
Ligiamycins A and B, Decalin-Amino-Maleimides from the Co-Culture of Streptomyces sp. and Achromobacter sp. Isolated from the Marine Wharf Roach, Ligia exotica

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
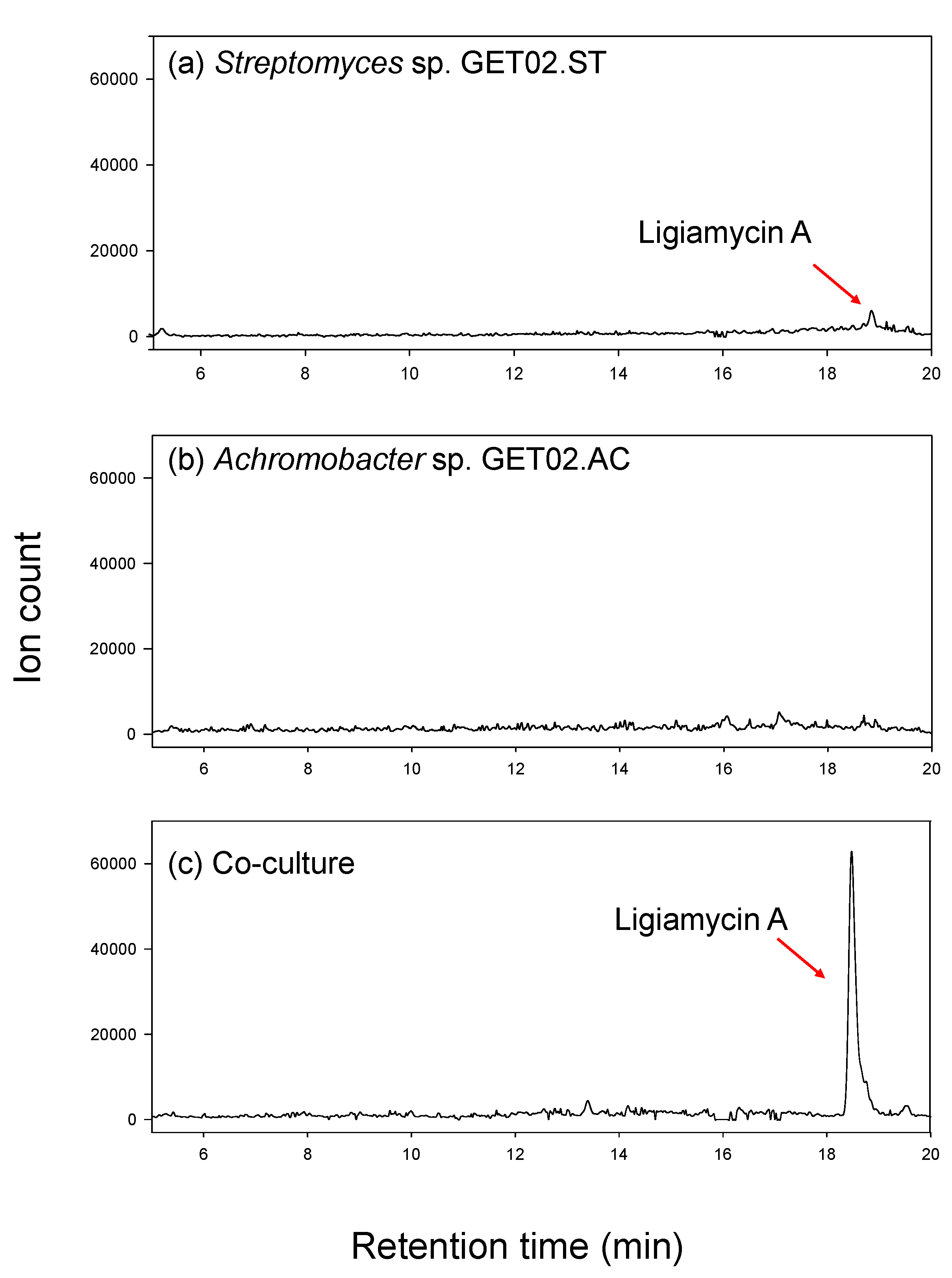
2.1. Co-Culture of Streptomyces sp. and Achromobacter sp. from Ligia exotica
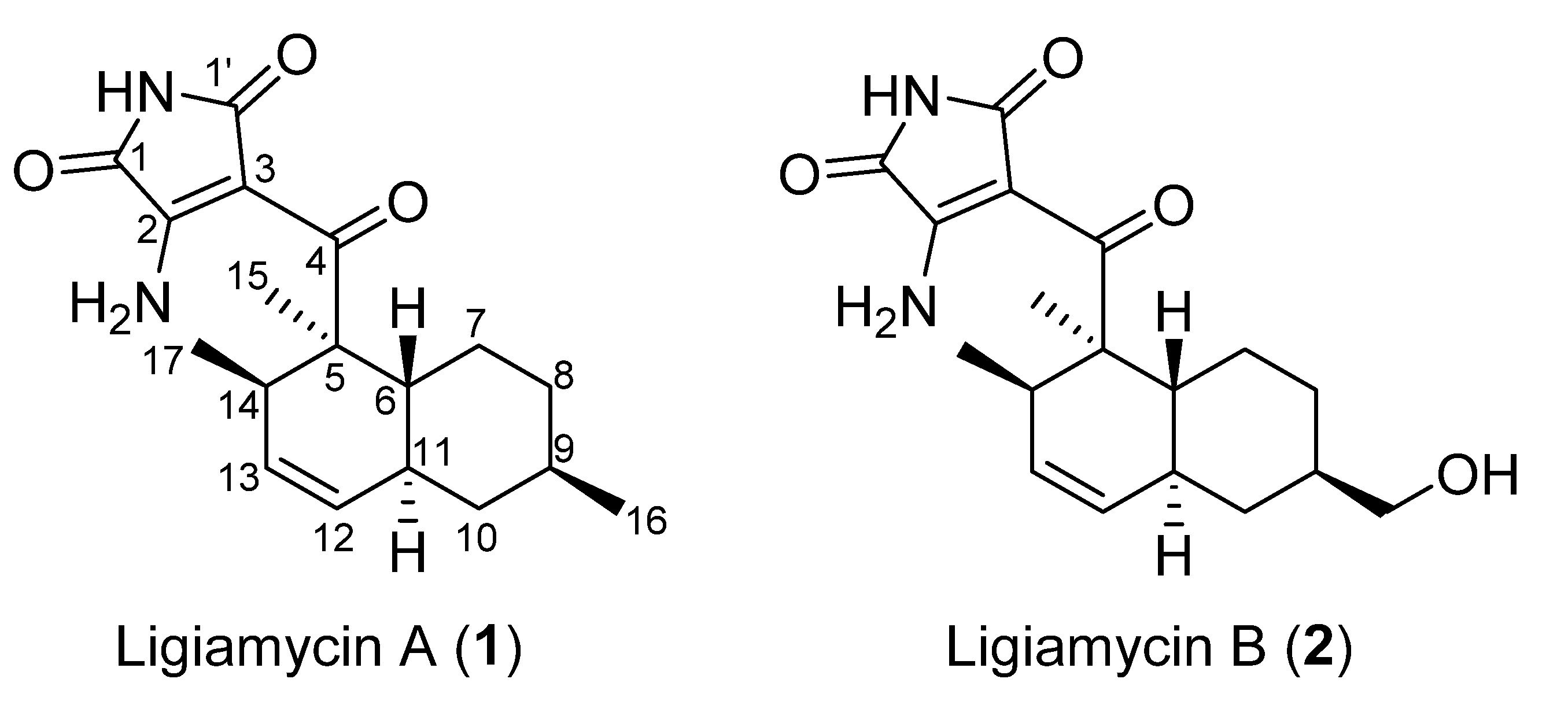
2.2. Structural Elucidation
2.3. Biological Evaluation
3. Materials and Methods
3.1. General Experimental Procedure
3.2. Collection and Bacterial Isolation of Wharf Roaches
3.3. Large-Scale Co-Culture and Extraction
3.4. Isolation and Purification of Ligiamycins
3.4.1. Ligiamycin A (1)
3.4.2. Ligiamycin B (2)
3.5. ECD and Optical Rotation Calculation
3.6. Cell Culture
3.7. Cell Proliferation Assays
3.8. Antibacterial Activity Bioassays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, P.; Ye, X.; Wei, B.; Emam, M.; Zhang, H.; Wang, H. The structural diversity of marine microbial secondary metabolites based on co-culture strategy: 2009–2019. Mar. Drugs 2020, 18, 449. [Google Scholar] [CrossRef]
- Cueto, M.; Jensen, P.R.; Kauffman, C.; Fenical, W.; Lobkovsky, E.; Clardy, J. Pestalone, a new antibiotic produced by a marine fungus in response to bacterial challenge. J. Nat. Prod. 2001, 64, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-C.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Libertellenones A–D: Induction of cytotoxic diterpenoid biosynthesis by marine microbial competition. Bioorg. Med. Chem. 2005, 13, 5267–5273. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Induced production of emericellamides A and B from the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 2007, 70, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Adnani, N.; Vazquez-Rivera, E.; Adibhatla, S.N.; Ellis, G.A.; Braun, D.R.; Bugni, T.S. Investigation of interspecies interactions within marine Micromonosporaceae using an improved co-culture approach. Mar. Drugs 2015, 13, 6082–6098. [Google Scholar] [CrossRef] [PubMed]
- Adnani, N.; Chevrette, M.G.; Adibhatla, S.N.; Zhang, F.; Yu, Q.; Braun, D.R.; Nelson, J.; Simpkins, S.W.; McDonald, B.R.; Myers, C.L.; et al. Coculture of marine invertebrate-associated bacteria and interdisciplinary technologies enable biosynthesis and discovery of a new antibiotic, keyicin. ACS Chem. Biol. 2017, 12, 3093–3102. [Google Scholar] [CrossRef]
- Shin, D.; Byun, W.S.; Moon, K.; Kwon, Y.; Bae, M.; Um, S.; Lee, S.K.; Oh, D.-C. Coculture of marine Streptomyces sp. with Bacillus sp. produces a new piperazic acid-bearing cyclic peptide. Front. Chem. 2018, 6, 498. [Google Scholar] [CrossRef]
- Hifnawy, S.M.; Hassan, H.M.; Mohammed, R.; Fouda, M.M.; Sayed, A.M.; Hamed, A.A.; AbouZid, S.F.; Rateb, M.E.; Alhadrami, H.A.; Abdelmohsen, U.R. Induction of antibacterial metabolites by co-cultivation of two red-sea-sponge-associated actinomycetes Micromonospora sp. UR56 and Actinokinespora sp. EG49. Mar. Drugs 2020, 18, 243. [Google Scholar] [CrossRef]
- Zhang, L.; Niaz, S.I.; Khan, D.; Wang, Z.; Zhu, Y.; Zhou, H.; Lin, Y.; Li, J.; Liu, L. Induction of diverse bioactive secondary metabolites from the mangrove endophytic fungus Trichoderma sp. (Strain 307) by co-cultivation with Acinetobacter johnsonii (Strain B2). Mar. Drugs 2017, 15, 35. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, C.; Lo Giudice, A. Marine invertebrates: Underexplored sources of bacteria producing biologically active molecules. Diversity 2018, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.H. Terrestrial Isopoda (Crustacea) from Korea. Korean J. Syst. Zool. 1993, 36, 133–158. [Google Scholar]
- Li, X.; Zhao, Z.; Ding, W.; Ye, B.; Wang, P.; Xu, J. Aspochalazine A, a novel polycyclic aspochalasin from the fungus Aspergillus sp. Z4. Tetrahedron Lett. 2017, 58, 2405–2408. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, S.; Yang, X. A new cyclopeptide metabolite of marine gut fungus from Ligia oceanica. Nat. Prod. Res. 2014, 28, 994–997. [Google Scholar] [CrossRef]
- Li, X.; Ding, W.; Wang, P.; Xu, J. Two novel aspochalasins from the gut fungus Aspergillus sp. Z4. Mar. Drugs 2018, 16, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-C.; Ferreira, D.; Ding, Y. Determination of absolute configuration of natural products: Theoretical calculation of electronic circular dichroism as a tool. Curr. Org. Chem. 2010, 14, 1678–1697. [Google Scholar] [CrossRef] [PubMed]
- Polavarapu, P.L. Optical rotation: Recent advances in determining the absolute configuration. Chirality 2002, 14, 768–781. [Google Scholar] [CrossRef]
- Stephens, P.J.; Pan, J.J.; Devlin, F.J.; Cheeseman, J.R. Determination of the absolute configurations of natural products using TDDFT optical rotation calculations: The iridoid oruwacin. J. Nat. Prod. 2008, 71, 285–288. [Google Scholar] [CrossRef]
- Taniguchi, T.; Martin, C.L.; Monde, K.; Nakanishi, K.; Berova, N.; Overman, L.E. Absolute configuration of actinophyllic acid as determined through chiroptical data. J. Nat. Prod. 2009, 72, 430–432. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.; An, J.S.; Hong, S.-H.; Bae, E.S.; Chung, B.; Kwon, Y.; Hong, S.; Oh, K.-B.; Shin, J.; Lee, S.K.; et al. Donghaecyclinones A–C: New cytotoxic rearranged angucyclinones from a volcanic island-derived marine Streptomyces sp. Mar. Drugs 2020, 18, 121. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Balasubramani, S.G.; Chen, G.P.; Coriani, S.; Diedenhofen, M.; Frank, M.S.; Franzke, Y.J.; Furche, F.; Grotjahn, R.; Harding, M.E.; Hättig, C.; et al. TURBOMOLE: Modular program suite for ab initio quantum-chemical and condensed-matter simulations. J. Chem. Phys. 2020, 152, 184107. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Cui, J.; Kim, E.; Riandi, E.S.; Park, S.H.; Byun, W.S.; Kal, Y.; Park, J.Y.; Hwang, S.; Shin, D.; et al. Structures and biosynthetic pathway of pulvomycins B–D: 22-membered macrolides from an estuarine Streptomyces sp. Org. Lett. 2020, 22, 5358–5362. [Google Scholar] [CrossRef] [PubMed]
- An, J.S.; Hong, S.-H.; Somers, E.; Lee, J.; Kim, B.-Y.; Woo, D.; Kim, S.W.; Hong, H.-J.; Jo, S.-I.; Shin, J.; et al. Lenzimycins A and B, metabolites with antibacterial properties from Brevibacillus sp. associated with the dung beetle Onthophagus lenzii. Front. Microbiol. 2020, 11, 599911. [Google Scholar] [CrossRef]
- Kim, D.-G.; Moon, K.; Kim, S.-H.; Park, S.-H.; Park, S.; Lee, S.K.; Oh, K.-B.; Shin, J.; Oh, D.-C. Bahamaolides A and B, antifungal polyene polyol macrolides from the marine actinomycete Streptomyces sp. J. Nat. Prod. 2012, 75, 959–967. [Google Scholar] [CrossRef]
- He, H.; Janso, J.E.; Yang, H.Y.; Singh, M.P.; Bernan, V.S.; Greenstein, M.; Carter, G.T. Oxasetin, a new antibacterial polyketide produced by fungus Vaginatispora aquatica, HK1821. J. Antibiot. 2002, 55, 821–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, W.-H.; Li, J.; Liu, Q.; Xu, T.-T.; Shi, G.-H.; Yu, H.-B.; Yang, F.; Han, B.-N.; Li, M.; Lin, H.-W. Dysidinoid A, an unusual meroterpenoid with anti-MRSA activity from the south China sea sponge Dysidea sp. Molecules 2014, 19, 18025–18032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.; Dander, J.E.; Sato, C.; Hung, Y.-S.; Gao, S.-S.; Tang, M.-C.; Hang, L.; Winter, J.M.; Garg, N.K.; Watanabe, K.; et al. Collaborative biosynthesis of maleimide- and succinimide-containing natural products by fungal polyketide megasynthases. J. Am. Chem. Soc. 2017, 139, 5317–5320. [Google Scholar] [CrossRef] [Green Version]
- Perel’son, M.E.; Aleksandrov, G.G.; Yakhontova, L.D.; Tolkachev, O.N.; Fesenko, D.A.; Komarova, M.N.; Esipov, S.E. Alkaloids of Hypecoum erectum, the structure of hyperectine. Chem. Nat. Compd. 1984, 20, 592–598. [Google Scholar] [CrossRef]
- Yakhontova, L.D.; Yartseva, I.V.; Klyuev, N.A.; Tolkachev, O.N. Structure of isohyperectine—An alkaloid from Hypecoum erectum. Chem. Nat. Compd. 1993, 29, 744–747. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Rücker, G.; Breitmaier, E.; Mayer, R. Alkaloids from Hypecoum leptocarpum. Phytochemistry 1995, 40, 1813–1816. [Google Scholar]
- Shank, E.A.; Kolter, R. New developments in microbial interspecies signaling. Curr. Opin. Microbiol. 2009, 12, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigali, S.; Titgemeyer, F.; Barends, S.; Mulder, S.; Thomae, A.W.; Hopwood, D.A.; van Wezel, G.P. Feast or famine: The global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Rep. 2008, 9, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligiamycin A (1) a | Ligiamycin B (2) b | |||
|---|---|---|---|---|
| Position | δC/δN, Type | δH, Mult (J in Hz) | δC, Type | δH, Mult (J in Hz) |
| 1 | 165.1, C | 165.2, C | ||
| 1’ | 169.8, C | 169.9, C | ||
| 2 | 157.8, C | 157.8, C | ||
| 3 | 98.7, C | 98.7, C | ||
| 4 | 200.0, C | 200.0, C | ||
| 5 | 50.1, C | 50.1, C | ||
| 6 | 39.3, CH | 1.55, t (10.0) | 39.6, CH | 1.56, t (10.0) |
| 7a | 27.2, CH2 | 1.88, m | 26.8, CH2 | 1.92, m |
| 7b | 0.88, m c | 0.86, m | ||
| 8a | 35.7, CH2 | 1.70, m | 30.3, CH2 | 1.75, m |
| 8b | 1.00, ddd (16.0, 12.5, 3.5) | 0.99, m | ||
| 9 | 32.9, CH | 1.48, m | 40.9, CH | 1.47, m |
| 10a | 42.1, CH2 | 1.77, m | 36.8, CH2 | 1.84, m |
| 10b | 0.78, m | 0.76, m | ||
| 11 | 38.0, CH | 1.75, m | 37.7, CH2 | 1.73, m |
| 12 | 129.0, CH | 5.31, dd (10.0, 2.0) | 129.1, CH | 5.33, dd (10.0, 2.0) |
| 13 | 130.2, CH | 5.52, m | 130.2, CH | 5.52, m |
| 14 | 34.6, CH | 3.06, m | 34.6, CH | 3.06, m |
| 15 | 14.8, CH3 | 1.27, s | 14.8, CH3 | 1.28, s |
| 16 | 22.5, CH3 | 0.88, m c | 66.5, CH2 | 3.22, d (6.0) |
| 17 | 18.5, CH3 | 0.68, d (7.0) | 18.5, CH3 | 0.68, d (6.5) |
| 1,1’-NH | 146.8 d, NH | 10.6, br s | 10.6, br s | |
| 2-NH2a | 103.9 d, NH2 | 9.16, s | 9.16, s | |
| 2-NH2b | 8.90, br s | 8.89, br s | ||
| 16-OH | 4.36, br s | |||
| Sample | Optical Rotation | ||
|---|---|---|---|
| Experimental | Calculated of 5R, 6R, 9R, 11S, 14R | Calculated of 5S, 6S, 9S, 11R, 14S | |
| Ligiamycin A (1) | −78 | −65 | +66 |
| Ligiamycin B (2) | −67 | −64 | +64 |
| Sample | MIC (μg/mL) | |||||
|---|---|---|---|---|---|---|
| Gram-Positive | Gram-Negative | |||||
| S. aureus | E. faecalis | E. faecium | K. pneumonia | S. enterica | E. coli | |
| Ligiamycin A (1) | 16 | >128 | >128 | >128 | 16 | >128 |
| Ligiamycin B (2) | 64 | >128 | >128 | >128 | >128 | >128 |
| Ampicillin | 0.13 | 0.25 | 0.5 | 128 | 0.28 | 32 |
| Tetracycline | 0.13 | 0.25 | 0.13 | 0.5 | 0.25 | 0.5 |
| IC50 (μM) | SNU638 | SK-HEP-1 | A549 | HCT116 | MDA-MB-23 |
|---|---|---|---|---|---|
| Ligiamycin A (1) | >50 | >50 | 42.0 | >50 | >50 |
| Ligiamycin B (2) | 46.2 | 34.6 | 25.9 | 20.1 | 23.7 |
| Etoposide | 2.17 | 0.24 | 0.35 | 0.66 | 2.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.-J.; An, J.S.; Bae, E.S.; Cho, E.; Hwang, S.; Nam, S.-J.; Oh, K.-B.; Lee, S.K.; Oh, D.-C. Ligiamycins A and B, Decalin-Amino-Maleimides from the Co-Culture of Streptomyces sp. and Achromobacter sp. Isolated from the Marine Wharf Roach, Ligia exotica. Mar. Drugs 2022, 20, 83. https://0-doi-org.brum.beds.ac.uk/10.3390/md20020083
Lim H-J, An JS, Bae ES, Cho E, Hwang S, Nam S-J, Oh K-B, Lee SK, Oh D-C. Ligiamycins A and B, Decalin-Amino-Maleimides from the Co-Culture of Streptomyces sp. and Achromobacter sp. Isolated from the Marine Wharf Roach, Ligia exotica. Marine Drugs. 2022; 20(2):83. https://0-doi-org.brum.beds.ac.uk/10.3390/md20020083
Chicago/Turabian StyleLim, Hyung-Ju, Joon Soo An, Eun Seo Bae, Eunji Cho, Sunghoon Hwang, Sang-Jip Nam, Ki-Bong Oh, Sang Kook Lee, and Dong-Chan Oh. 2022. "Ligiamycins A and B, Decalin-Amino-Maleimides from the Co-Culture of Streptomyces sp. and Achromobacter sp. Isolated from the Marine Wharf Roach, Ligia exotica" Marine Drugs 20, no. 2: 83. https://0-doi-org.brum.beds.ac.uk/10.3390/md20020083