
The Structural Characteristics and Bioactivity Stability of Cucumaria frondosa Intestines and Ovum Hydrolysates Obtained by Different Proteases
 ,
,
Abstract
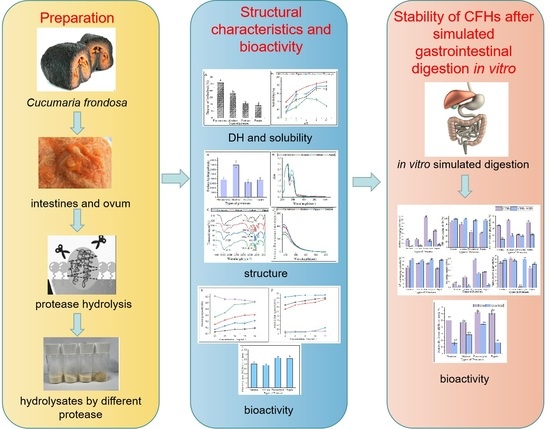
:
1. Introduction
2. Results and Discussion
2.1. Degree of Hydrolysis (DH) of Hydrolysates Obtained by Different Proteases
2.2. Solubility of Hydrolysates Obtained by Different Proteases
2.3. Structural Characteristics of Hydrolysates
2.3.1. Surface Hydrophobicity
2.3.2. Ultraviolet-Visible (UV-Vis) Spectroscopy Analysis
2.3.3. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.3.4. Intrinsic Fluorescence Spectroscopy Analysis
2.4. Antioxidant Activity of Hydrolysates Obtained by Different Proteases
2.5. Effect of Different Proteases on the Pancreatic Lipase Inhibitory Activity of Hydrolysates
2.6. Bioactivity Stability of CFHs after Simulated Gastrointestinal Digestion In Vitro
2.6.1. Antioxidant Activity
2.6.2. Pancreatic Lipase Inhibitory Activity
3. Materials and Methods
3.1. Materials and Reagents
3.2. Preparation of Hydrolysates
3.3. Determination of DH
3.4. Solubility
3.5. Structural Characteristics of CFHs
3.5.1. Surface Hydrophobicity
3.5.2. UV-Vis Spectroscopy
3.5.3. FTIR
3.5.4. Intrinsic Fluorescence Spectroscopy
3.6. Antioxidant Activity of CFHs
3.6.1. DPPH Radical Scavenging Activity
3.6.2. Hydroxyl Radical Scavenging Activity
3.6.3. Superoxide Anion Scavenging Activity
3.6.4. Reduction Capacity
3.6.5. Metal-Chelating Activity
3.6.6. ABTS Radical Scavenging Activity
3.7. Pancreatic Lipase Inhibitory Activity
3.8. Simulated Gastrointestinal Digestion In Vitro of Hydrolysates
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, P.H.; Lu, W.C.; Chan, Y.J.; Ko, W.C.; Jung, C.C.; Huynh, D.T.L.; Ji, Y.X. Extraction and characterization of collagen from sea cucumber (Holothuria cinerascens) and its potential application in moisturizing cosmetics. Aquaculture 2020, 515, 734590. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, R.; Wen, Z. Bioactive compounds and biological functions of sea cucumbers as potential functional foods. J. Funct. Foods 2018, 49, 73–84. [Google Scholar] [CrossRef]
- Jattujan, P.; Chalorak, P.; Siangcham, T.; Sangpairoj, K.; Nobsathian, S.; Poomtong, T.; Sobhon, P.; Meemon, K. Holothuria scabra extracts possess anti-oxidant activity and promote stress resistance and lifespan extension in Caenorhabditis elegans. Exp. Gerontol. 2018, 110, 158–171. [Google Scholar] [CrossRef]
- Li, X.R.; Yang, R.W.; Ju, H.P.; Wang, K.; Lin, S.Y. Identification of dominant spoilage bacteria in sea cucumber protein peptide powders (SCPPs) and methods for controlling the growth of dominant spoilage bacteria by inhibiting hygroscopicity. LWT Food Sci. Technol. 2021, 136, 110355. [Google Scholar] [CrossRef]
- Gong, P.X.; Wang, B.K.; Wu, Y.C.; Li, Q.Y.; Qin, B.W.; Li, H.J. Release of antidiabetic peptides from Stichopus japonicas by simulated gastrointestinal digestion. Food Chem. 2020, 315, 126273. [Google Scholar] [CrossRef]
- Wang, T.T.; Zheng, L.; Wang, S.G.; Zhao, M.M.; Liu, X.L. Anti-diabetic and anti-hyperlipidemic effects of sea cucumber (Cucumaria frondosa) gonad hydrolysates in type II diabetic rats. Food Sci. Hum. Wellness 2022, 11, 1614–1622. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Hamid, A.A.; Saari, N. Purification and characterization of angiotensin converting enzyme-inhibitory peptides derived from Stichopus horrens: Stability study against the ACE and inhibition kinetics. J. Funct. Foods 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Northern sea cucumber (Cucumaria frondosa): A potential candidate for functional food, nutraceutical, and pharmaceutical sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Dave, D.; Shahidi, F. Antioxidant potential and physicochemical properties of protein hydrolysates from body parts of North Atlantic sea cucumber (Cucumaria frondosa). Food Prod. Process. Nutr. 2021, 3, 3. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [Green Version]
- Tripoteau, L.; Bedoux, G.; Gagnon, J.; Bourgougnon, N. In vitro antiviral activities of enzymatic hydrolysates extracted from byproducts of the Atlantic holothurian Cucumaria frondosa. Process Biochem. 2015, 50, 867–875. [Google Scholar] [CrossRef]
- Mamelona, J.; Saint-Louis, R.; Pelletier, E. Proximate composition and nutritional profile of by-products from green urchin and Atlantic sea cucumber processing plants. Int. J. Food Sci. Technol. 2010, 45, 2119–2126. [Google Scholar] [CrossRef]
- Karami, Z.; Butkinaree, C.; Yingchutrakul, Y.; Simanon, N.; Duangmal, K. Comparative study on structural, biological and functional activities of hydrolysates from Adzuki bean (Vigna angularis) and mung bean (Vigna radiata) protein concentrates using Alcalase and Flavourzyme. Food Res. Int. 2022, 161, 111797. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.J.; Li, J.X.; Li, J.W.; Sun, H.; Liu, Y.F. Physicochemical and antioxidative characteristics of black bean protein hydrolysates obtained from different enzymes. Food Hydrocoll. 2019, 97, 105222. [Google Scholar] [CrossRef]
- Zhang, X.D.; Dai, Z.Y.; Zhang, Y.Q.; Dong, Y.; Hu, X.J. Structural characteristics and stability of salmon skin protein hydrolysates obtained with different proteases. LWT Food Sci. Technol. 2022, 153, 112460. [Google Scholar] [CrossRef]
- Sharma, S.; Pradhan, R.; Manickavasagan, A.; Tsopmo, A.; Thimmanagari, M.; Dutta, A. Corn distillers solubles by two-step proteolytic hydrolysis as a new source of plant-based protein hydrolysates with ACE and DPP4 inhibition activities. Food Chem. 2023, 401, 134120. [Google Scholar] [CrossRef] [PubMed]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W.; Wu, H.P. Antioxidative and functional properties of protein hydrolysate from defatted skipjack (Katsuwonous pelamis) roe. Food Chem. 2012, 135, 3039–3048. [Google Scholar] [CrossRef]
- Fadimu, G.J.; Gill, H.; Farahnaky, A.; Truong, T. Improving the enzymolysis efficiency of lupin protein by ultrasound pretreatment: Effect on antihypertensive, antidiabetic and antioxidant activities of the hydrolysates. Food Chem. 2022, 383, 132457. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Dynamic rheological, microstructural and physicochemical properties of blend fish protein recovered from kilka (Clupeonella cultriventris) and silver carp (Hypophthalmichthys molitrix) by the pH-shift process or washing-based technology. Food Chem. 2017, 229, 695–709. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Du, M.; Shen, M.; Wu, T.; Lin, L. Physico-chemical properties, antioxidant activities and angiotensin-I converting enzyme inhibitory of protein hydrolysates from Mung bean (Vigna radiate). Food Chem. 2019, 270, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.F.; Li, Y.Q.; Wang, C.Y.; Liang, Y.; Zhao, X.Z.; He, J.X.; Mo, H.Z. Physicochemical, functional and antioxidant properties of mung bean protein enzymatic hydrolysates. Food Chem. 2022, 393, 133397. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, S.; Li, F.; Du, X.; Kong, B.; Wang, H.; Xia, X. Physicochemical properties and antioxidant activity of polysaccharides obtained from sea cucumber gonads via ultrasound-assisted enzymatic techniques. LWT Food Sci. Technol. 2022, 160, 113307. [Google Scholar] [CrossRef]
- Du, X.; Jing, H.; Wang, L.; Huang, X.; Wang, X.; Wang, H. Characterization of structure, physicochemical properties, and hypoglycemic activity of goat milk whey protein hydrolysate processed with different proteases. LWT Food Sci. Technol. 2022, 159, 113257. [Google Scholar] [CrossRef]
- Wardani, D.W.; Ningrum, A.; Manikharda; Vanidia, N.; Munawaroh, H.S.H.; Susanto, E.; Show, P.-L. In silico and in vitro assessment of yellowfin tuna skin (Thunnus albacares) hydrolysate antioxidation effect. Food Hydrocoll. Health 2023, 3, 100126. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, Y.; Ma, R.; Tang, Y.; Li, Y.; Zhang, S. Structural properties and antioxidant activities of soybean protein hydrolysates produced by Lactobacillus delbrueckii subsp. bulgaricus cell envelope proteinase. Food Chem. 2023, 410, 135392. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Tan, Y.; Hong, H.; Luo, Y. Generation of novel antioxidant peptides from silver carp muscle hydrolysate: Gastrointestinal digestion stability and transepithelial absorption property. Food Chem. 2023, 403, 134136. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Taie, H.A.A.; Abdel Wahab, W.A. Antioxidant capacity and antitumor activity of the bioactive protein prepared from orange peel residues as a by-product using fungal protease. Int. J. Biol. Macromol. 2023, 234, 123578. [Google Scholar] [CrossRef]
- Pv, S. Protein hydrolysate from duck egg white by Flavourzyme® digestion: Process optimisation by model design approach and evaluation of antioxidant capacity and characteristic properties. LWT Food Sci. Technol. 2022, 156, 113018. [Google Scholar] [CrossRef]
- Liu, F.F.; Li, Y.Q.; Sun, G.J.; Wang, C.Y.; Liang, Y.; Zhao, X.Z.; He, J.X.; Mo, H.Z. Influence of ultrasound treatment on the physicochemical and antioxidant properties of mung bean protein hydrolysate. Ultrason. Sonochem. 2022, 84, 105964. [Google Scholar] [CrossRef]
- Mudgil, P.; Kamal, H.; Yuen, G.C.; Maqsood, S. Characterization and identification of novel antidiabetic and anti-obesity peptides from camel milk protein hydrolysates. Food Chem. 2018, 259, 46–54. [Google Scholar] [CrossRef]
- Baba, W.N.; Mudgil, P.; Baby, B.; Vijayan, R.; Gan, C.Y.; Maqsood, S. New insights into the cholesterol esterase- and lipase-inhibiting potential of bioactive peptides from camel whey hydrolysates: Identification, characterization, and molecular interaction. J. Dairy Sci. 2021, 104, 7393–7405. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, D.Y.; Hur, S.J. Changes in the stability and antioxidant activities of different molecular weight bioactive peptide extracts obtained from beef during in vitro human digestion by gut microbiota. Food Res. Int. 2021, 141, 110116. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.F.; Li, B.F.; Hou, H.; Zhang, H.W.; Zhao, X. Identification of iron-chelating peptides from Pacific cod skin gelatin and the possible binding mode. J. Funct. Foods 2017, 35, 418–427. [Google Scholar] [CrossRef]
- Huang, C.; Tang, X.; Liu, Z.; Huang, W.; Ye, Y. Enzymes-dependent antioxidant activity of sweet apricot kernel protein hydrolysates. LWT Food Sci. Technol. 2022, 154, 112825. [Google Scholar] [CrossRef]
- Vásquez, P.; Sepúlveda, C.T.; Zapata, J.E. Functional properties of rainbow trout (Oncorhynchus mykiss) viscera protein hydrolysates. Biocatal. Agric. Biotechnol. 2022, 39, 102268. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.; Farr, A.L.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Zhang, Q.T.; Tu, Z.C.; Xiao, H.; Wang, H.; Huang, X.Q.; Liu, G.X.; Liu, C.M.; Shi, Y.; Fan, L.L.; Lin, D.R. Influence of ultrasonic treatment on the structure and emulsifying properties of peanut protein isolate. Food Bioprod. Process. 2014, 92, 30–37. [Google Scholar] [CrossRef]
- Du, M.X.; Xie, J.H.; Gong, B.; Xu, X.; Tang, W.; Li, X.; Li, C.; Xie, M.Y. Extraction, physicochemical characteristics and functional properties of Mung bean protein. Food Hydrocoll. 2018, 76, 131–140. [Google Scholar] [CrossRef]
- Zhou, C.; Mi, S.; Li, J.; Gao, J.; Wang, X.H.; Sang, Y.X. Purification, characterisation and antioxidant activities of chondroitin sulphate extracted from Raja porosa cartilage. Carbohydr. Polym. 2020, 241, 116306. [Google Scholar] [CrossRef]
- Xie, J.H.; Wang, Z.J.; Shen, M.Y.; Nie, S.P.; Gong, B.; Li, H.S.; Zhao, Q.; Li, W.J.; Xie, M.Y. Sulfated modification, characterization and antioxidant activities of polysaccharide from Cyclocarya paliurus. Food Hydrocoll. 2016, 53, 7–15. [Google Scholar] [CrossRef]
- Liu, J.B.; Jin, Y.; Lin, S.Y.; Jones, G.S.; Chen, F. Purification and identification of novel antioxidant peptides from egg white protein and their antioxidant activities. Food Chem. 2015, 175, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Fisayo Ajayi, F.; Mudgil, P.; Gan, C.Y.; Maqsood, S. Identification and characterization of cholesterol esterase and lipase inhibitory peptides from amaranth protein hydrolysates. Food Chem. X 2021, 12, 100165. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food––An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteases | Temperature (°C) | pH | Solid–Liquid Ratio (w/v) | Proteases Addition (U/g Protein) | Time (h) |
|---|---|---|---|---|---|
| Neutrase | 50 | 7 | 1:20 | 6000 | 7 |
| Alcalase | 60 | 10.5 | 1:20 | 7000 | 7 |
| Flavourzyme | 35 | 5.5 | 1:15 | 10,000 | 9 |
| Papain | 60 | 7.5 | 1:8 | 7000 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Wang, G.; Liu, C.; Sun, Z.; Li, R.; Gao, J.; Li, M.; Sun, L. The Structural Characteristics and Bioactivity Stability of Cucumaria frondosa Intestines and Ovum Hydrolysates Obtained by Different Proteases. Mar. Drugs 2023, 21, 395. https://0-doi-org.brum.beds.ac.uk/10.3390/md21070395
Wang Q, Wang G, Liu C, Sun Z, Li R, Gao J, Li M, Sun L. The Structural Characteristics and Bioactivity Stability of Cucumaria frondosa Intestines and Ovum Hydrolysates Obtained by Different Proteases. Marine Drugs. 2023; 21(7):395. https://0-doi-org.brum.beds.ac.uk/10.3390/md21070395
Chicago/Turabian StyleWang, Qiuting, Gongming Wang, Chuyi Liu, Zuli Sun, Ruimin Li, Jiarun Gao, Mingbo Li, and Leilei Sun. 2023. "The Structural Characteristics and Bioactivity Stability of Cucumaria frondosa Intestines and Ovum Hydrolysates Obtained by Different Proteases" Marine Drugs 21, no. 7: 395. https://0-doi-org.brum.beds.ac.uk/10.3390/md21070395