
Structure and Metabolically Oriented Efficacy of Fucoidan from Brown Alga Sargassum muticum in the Model of Colony Formation of Melanoma and Breast Cancer Cells

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Investigation of the Structure of Fucoidan 2SmF2 from S. muticum by Methylation Analysis and NMR Spectroscopy of Its Desulfated, Deacetylated Derivative
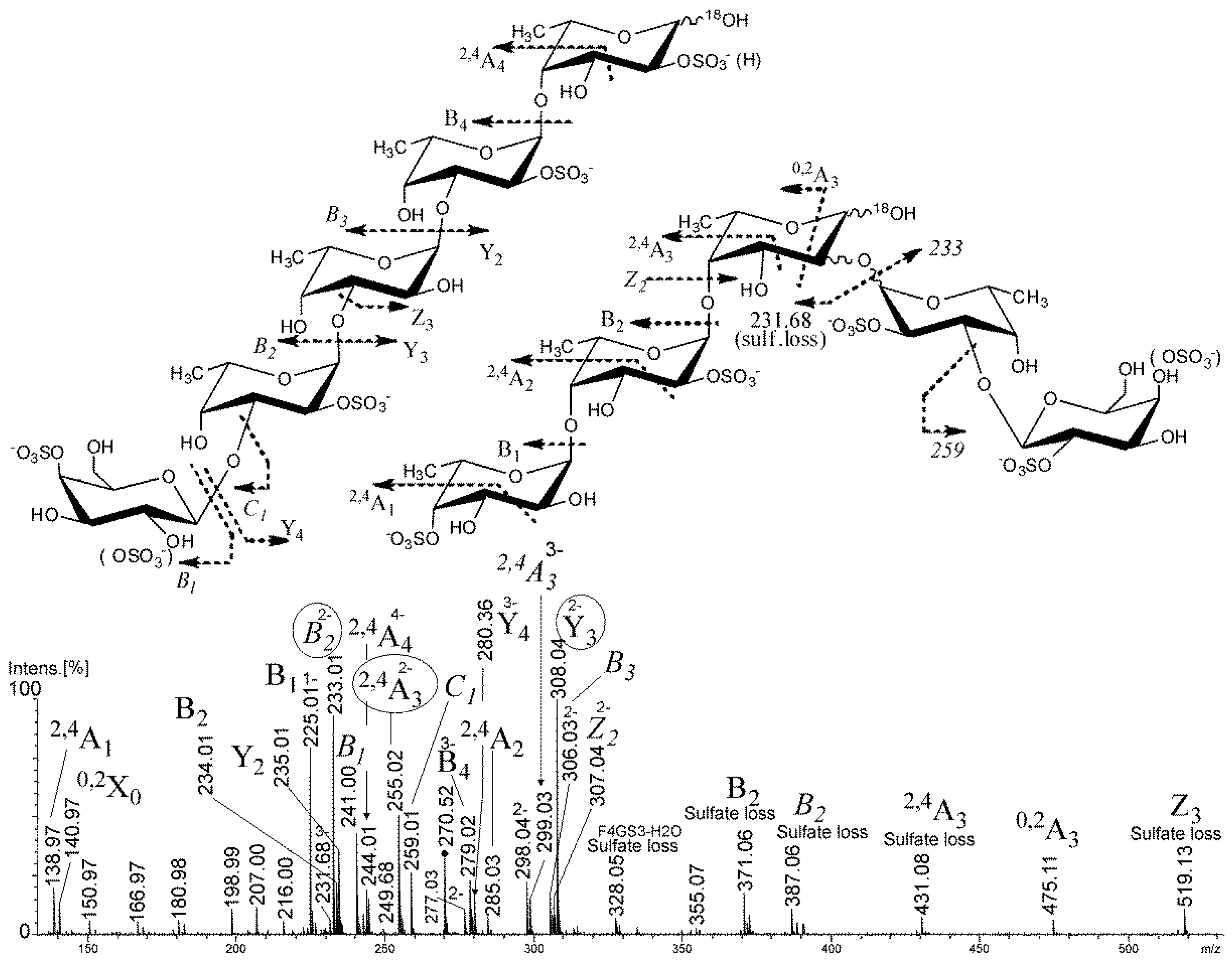
2.2. Mass Spectrometry of the Partial Degradation Products
2.2.1. Mass Spectrometry of Oligosaccharides, Obtained by Autohydrolysis in Heavy-Oxygen Water
2.2.2. Mass Spectrometry of Oligosaccharides, Obtained by Mild Acid Hydrolysis in Heavy-Oxygen Water
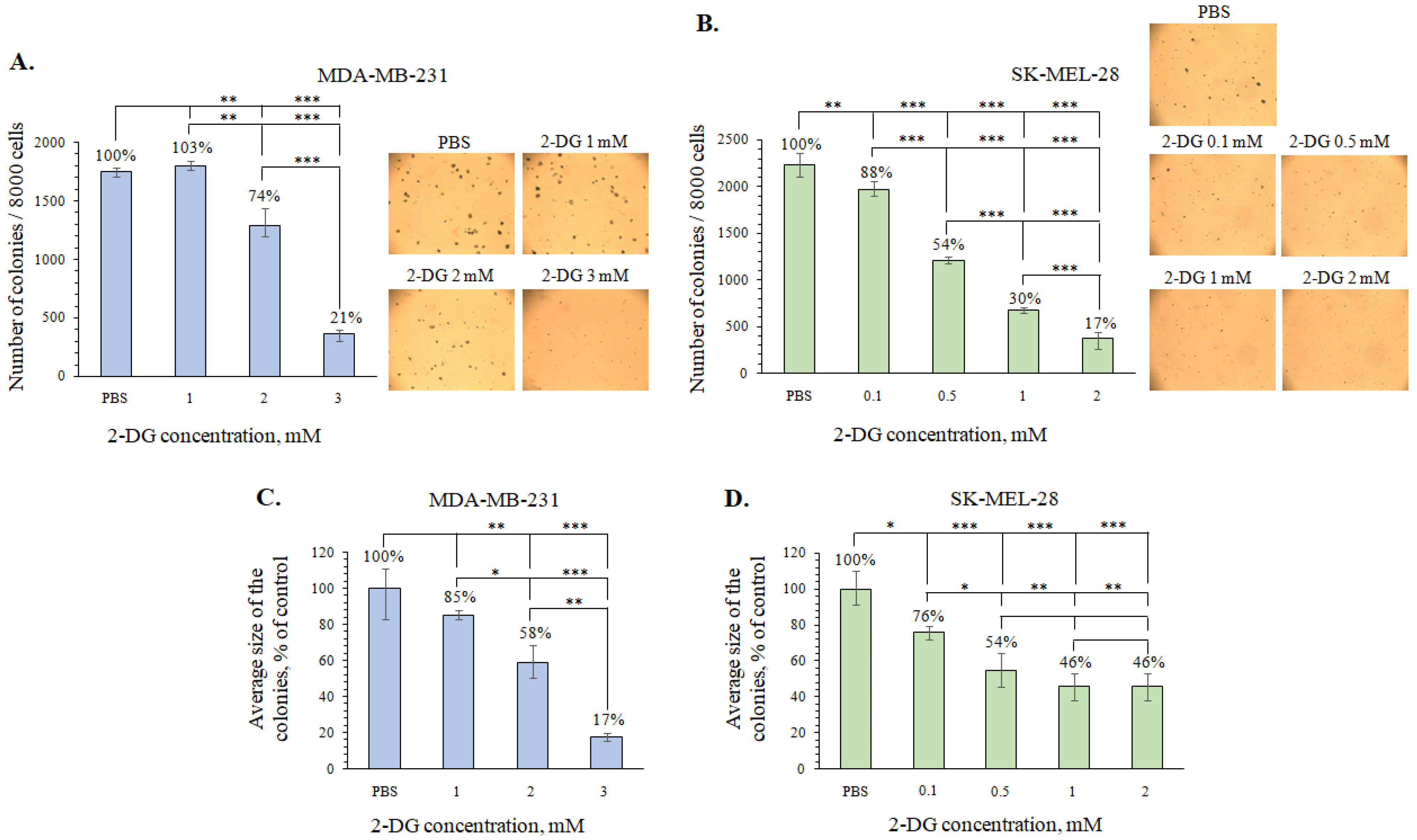
2.3. The Investigation of the Metabolically Oriented Effect of Fucoidan 2SmF2 from S. muticum
3. Materials and Methods
3.1. Materials
3.2. Instruments
3.3. General Methods
3.3.1. Isolation of Fucoidan 2SmF2 from S. muticum
3.3.2. Modification of Fucoidan 2SmF2
3.3.3. Methylation of Desulfated and Deacetylated Fucoidan 2SmF2DADS
3.3.4. Autohydrolysis of Fucoidan 2SmF2
3.3.5. Mild Acid Hydrolysis of Desulfated and Deacetylated Fucoidan 2SmF2DADS
3.4. Anticancer Activity
3.4.1. Cell Culturing
3.4.2. Glucose Uptake Assay
3.4.3. Cell Cytotoxicity Assay
3.4.4. Soft Agar Cologenic Assay
3.4.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicolini, A.; Carpi, A.; Ferrari, P.; Biava, P.M.; Rossi, G. Immunotherapy and hormone-therapy in metastatic breast cancer: A review and an update. Curr. Drug Targets 2016, 17, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting cancer metabolism in the era of precision oncology. Nat. Rev. Drug Discov. 2022, 21, 141–162. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, O.; Navon, G.; Lyon, R.C.; Faustino, P.J.; Straka, E.J.; Cohen, J.S. Effects of 2-deoxyglucose on drug-sensitive and drug-resistant human breast cancer cells: Toxicity and magnetic resonance spectroscopy studies of metabolism. Cancer Res. 1990, 50, 544–551. [Google Scholar]
- Haga, N.; Naito, M.; Seimiya, H.; Tomida, A.; Dong, J.; Tsuruo, T. 2-Deoxyglucose inhibits chemotherapeutic drug-induced apoptosis in human monocytic leukemia U937 cells with inhibition of c-Jun N-terminal kinase 1/stress-activated protein kinase activation. Int. J. Cancer 1998, 76, 86–90. [Google Scholar] [CrossRef]
- Landau, B.R.; Laszlo, J.; Stengle, J.; Burk, D. Certain metabolic and pharmacologic effects in cancer patients given infusions of 2-deoxy-d-glucose. J. Natl. Cancer Inst. 1958, 21, 485–494. [Google Scholar]
- Gill, K.S.; Fernandes, P.; O’Donovan, T.R.; McKenna, S.L.; Doddakula, K.K.; Power, D.G.; Soden, D.M.; Forde, P.F. Glycolysis inhibition as a cancer treatment and its role in an anti-tumour immune response. Biochim. Biophys. Acta 2016, 1866, 87–105. [Google Scholar] [CrossRef]
- Jin, J.O.; Yadav, D.; Madhwani, K.; Puranik, N.; Chavda, V.; Song, M. Seaweeds in the oncology arena: Anti-cancer potential of fucoidan as a drug—A review. Molecules 2022, 27, 6032. [Google Scholar] [CrossRef]
- Abdollah, M.R.A.; Ali, A.A.; Elgohary, H.H.; Elmazar, M.M. Antiangiogenic drugs in combination with seaweed fucoidan: A mechanistic in vitro and in vivo study exploring the VEGF receptor and its downstream signaling molecules in hepatic cancer. Front. Pharmacol. 2023, 14, 1108992. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Malyarenko, T.V.; Usoltseva, R.V.; Silchenko, A.S.; Kicha, A.A.; Ivanchina, N.V.; Ermakova, S.P. Fucoidan from brown algae Fucus evanescens potentiates the anti-proliferative efficacy of asterosaponins from starfish Asteropsis carinifera in 2D and 3D models of melanoma cells. Int. J. Biol. Macromol. 2021, 185, 31–39. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Malyarenko, T.V.; Usoltseva, R.V.; Kicha, A.A.; Ivanchina, N.V.; Ermakova, S.P. Combined radiomodifying effect of fucoidan from the brown alga Saccharina cichorioides and pacificusoside D from the starfish Solaster pacificus in the model of 3D melanoma cells. Biomolecules 2023, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Burney, M.; Mathew, L.; Gaikwad, A.; Nugent, E.K.; Gonzalez, A.O.; Smith, J.A. Evaluation fucoidan extracts from Undaria pinnatifida and Fucus vesiculosus in combination with anticancer drugs in human cancer orthotopic mouse models. Integr. Cancer Ther. 2018, 17, 755–761. [Google Scholar] [CrossRef]
- Abudabbus, A.; Badmus, J.A.; Shalaweh, S.; Bauer, R.; Hiss, D. Effects of fucoidan and chemotherapeutic agent combinations on malignant and non-malignant breast cell lines. Curr. Pharm. Biotechnol. 2017, 18, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Teruya, K.; Yoshida, T.; Eto, H.; Shirahata, S. Fucoidan extract enhances the anti-cancer activity of chemotherapeutic agents in MDA-MB-231 and MCF-7 breast cancer cells. Mar. Drugs 2013, 11, 81–98. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Usoltseva, R.V.; Shevchenko, N.M.; Surits, V.V.; Imbs, T.I.; Malyarenko, O.S.; Besednova, N.N.; Ivanushko, L.A.; Ermakova, S.P. Structural diversity of fucoidans and their radioprotective effect. Carbohydr. Polym. 2021, 273, 118551. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Malyarenko, O.S.; Anastyuk, S.D.; Shevchenko, N.M.; Silchenko, A.S.; Zvyagintseva, T.N.; Isakov, V.V.; Thinh, P.D.; Khanh, H.H.N.; Hang, C.T.T.; et al. The structure of fucoidan from Sargassum oligocystum and radiosensitizing activity of galactofucans from some algae of genus Sargassum. Int. J. Biol. Macromol. 2021, 183, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Thinh, P.D.; Menshova, R.V.; Ermakova, S.P.; Anastyuk, S.D.; Ly, B.M.; Zvyagintseva, T.N. Structural characteristics and anticancer activity of fucoidan from the brown alga Sargassum mcclurei. Mar. Drugs 2013, 11, 1456–1476. [Google Scholar] [CrossRef]
- Herawati, D.; Hendradi, E.; Zaidan, A.H.; Pudjiastuti, P. Microwave-assisted extraction of fucoidan from Sargassum plagiophyllum and its activities. Pak. J. Biol. Sci. 2022, 25, 1008–1013. [Google Scholar]
- Ji, Y.-B.; Ji, C.-F.; Yue, L. Human gastric cancer cell line SGC-7901 apoptosis induced by SFPS-B2 via a mitochondrial-mediated pathway. Bio-Med. Mater. Eng. 2014, 24, 1141–1147. [Google Scholar] [CrossRef]
- Sung, C.-J.; Wang, H.-H.; Sun, K.-H.; Hsieh, C.-C.; Huang, R.; Sun, G.-H.; Tang, S.-J. Fucoidan from Sargassum hemiphyllum inhibits the stemness of cancer stem cells and epithelial-mesenchymal transitions in bladder cancer cells. Int. J. Biol. Macromol. 2022, 221, 623–633. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Manikandakrishnan, M.; Anjali, R.; Rajasekar, P.; Marudhupandi, T.; Manikandan, R.; Vaseeharan, B.; Prabhu, N.M. Investigation of antioxidant and anticancer potential of fucoidan from Sargassum polycystum. Int. J. Biol. Macromol. 2018, 116, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Usol’tseva, R.V.; Zhao, P.; Kusaikin, M.I.; Jia, A.; Yuan, W.; Zhang, M.; Liu, C.; Ermakova, S.P. Structural characteristics and antitumor activity of fucoidans from the brown alga Sargassum muticum. Chem. Nat. Compd. 2017, 53, 219–223. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Kelly, M.; Sanderson, C.J.; Nifantiev, N.E.; Usov, A.I. Further studies on the composition and structure of a fucoidan preparation from the brown alga Saccharina latissima. Carbohydr. Res. 2010, 345, 2038–2047. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Thuy, T.T.; Van, T.T.; Ly, B.M.; Nifantiev, N.E.; Usov, A.I. Preliminary investigation of a highly sulfated galactofucan fraction isolated from the brown alga Sargassum polycystum. Carbohydr. Res. 2013, 377, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Björndal, H.; Hellerqvist, C.G.; Lindberg, B.; Svensson, S. Gas-liquid chromatography and mass spectrometry in methylation analysis of polysaccharides. Angew. Chem. Int. Ed. 1970, 9, 610–619. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Shevchenko, N.M.; Surits, V.V.; Silchenko, A.S.; Isakov, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Ermakova, S.P. Polysaccharides from brown algae Sargassum duplicatum: The structure and anticancer activity in vitro. Carbohydr. Polym. 2017, 175, 547–556. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Surits, V.V.; Shevchenko, N.M.; Thinh, P.D.; Zadorozhny, P.A.; Ermakova, S.P. Comparison of structure and in vitro anticancer activity of native and modified fucoidans from Sargassum feldmannii and S. duplicatum. Int. J. Biol. Macromol. 2019, 124, 220–228. [Google Scholar] [CrossRef]
- Jin, W.; Wu, W.; Tang, H.; Wei, B.; Wang, H.; Sun, J.; Zhang, W.; Zhong, W. Structure analysis and anti-tumor and anti-angiogenic activities of sulfated galactofucan extracted from Sargassum thunbergii. Mar. Drugs 2019, 17, 52. [Google Scholar] [CrossRef]
- Shevchenko, N.M.; Anastyuk, S.D.; Menshova, R.V.; Vishchuk, O.S.; Isakov, V.I.; Zadorozhny, P.A.; Sikorskaya, T.V.; Zvyagintseva, T.N. Further studies on structure of fucoidan from brown alga Saccharina gurjanovae. Carbohydr. Polym. 2015, 121, 207–216. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Shevchenko, N.M.; Zvyagintseva, T.N.; Ermakova, S.P. The comparison of structure and anticancer activity in vitro of polysaccharides from brown algae Alaria marginata and A. angusta. Carbohydr. Polym. 2016, 153, 258–265. [Google Scholar] [CrossRef]
- Tissot, B.; Salpin, J.-Y.; Martinez, M.; Gaigeot, M.-P.; Daniel, R. Differentiation of the fucoidan sulfated L-fucose isomers constituents by CE-ESIMS and molecular modeling. Carbohydr. Res. 2006, 341, 598–609. [Google Scholar] [CrossRef]
- Anastyuk, S.D.; Shevchenko, N.M.; Belokozova, K.V.; Dmitrenok, P.S. Tandem mass spectrometry of fucoidan-derived fragments; labeled with heavy-oxygen. Carbohydr. Res. 2018, 455, 10–13. [Google Scholar] [CrossRef]
- Zueva, A.O.; Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Usoltseva, R.V.; Kalinovsky, A.I.; Kurilenko, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Ermakova, S.P. Expression and biochemical characterization of two recombinant fucoidanases from the marine bacterium Wenyingzhuangia fucanilytica CZ1127T. Int. J. Biol. Macromol. 2020, 164, 3025–3037. [Google Scholar] [CrossRef] [PubMed]
- Daniel, R.; Chevolot, L.; Carrascal, M.; Tissot, B.; Mourao, P.A.S.; Abian, J. Electrospray ionization mass spectrometry of oligosaccharides derived from fucoidan of Ascophyllum nodosum. Carbohydr. Res. 2007, 342, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, S.P.; Menshova, R.V.; Anastyuk, S.D.; Malyarenko, O.S.; Zakharenko, A.M.; Thinh, P.D.; Ly, B.M.; Zvyagintseva, T.N. Structure, chemical and enzymatic modification, and anticancer activity of polysaccharides from the brown alga Turbinaria ornata. J. Appl. Phycolol. 2016, 28, 2495–2505. [Google Scholar] [CrossRef]
- Zhou, D.; Duan, Z.; Li, Z.; Ge, F.; Wei, R.; Kong, L. The significance of glycolysis in tumor progression and its relationship with the tumor microenvironment. Front. Pharmacol. 2022, 13, 1091779. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, X.; Sun, X.; Wang, L.; Chen, S. The glycolytic switch in tumors: How many players are involved? J. Cancer. 2017, 8, 3430–3440. [Google Scholar] [CrossRef]
- Martinez-Outschoorn, U.E.; Peiris-Pagés, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Raez, L.E.; Papadopoulos, K.; Ricart, A.D.; Chiorean, E.G.; Dipaola, R.S.; Stein, M.N.; Rocha Lima, C.M.; Schlesselman, J.J.; Tolba, K.; Langmuir, V.K.; et al. A phase I dose-escalation trial of 2-deoxy-D-glucose alone or combined with docetaxel in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2013, 71, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Wick, A.N.; Drury, D.R.; Nakada, H.I.; Wolfe, J.B. Localization of the primary metabolic block produced by 2-deoxyglucose. J. Biol. Chem. 1957, 224, 963–969. [Google Scholar] [CrossRef]
- Chen, W.; Gueron, M. The inhibition of bovine heart hexokinase by 2-deoxy-d-glucose-6-phosphate: Characterization by 31P NMR and metabolic implications. Biochimie 1992, 74, 867–873. [Google Scholar] [CrossRef]
- Sim, S.-Y.; Shin, Y.-E.; Kim, H.-K. Fucoidan from Undaria pinnatifida has anti-diabetic effects by stimulation of glucose uptake and reduction of basal lipolysis in 3T3-L1 adipocytes. Nutr. Res. 2019, 65, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Wang, X.; Jiang, H.; Cai, C.; Hao, J.; Yu, G. Fucoidan from Ascophyllum nodosum suppresses postprandial hyperglycemia by inhibiting Na+/glucose cotransporter 1 activity. Mar. Drugs 2020, 18, 485. [Google Scholar] [CrossRef] [PubMed]
- Malyarenko, O.S.; Usoltseva, R.V.; Silchenko, A.S.; Zueva, A.O.; Ermakova, S.P. The combined metabolically oriented effect of fucoidan from the brown alga Saccharina cichorioides and its carboxymethylated derivative with 2-deoxy-d-glucose on human melanoma cells. Int. J. Mol. Sci. 2023, 24, 12050. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef] [PubMed]
- Chiucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Larson, M.G. Analysis of variance. Circulation 2008, 117, 115–121. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Partially Methylated Fucitol or Galactitol Acetates | mol% | Linkage Type |
|---|---|---|
| 2,3,4-tri-O-methyl-fucitol | 5 | Fuc1→ |
| 2,3-di-O-methyl-fucitol | 26 | →4Fuc1→ |
| 2,4-di-O-methyl-fucitol | 20 | →3Fuc1→ |
| 2-O-methyl-fucitol | 15 | →3,4Fuc1→ |
| 3-O-methyl-fucitol | 11 | →2,4Fuc1→ |
| 2,3,4,6-tetra-O-methyl-galactitol | 3 | Gal1→ |
| 2,3,6-tri-O-methyl-galactitol | 13 | →4Gal1→ |
| 2,4,6-tri-O-methyl-galactitol | 1 | →3Gal1→ |
| 2,3,4-tri-O-methyl-galactitol | 2 | →6Gal1→ |
| 3,4-di-O-methyl-galactitol | 1 | →2,6Gal1→ |
| 2,3-di-O-methyl-galactitol | 2 | →4,6Gal1→ |
| 2,4-di-O-methyl-galactitol | 1 | →3,6Gal1→ |
| Residue | C1/H1 | C2/H2 | C3/H3 | C4/H4 | C5/H5 | C6/H6 |
|---|---|---|---|---|---|---|
| A →4)-α-l-Fucp-(1→ | 101.7/5.25 | 70.0/3.82 | 69.8/4.04 | 81.4/3.85 | 69.4/4.29 | 16.9/1.32 |
| B →3)-α-l-Fucp-(1→ | 101.8/4.97 | 69.3/3.97 | 79/3.98 | 73/4.0 | 68.4/4.48 | 16.9/1.32 |
| C →4)-α-d-Galp-(1→ or α-d-Galp-4OSO3−-(1→ | 101.7/5.26 | 70.0/3.82 | 70.5/3.97 | 78.8/4.71 | 72/4.17 | 62.4/3.77, 3.72 |
| m/z | Composition | m/z | Composition |
|---|---|---|---|
| 231.01 | [Fuc3(SO3Na)3 − 3Na]3− | 306.03 | [Fuc2Gal(SO3Na)2 − 2Na − H2O]2− |
| 231.68 | [Fuc3(SO3Na)3 − 3Na − O + 18O]3− | 308.04 | [Fuc3(SO3Na)2 − 2Na − O + 18O]2− |
| 233.01 | [FucGal(SO3Na)2 − 2Na − H2O]2− | 316.04 | [Fuc2Gal(SO3Na)2 − 2Na − O + 18O]2− |
| 235.01 | [Fuc2(SO3Na)2 − 2Na − O + 18O]2− | 327.71 | [Fuc4Gal(SO3Na)3 − 3Na − H2O]3− |
| 237.01 | [Fuc2Gal(SO3Na)3 − 3Na − O + 18O]3− | 334.38 | [Fuc4Gal(SO3Na)3 − 3Na − O + 18O]3− |
| 243.00 | [FucSO3Na − Na)]− | 389.07 | [Fuc2SO3Na − Na]− |
| 245.00 | [FucSO3Na − Na − O + 18O]− | 391.08 | [Fuc2SO3Na − O + 18O]− |
| 270.52 | [Fuc4Gal(SO3Na)4 − 4Na]4− | 535.31 | [Fuc3SO3Na − Na]− |
| 280.36 | [Fuc4(SO3Na)3 − 3Na − O + 18O]3− | 537.13 | [Fuc3SO3Na − Na − O + 18O]− |
| 285.70 | [Fuc3Gal(SO3Na)3 − 3Na]3− |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usoltseva, R.V.; Zueva, A.O.; Malyarenko, O.S.; Anastyuk, S.D.; Moiseenko, O.P.; Isakov, V.V.; Kusaykin, M.I.; Jia, A.; Ermakova, S.P. Structure and Metabolically Oriented Efficacy of Fucoidan from Brown Alga Sargassum muticum in the Model of Colony Formation of Melanoma and Breast Cancer Cells. Mar. Drugs 2023, 21, 486. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090486
Usoltseva RV, Zueva AO, Malyarenko OS, Anastyuk SD, Moiseenko OP, Isakov VV, Kusaykin MI, Jia A, Ermakova SP. Structure and Metabolically Oriented Efficacy of Fucoidan from Brown Alga Sargassum muticum in the Model of Colony Formation of Melanoma and Breast Cancer Cells. Marine Drugs. 2023; 21(9):486. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090486
Chicago/Turabian StyleUsoltseva, Roza V., Anastasiya O. Zueva, Olesya S. Malyarenko, Stanislav D. Anastyuk, Olga P. Moiseenko, Vladimir V. Isakov, Mikhail I. Kusaykin, Airong Jia, and Svetlana P. Ermakova. 2023. "Structure and Metabolically Oriented Efficacy of Fucoidan from Brown Alga Sargassum muticum in the Model of Colony Formation of Melanoma and Breast Cancer Cells" Marine Drugs 21, no. 9: 486. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090486