
A Chemo-Ecological Investigation of Dendrilla antarctica Topsent, 1905: Identification of Deceptionin and the Effects of Heat Stress and Predation Pressure on Its Terpene Profiles
 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Major Diterpenes in D. antarctica Populations from Two Different Localities
2.2. Structural Characterization of Deceptionin (7)
2.3. Terpene Profile of Sponges in Their Natural Habitat
2.4. Chemotyping
2.5. Effect of Experimental Heat Stress on the Secondary Metabolites of D. antarctica
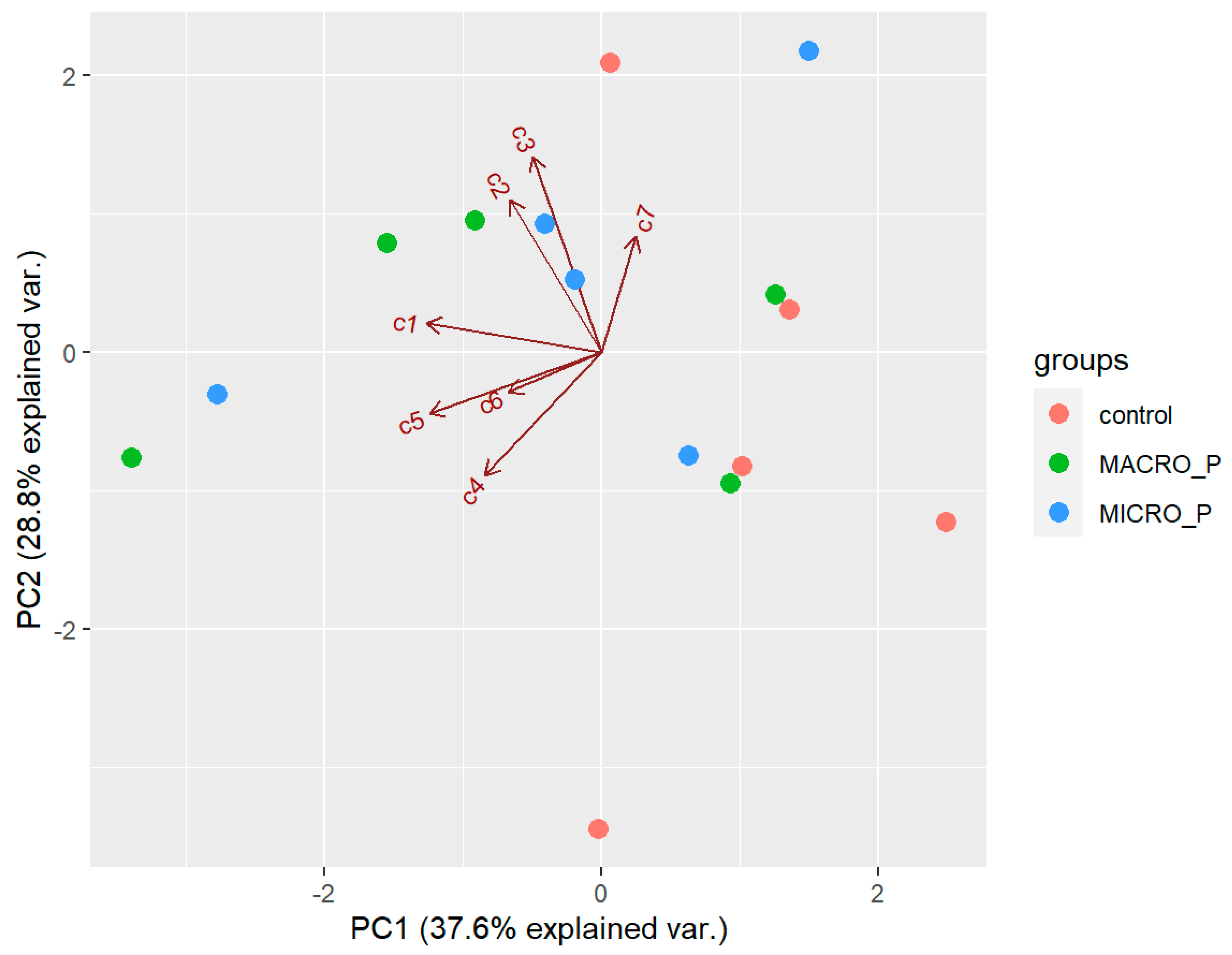
2.6. Effect of Experimental Predation Pressure on the Secondary Metabolites of D. antarctica
2.7. Antimicrobial Assays
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Heat Stress and Predation Pressure Experiments in Aquaria
4.3. General Analysis
4.4. Metabolomic Analysis
4.5. Isolation and Characterization of Diterpenes from D. antarctica
4.6. Deceptionin (7)
4.7. Data Analysis
4.8. Antimicrobial Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dayton, P.; Robilliard, G.A.; Paine, R.T. Benthic Faunal Zonation as a Result of Anchor Ice at McMurdo Sound, Antarctica. In Antarctic Ecology; Holdgate, M.W., Ed.; Academic Press: New York, NY, USA, 1970; Volume 1, pp. 244–258. [Google Scholar]
- Janussen, D.; Downey, R.V. Porifera. In Biogeographic Atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H., Raymond, B., d’Udekem d’Acoz, C., Van de Putte, A., Danis, B., David, B., Grant, S., Gutt, J., et al., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 94–102. [Google Scholar]
- Cárdenas, C.A.; Montiel, A. Coexistence in Cold Waters: Animal Forests in Seaweed-Dominated Habitats in Southern High Latitudes. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 257–276. [Google Scholar] [CrossRef]
- Cárdenas, C.A.; González-Aravena, M.; Font, A.; Hestetun, J.T.; Hajdu, E.; Trefault, N.; Malmberg, M.; Bongcam-Rudloff, E. High Similarity in the Microbiota of Cold- Water Sponges of the Genus Mycale from Two Different Geographical Areas. PeerJ 2018, 6, e4935. [Google Scholar] [CrossRef]
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Lebar, M.D.; Heimbegner, J.L.; Baker, B.J.; Baker, B. Cold-Water Marine Natural Products. Nat. Prod. Rep. 2007, 24, 774–797. [Google Scholar] [CrossRef]
- Angulo-Preckler, C.; Castro-Fernandez, P.; Martín-Martín, R.; Figuerola, B.; Avila, C. Chemical ecology in the Southern Ocean. In Life in Extreme Environments: Insights in Biological Capability; Di Prisco, G., Edwards, H., Elster, J., Huiskes, A., Eds.; Cambridge University Press: Cambridge, UK, 2020; pp. 251–278. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Avila, C. Natural Products Mediating Ecological Interactions in Antarctic Benthic Communities: A Mini-Review of the Known Molecules. Nat. Prod. Rep. 2015, 32, 1114–1130. [Google Scholar] [CrossRef]
- Soldatou, S.; Baker, B.J. Cold-Water Marine Natural Products, 2006 to 2016. Nat. Prod. Rep. 2017, 34, 585–626. [Google Scholar] [CrossRef]
- Figuerola, B.; Núñez-Pons, L.; Moles, J.; Avila, C. Feeding Repellence in Antarctic Bryozoans. Naturwissenschaften 2013, 100, 1069–1081. [Google Scholar] [CrossRef]
- Moles, J.; Núñez, L.; Taboada, S.; Figuerola, B. Anti-Predatory Chemical Defences in Antarctic Benthic Fauna. Mar. Biol. 2015, 162, 1813–1821. [Google Scholar] [CrossRef]
- Angulo-Preckler, C.; San Miguel, O.; García-Aljaro, C.; Avila, C. Antibacterial Defenses and Palatability of Shallow-Water Antarctic Sponges. Hydrobiologia 2018, 806, 123–138. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Sneed, J.M.; Ritson-Williams, R.; Young, R. Marine Chemical Ecology in Benthic Environments. Nat. Prod. Rep. 2019, 36, 410–429. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef]
- Goodwin, C.E.; Berman, J.; Hendry, K.R. Demosponges from the Sublittoral and Shallow-Circalittoral (<24 m Depth) Antarctic Peninsula with a Description of Four New Species and Notes on in situ Identification Characteristics. Zootaxa 2019, 4658, 461–508. [Google Scholar] [CrossRef]
- Koltun, V.M. Porifera-Part 1: Antarctic Sponges. Rep. B.A.N.Z. Antarct. Res. Exped. 1929-1931 B (Zool. Bot.) 1976, 5, 153–198. [Google Scholar]
- Brueggeman, P. Underwater Field Guide to Ross Island & McMurdo Sound, Antarctica, Volume 4: Echinodermata, 1998. Available online: https://www.peterbrueggeman.com/nsf/fguide/ (accessed on 30 July 2023).
- von Salm, J.L.; Witowski, C.G.; Amsler, M.O.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Amphipod Diversity and Metabolomics of the Antarctic Sponge Dendrilla antarctica. Antarct. Sci. 2022, 34, 349–360. [Google Scholar] [CrossRef]
- Molinski, T.F.; Faulkner, D.J. Metabolites of the Antarctic Sponge Dendrilla membranosa. J. Org. Chem. 1987, 52, 296–298. [Google Scholar] [CrossRef]
- Fontana, A.; Scognamiglio, G.; Cimino, G. Dendrinolide, a New Degraded Diterpenoid from the Antarctic Sponge Dendrilla membranosa. J. Nat. Prod. 1997, 60, 475–477. [Google Scholar] [CrossRef]
- Ankisetty, S.; Nandiraju, S.; Win, H.; Park, Y.C.; Amsler, C.D.; McClintock, J.B.; Baker, J.A.; Diyabalanage, T.K.; Pasaribu, A.; Singh, M.P.; et al. Chemical Investigation of Predator-Deterred Macroalgae from the Antarctic. J. Nat. Prod. 2004, 67, 1295–1302. [Google Scholar] [CrossRef]
- Prieto, I.M.; Paola, A.; Pérez, M.; García, M.; Blustein, G.; Schejter, L.; Palermo, J.A. Antifouling Diterpenoids from the Sponge Dendrilla antarctica. Chem. Biodivers. 2022, 19, e202100618. [Google Scholar] [CrossRef]
- Baker, B.J.; Yoshida, W.Y.; McClintock, J.B.; Kopitzke, R.W. Chemical and Ecological Studies of the Antarctic Sponge Dendrilla membranosa. J. Nat. Prod. 1995, 58, 1459–1462. [Google Scholar] [CrossRef]
- Bory, A.; Shilling, A.J.; Allen, J.; Azhari, A.; Roth, A.; Shaw, L.N.; Kyle, D.E.; Adams, J.H.; Amsler, C.D.; McClintock, J.B.; et al. Bioactivity of Spongian Diterpenoid Scaffolds from the Antarctic Sponge Dendrilla antarctica. Mar. Drugs 2020, 18, 327. [Google Scholar] [CrossRef]
- von Salm, J.L.; Witowski, C.G.; Fleeman, R.M.; McClintock, J.B.; Amsler, C.D.; Shaw, L.N.; Baker, B.J. Darwinolide, a New Diterpene Scaffold That Inhibits Methicillin-Resistant Staphylococcus aureus Biofilm from the Antarctic Sponge Dendrilla membranosa. Org. Lett. 2016, 18, 2596–2599. [Google Scholar] [CrossRef]
- Amsler, C.D.; Iken, K.; McClintock, B.; Baker, B.J. Defenses of Polar Macroalgae against Herbivores and Biofoulers. Bot. Mar. 2009, 52, 535–545. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Bullough, L.W. Some Observations on the Diet and Distribution of Nudibranchs at Signy Island, Antarctica. J. Mollus. Stud. 1996, 62, 281–287. [Google Scholar] [CrossRef]
- Avila, C. Biological and Chemical Diversity in Antarctica: From New Species to New Natural Products. Biodiversity 2016, 17, 5–11. [Google Scholar] [CrossRef]
- Peters, K.J.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Potential Chemical Defenses of Antarctic Sponges against Sympatric Microorganisms. Polar Biol. 2010, 33, 649–658. [Google Scholar] [CrossRef]
- Peters, K.J.; Amsler, C.D.; McClintock, J.B.; Van Soest, R.W.M.; Baker, B.J. Palatability and Chemical Defenses of Sponges from the Western Antarctic Peninsula. Mar. Ecol. Prog. Ser. 2009, 385, 77–85. [Google Scholar] [CrossRef]
- Cronin, G.; Hay, M.E. Induction of Seaweed Chemical Defenses by Amphipod Grazing. Ecology 1996, 77, 2287–2301. [Google Scholar] [CrossRef]
- Ankisetty, S.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Further Membranolide Diterpenes from the Antarctic Sponge Dendrilla membranosa. J. Nat. Prod. 2004, 67, 1172–1174. [Google Scholar] [CrossRef]
- Shilling, A.J.; Witowski, C.G.; Maschek, J.A.; Azhari, A.; Vesely, B.A.; Kyle, D.E.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Spongian Diterpenoids Derived from the Antarctic Sponge Dendrilla antarctica Are Potent Inhibitors of the Leishmania Parasite. J. Nat. Prod. 2020, 83, 1553–1562. [Google Scholar] [CrossRef]
- Ottaviani, A.; Welsch, J.; Agama, K.; Pommier, Y.; Desideri, A.; Baker, B.J.; Fiorani, P. From Antarctica to Cancer Research: A Novel Human DNA Topoisomerase 1B Inhibitor from Antarctic Sponge Dendrilla antarctica. J. Enzym. Inhib. Med. Chem. 2022, 37, 1404–1410. [Google Scholar] [CrossRef]
- Ciaglia, E.; Malfitano, A.M.; Laezza, C.; Fontana, A.; Nuzzo, G.; Cutignano, A.; Abate, M.; Pelin, M.; Sosa, S.; Bifulco, M.; et al. Immuno-Modulatory and Anti-Inflammatory Effects of Dihydrogracilin A, a Terpene Derived from the Marine Sponge Dendrilla membranosa. Int. J. Mol. Sci. 2017, 18, 1643. [Google Scholar] [CrossRef]
- Molinski, T.F.; Faulkner, D.J. An Antibacterial Pigment from the Sponge Dendrilla membranosa. Tetrahedron Lett. 1988, 29, 2137–2138. [Google Scholar] [CrossRef]
- Pankey, M.S.; Plachetzki, D.C.; Macartney, K.J.; Gastaldi, M.; Slattery, M.; Gochfeld, D.J.; Lesser, M.P. Cophylogeny and convergence shape holobiont evolution in sponge–microbe symbioses. Nat. Ecol. Evol. 2022, 6, 750–762. [Google Scholar] [CrossRef]
- Posadas, N.; Baquiran, J.I.P.; Nada, M.A.L.; Kelly, M.; Conaco, C. Microbiome Diversity and Host Immune Functions Influence Survivorship of Sponge Holobionts under Future Ocean Conditions. ISME J. 2021, 16, 58–67. [Google Scholar] [CrossRef]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The Sponge Holobiont in a Changing Ocean: From Microbes to Ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Björk, J.R.; Easson, C.; Astudillo-García, C.; Olson, J.B.; Erwin, P.M.; López-Legentil, S.; Luter, H.; et al. Diversity, Structure and Convergent Evolution of the Global Sponge Microbiome. Nat. Commun. 2016, 7, 11870. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Azzaro, M.; Schiaparelli, S. Microbial Symbionts of Antarctic Marine Benthic Invertebrates. In The Ecological Role of Micro-Organisms in the Antarctic Environment; Springer Polar Sciences; Castro-Sowinski, S., Ed.; Springer: Cham, Switzerland, 2019; pp. 277–296. [Google Scholar]
- Sacristán-Soriano, O.; Pérez Criado, N.; Avila, C. Host Species Determines Symbiotic Community Composition in Antarctic Sponges (Porifera: Demospongiae). Front. Mar. Sci. 2020, 7, 474. [Google Scholar] [CrossRef]
- Happel, L.; Rondon, R.; Font, A.; González-Aravena, M.; Cárdenas, C.A. Stability of the Microbiome of the Sponge Mycale (Oxymycale) acerata in the Western Antarctic Peninsula. Front. Microbiol. 2022, 13, 827863. [Google Scholar] [CrossRef]
- Murray, A.E.; Avalon, N.E.; Bishop, L.; Davenport, K.W.; Delage, E.; Dichosa, A.E.K.; Eveillard, D.; Higham, M.L.; Kokkaliari, S.; Lo, C.C.; et al. Uncovering the Core Microbiome and Distribution of Palmerolide in Synoicum adareanum Across the Anvers Island Archipelago, Antarctica. Mar. Drugs 2020, 18, 298. [Google Scholar] [CrossRef]
- Easson, C.G.; Chaves-Fonnegra, A.; Thacker, R.W.; Lopez, J.V. Host Population Genetics and Biogeography Structure the Microbiome of the Sponge Cliona delitrix. Ecol. Evol. 2020, 10, 2007–2020. [Google Scholar] [CrossRef]
- Freeman, C.J.; Easson, C.G.; Fiore, C.L.; Thacker, R.W. Sponge–Microbe Interactions on Coral Reefs: Multiple Evolutionary Solutions to a Complex Environment. Front. Mar. Sci. 2021, 8, 917. [Google Scholar] [CrossRef]
- De Castro-Fernández, P.; Ballesté, E.; Angulo-Preckler, C.; Biggs, J.; Avila, C.; García-Aljaro, C. How Does Heat Stress Affect Sponge Microbiomes? Structure and Resilience of Microbial Communities of Marine Sponges from Different Habitats. Front. Mar. Sci. 2023, 9, 1072696. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; Dorta, E.; Cueto, M.; San-Martín, A.; Darias, J. Conformational Analysis and Absolute Stereochemistry of ‘Spongian’-Related Metabolites. Tetrahedron 2004, 60, 1073–1078. [Google Scholar] [CrossRef]
- Mayol, L.; Piccialli, V.; Sica, D. New Degraded Diterpenes from the Sponge Spongionella gracilis. Gazz. Chim. Ital. 1988, 118, 559–563. [Google Scholar]
- Karuso, P.; Skelton, B.W.; Taylor, W.C.; White, A.H. The Constituents of Marine Sponges. I the Isolation from Aplysilla sulphurea (Dendroceratida) of (1r*, 1′s*, 1″r*, 3r*)-1-Acetoxy-4-Ethyl-5-(1, 3, 3-Trimethylcyclohexyl)-1, 3-Dihydroisobenzofuran-L′(4), 3-Carbolactone and the Determination of Its Crystal Structure. Aust. J. Chem. 1984, 37, 1081–1093. [Google Scholar] [CrossRef]
- Karuso, P.; Bergquist, P.R.; Cambie, R.C.; Buckleton, J.S.; Clark, G.R.; Rickard, C.E.F. Terpenoid Constituents of Morphologically Similar Sponges in the Family Aplysillidae. Aust. J. Chem. 1986, 39, 1643–1653. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Northcote, P.T.; Zubkov, O.A. Novel Anti-Inflammatory Spongian Diterpenes from the New Zealand Marine Sponge Chelonaplysilla violacea. Eur. J. Org. Chem. 2004, 2004, 419–425. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Northcote, P.T.; Davies-Coleman, M.T. Spongian diterpenoids from marine sponges. Nat. Prod. Rep. 2006, 23, 321–334. [Google Scholar] [CrossRef]
- Mahon, A.R.; Amsler, C.D.; McClintock, J.B.; Amsler, M.O.; Baker, B.J. Tissue-Specific Palatability and Chemical Defenses against Macropredators and Pathogens in the Common Articulate Brachiopod Liothyrella uva from the Antarctic Peninsula. J. Exp. Mar. Biol. Ecol. 2003, 290, 197–210. [Google Scholar] [CrossRef]
- Puyana, M.; Fenical, W.; Pawlik, J.R. Are There Activated Chemical Defenses in Sponges of the Genus Aplysina from the Caribbean? Mar. Ecol. Prog. Ser. 2003, 246, 127–135. [Google Scholar] [CrossRef]
- Rohde, S.; Gochfeld, D.J.; Ankisetty, S.; Avula, B.; Schupp, P.J.; Slattery, M. Spatial Variability in Secondary Metabolites of the Indo-Pacific Sponge Stylissa massa. J. Chem. Ecol. 2012, 38, 463–475. [Google Scholar] [CrossRef]
- Cutignano, A.; Zhang, W.; Avila, C.; Cimino, G.; Fontana, A. Intrapopulation Variability in the Terpene Metabolism of the Antarctic Opisthobranch Mollusc Austrodoris kerguelenensis. Eur. J. Org. Chem. 2011, 2011, 5383–5389. [Google Scholar] [CrossRef]
- Ortiz, R.; Vila, J.; García, A.; Camacho, A.G.; Diez, J.L.; Aparicio, A.; Petrinovic, I. Geophysical Features of Deception Island. In Recent Progress in Antarctic Earth Science; Yoshida, Y., Kaminuma, K., Shiraishi, K., Eds.; Terra Scientific Publishing Company: Tokyo, Japan, 1992; pp. 443–448. [Google Scholar]
- Smith, K.L.; Baldwin, R.J.; Kaufmann, R.S.; Sturz, A. Ecosystem Studies at Deception Island, Antarctica: An Overview. Deep-Sea Res. Part II-Top. Stud. Oceanogr. 2003, 50, 1595–1609. [Google Scholar] [CrossRef]
- Deheyn, D.D.; Gendreau, P.; Baldwin, R.J.; Latz, M.I. Evidence for Enhanced Bioavailability of Trace Elements in the Marine Ecosystem of Deception Island, a Volcano in Antarctica. Mar. Environ. Res. 2005, 60, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Elderfield, H. Effects of Volcanism on Water Chemistry, Deception Island, Antarctica. Mar. Geol. 1972, 13, M1–M6. [Google Scholar] [CrossRef]
- Shilling, A.J. The Chemical Ecology and Drug Discovery Potential of the Antarctic Red Alga Plocamium cartilagineum and the Antarctic Sponge Dendrilla membranosa. Ph.D. Dissertation, University of South Florida, Tampa, FL, USA, 2019. [Google Scholar]
- Young, R.M.; von Salm, J.L.; Amsler, M.O.; Lopez-Bautista, J.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Site-Specific Variability in the Chemical Diversity of the Antarctic Red Alga Plocamium cartilagineum. Mar. Drugs 2013, 11, 2126–2139. [Google Scholar] [CrossRef]
- Shilling, A.J.; Heiser, S.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Hidden Diversity in an Antarctic Algal Forest: Metabolomic Profiling Linked to Patterns of Genetic Diversification in the Antarctic Red Alga Plocamium sp. Mar. Drugs 2021, 19, 607. [Google Scholar] [CrossRef]
- Wilson, N.G.; Maschek, J.A.; Baker, B.J. A Species Flock Driven by Predation? Secondary Metabolites Support Diversification of Slugs in Antarctica. PLoS ONE 2013, 8, e80277. [Google Scholar] [CrossRef]
- Maroni, P.J.; Baker, B.J.; Moran, A.L.; Woods, H.A.; Avila, C.; Johnstone, G.J.; Stark, J.S.; Kocot, K.M.; Lockhart, S.; Saucède, T.; et al. One Antarctic Slug to Confuse Them All: The Underestimated Diversity of Doris kerguelenensis. Invertebr. Syst. 2022, 36, 419–435. [Google Scholar] [CrossRef]
- Leiva, C.; Taboada, S.; Kenny, N.J.; Combosch, D.; Giribet, G.; Jombart, T.; Riesgo, A. Population Substructure and Signals of Divergent Adaptive Selection despite Admixture in the Sponge Dendrilla antarctica from Shallow Waters Surrounding the Antarctic Peninsula. Mol. Ecol. 2019, 28, 3151–3170. [Google Scholar] [CrossRef]
- Angulo-Preckler, C.; Figuerola, B.; Núñez-Pons, L.; Moles, J.; Martín-Martín, R.; Rull-Lluch, J.; Gómez-Garreta, A.; Avila, C. Macrobenthic Patterns at the Shallow Marine Waters in the Caldera of the Active Volcano of Deception Island, Antarctica. Cont. Shelf Res. 2018, 157, 20–31. [Google Scholar] [CrossRef]
- Angulo-Preckler, C.; Tuya, F.; Avila, C. Abundance and Size Patterns of Echinoderms in Coastal Soft-Bottoms at Deception Island (South Shetland Islands, Antarctica). Cont. Shelf Res. 2017, 137, 131–141. [Google Scholar] [CrossRef]
- McClintock, J.B. Trophic Biology of Antarctic Shallow-Water Echinoderms. Mar. Ecol. Prog. Ser. 1994, 111, 191–202. [Google Scholar] [CrossRef]
- Maschek, J.A. Chemical Investigation of the Antarctic Marine Invertebrates Austrodoris kerguelenensis & Dendrilla membranosa and the Antarctic Red Alga Gigartina skottsbergii. Ph.D. Dissertation, University of South Florida, Tampa, FL, USA, 2011. [Google Scholar]
- Yamada, Y.; Kuzuyama, T.; Komatsu, M.; Shin-ya, K.; Omura, S.; Cane, D.E.; Ikeda, H. Terpene Synthases Are Widely Distributed in Bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 857–862. [Google Scholar] [CrossRef] [PubMed]
- De Broyer, C.; Jazdzewska, A. Biogeographic Patterns of Southern Ocean Benthic Amphipods. In Biogeographic Atlas of The Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H., Raymond, B., d’Udekem d’Acoz, C., Van de Putte, A., Danis, B., David, B., Grant, S., Gutt, J., et al., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 155–165. [Google Scholar]
- Kupchan, S.M.; Britton, R.W.; Ziegler, M.F.; Sigel, C.W. Bruceantin, a new potent antileukemic simaroubolide from Brucea antidysenterica. J. Org. Chem. 1973, 38, 178–179. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Deceptionin (7) | Methyl Pourewate A | ||
|---|---|---|---|---|
 |  | |||
| 1H, δ, mult, J (Hz) | 13C, ppm | 1H, δ, mult, J (Hz) | 13C, ppm | |
| 1a 1b | 1.29, m; 2.09, m | 39.1 | 1.26, m; 2.18, m | 39.2 |
| 2a 2b | 1.49, m; 1.80, m | 20.1 | 1.49, m; 1.94, m | 20.0 |
| 3a 3b | 1.21, m; 1.33, m | 39.9 | 1.18, m; 1.38, m | 40.1 |
| 4 | - | 31.6 | - | 31.6 |
| 5a 5b | 1.06, m; 1.77, d, 13.5 | 50.7 | 0.97, m; 1.84, m | 50.9 |
| 6 | 1.24, d, 6.9 | 15.9 | 1.20, d, 6.8 | 16.1 |
| 7 | 4.22, q, 6.9 | 41.9 | 4.18, q, 4.4 | 42.0 |
| 8 | - | 127.6 | - | 128.2 |
| 9 | - | 146.0 | - | 143.9 |
| 10 | - | 41.4 | - | 41.6 |
| 11a 11b | 1.94, ddd, 16.0, 14.0, 4.2; 2.23, ddd, 16.0, 4.2, 4.2 | 27.4 | 1.86, m; 2.20, m | 27.7 |
| 12a 12b | 1.26, m; 1.64, dq, 13.0, 4.2 | 30.2 | 1.24, m; 1.65, m | 31.2 |
| 13 | 2.41, m | 37.8 | 2.34, q, 7.6 | 38.0 |
| 14 | 2.91, dd, 8.1, 2.5 | 47.1 | 2.65, dd, 8.1, 2.4 | 49.0 |
| 15 | 5.87, d, 2.5 | 103.5 | 4.62, d, 2.4 | 110.8 |
| 16a 16b | 3.85, d, 8.6; 4.09, dd, 8.6, 5.6 | 76.0 | 3.76, dd, 8.8, 3.4; 4.02, dd, 8.5, 6.4 | 74.9 |
| 17 | - | 174.9 | - | 174.7 |
| 18 | 0.89, s | 27.1 | 0.92, s | 26.6 |
| 19 | 0.88, s | 32.6 | 0.88, s | 33.3 |
| 20 | 1.08, s | 30.4 | 1.02, s | 30.9 |
| OCOCH3 | - | 170.6 | ||
| OCOCH3 | 2.04, s | 21.5 | ||
| OCH3 (methyl ester) | 3.59, s | 51.6 | 3.67, s | 51.7 |
| OCH3 (methoxy) | 3.24, s | 54.6 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
|---|---|---|---|---|---|---|---|
| Natural habitat | |||||||
| Deception Island | 0.00 ± 0.01 | 0.16 ± 0.07 | 0.14 ± 0.12 | 0.92 ± 0.98 | 0.10 ± 0.18 | 0.01 ± 0.03 | 1.16 ± 0.30 |
| Livingston Island | 2.38 ± 2.01 | 0.11 ± 0.09 | 0.12 ± 0.10 | 2.99 ± 2.76 | 0.03 ± 0.07 | 0.07 ± 0.16 | 0.30 ± 0.42 |
| Heat stress experiment | |||||||
| CT | 0.02 ± 0.03 | 0.20 ± 0.07 | 0.20 ± 0.11 | 1.44 ± 0.85 | 0.55 ± 0.38 | 0.15 ± 0.14 | 1.04 ± 0.53 |
| HST | 0.01 ± 0.01 | 0.27 ± 0.09 | 0.20 ± 0.09 | 1.29 ± 1.53 | 0.67 ± 0.92 | 0.35 ± 0.24 | 1.57 ± 0.49 |
| EHST | 0.00 ± 0.01 | 0.14 ± 0.11 | 0.12 ± 0.10 | 0.45 ± 0.39 | 0.01 ± 0.02 | 0.41 ± 0.54 | 0.79 ± 0.62 |
| Predation experiment | |||||||
| Macropredation | 0.04 ± 0.04 | 0.21 ± 0.04 | 0.19 ± 0.06 | 1.34 ± 0.51 | 0.35 ± 0.40 | 0.12 ± 0.18 | 1.32 ± 0.15 |
| Micropredation | 0.02 ± 0.03 | 0.22 ± 0.04 | 0.21 ± 0.04 | 0.81 ± 0.84 | 0.21 ± 0.34 | 0.22 ± 0.28 | 1.28 ± 0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Castro-Fernández, P.; Angulo-Preckler, C.; García-Aljaro, C.; Avila, C.; Cutignano, A. A Chemo-Ecological Investigation of Dendrilla antarctica Topsent, 1905: Identification of Deceptionin and the Effects of Heat Stress and Predation Pressure on Its Terpene Profiles. Mar. Drugs 2023, 21, 499. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090499
De Castro-Fernández P, Angulo-Preckler C, García-Aljaro C, Avila C, Cutignano A. A Chemo-Ecological Investigation of Dendrilla antarctica Topsent, 1905: Identification of Deceptionin and the Effects of Heat Stress and Predation Pressure on Its Terpene Profiles. Marine Drugs. 2023; 21(9):499. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090499
Chicago/Turabian StyleDe Castro-Fernández, Paula, Carlos Angulo-Preckler, Cristina García-Aljaro, Conxita Avila, and Adele Cutignano. 2023. "A Chemo-Ecological Investigation of Dendrilla antarctica Topsent, 1905: Identification of Deceptionin and the Effects of Heat Stress and Predation Pressure on Its Terpene Profiles" Marine Drugs 21, no. 9: 499. https://0-doi-org.brum.beds.ac.uk/10.3390/md21090499