun, X.-P.; Chen, G.-Y.; Wang, C.-Y. Bioactive Phenylalanine Derivatives and Cytochalasins from the Soft Coral-Derived Fungus. Aspergillus Elegans. Mar. Drugs 2013, 11, 2054–2068. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-G.; Bao, L.; Liu, H.-W.; Ren, J.-W.; Wang, W.-Z.; Wang, L.; Li, W.; Yin, W.-B. Chemical Diversity from the Tibetan Plateau Fungi Penicillium kongii and P. brasilianum. Mycology 2018, 9, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Pathogens | Temperature | Medium |
|---|---|---|
| Enterobacter aerogenes CIP 6086 | 30 °C | TSB |
| Escherichia coli ATCC 25922 | 37 °C | TSB |
| Klebsiella oxytoca CIP 7932 | 37 °C | TSB |
| Pseudomonas aeruginosa ATCC 27853 | 37 °C | TSB |
| Salmonella enterica CIP 8297 | 37 °C | TSB |
| Yersinia enterocolitica ATCC 9610 | 30 °C | TSB + NaCl |
| Enterococcus faecalis CIP A 186 | 37 °C | TSB |
| Lactococcus garviae ATCC 43921 | 30 °C | BHI |
| Listeria monocytogenes SOR 100 | 37 °C | BHI |
| Staphylococcus aureus ATCC 25923 | 37 °C | TSB |
| Candida albicans ATCC 90028 | 37 °C | Yeast medium |
| Basic Pathogens | MDR Pathogens |
|---|---|
| Enterococcus faecalis CIP A 186 | Pseudomonas aeruginosa VIM2 |
| Staphylococcus aureus ATCC 25923 | Pseudomonas aeruginosa Brse |
| Escherichia coli ATCC 25922 | Staphylococcus aureus MecC |
| Pseudomonas aeruginosa ATCC 27853 | Staphylococcus aureus MecA III |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quemener, M.; Dayras, M.; Frotté, N.; Debaets, S.; Le Meur, C.; Barbier, G.; Edgcomb, V.; Mehiri, M.; Burgaud, G. Highlighting the Biotechnological Potential of Deep Oceanic Crust Fungi through the Prism of Their Antimicrobial Activity. Mar. Drugs 2021, 19, 411. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080411
Quemener M, Dayras M, Frotté N, Debaets S, Le Meur C, Barbier G, Edgcomb V, Mehiri M, Burgaud G. Highlighting the Biotechnological Potential of Deep Oceanic Crust Fungi through the Prism of Their Antimicrobial Activity. Marine Drugs. 2021; 19(8):411. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080411
Chicago/Turabian StyleQuemener, Maxence, Marie Dayras, Nicolas Frotté, Stella Debaets, Christophe Le Meur, Georges Barbier, Virginia Edgcomb, Mohamed Mehiri, and Gaëtan Burgaud. 2021. "Highlighting the Biotechnological Potential of Deep Oceanic Crust Fungi through the Prism of Their Antimicrobial Activity" Marine Drugs 19, no. 8: 411. https://0-doi-org.brum.beds.ac.uk/10.3390/md19080411