An Emerging Pathogen from Rotted Chestnut in China: Gnomoniopsis daii sp. nov.


The Key Laboratory for Silviculture and Conservation of Ministry of Education, Beijing Forestry University, Beijing 100083, China
*
Author to whom correspondence should be addressed.
Forests 2019, 10(11), 1016; https://0-doi-org.brum.beds.ac.uk/10.3390/f10111016
Submission received: 23 October 2019
/
Revised: 11 November 2019
/
Accepted: 11 November 2019
/
Published: 13 November 2019
(This article belongs to the Special Issue Emerging Pathogens in Forest Ecosystems)
Abstract
:Nut quality is fundamental to the economic viability of the Chinese sweet chestnut industry, but fruit rot disease significantly reduces this quality. In this study, we investigated chestnut rot in Anhui and Hubei provinces in China. Typical brown rot symptoms were observed, affecting nuts from different plantations. Isolates were obtained from symptomatic tissues of rotted fruits that were identified based on morphological comparison and phylogenetic analyses of partial internal transcribed spacer (ITS), and tef1 and tub2 gene sequences. The inoculation results showed that the tested fungal species is pathogenic to chestnut fruits. Hence, a new and severe pathogen that causes Chinese sweet chestnut brown rot, Gnomoniopsis daii sp. nov., is introduced herein.
1. Introduction
China has the largest chestnut industry in the world, producing more than 2 × 106 tons of chestnuts annually since 2013 [1]. The Chinese sweet chestnut (Castanea mollissima Bl.) is widely cultivated in most provinces in China, providing gluten-free, low fat, and cholesterol-free crop nuts for human consumption [2]. Chestnut orchards and stands are also important to the economy as sources of timber [3].
Traditionally, several pathogens were considered the causal agents of chestnut rot in China, including Alternaria Nees, Botryosphaeria Ces. & De Not., Colletotrichum Corda, Diaporthe Nitschke, Fusarium Link, and Penicillium Link species [4,5,6,7]. However, no detailed studies on these pathogens have been conducted in China in the past decade. European sweet chestnut (Castanea sativa Mill.), known as one of the four major chestnut species in the world, has been well studied in nut rot by several phytopathologists and taxonomists [8,9,10,11,12,13,14,15]. Several important fungal species, Cryphonectria parasitica M.E. Barr, Gnomoniopsis smithogilvyi L.A. Shuttlew., E.C.Y. Liew & D.I. Guest (syn. G. castaneae Tamietti), Phytophthora cinnamomi Rands, and Sirococcus castanea J.B. Mey., Senn-Irlet & T.N. Sieber, have been reported on Castanea sativa from Australia and Europe [16,17,18,19,20].
The genus Gnomoniopsis Berl. (Gnomoniaceae G. Winter, Diaporthales Nannf.) was first described as a subgenus within Gnomonia Ces. & De Not. for species having ascospores that develop additional septa [21]. However, the development of additional septa was thought to be an occasional occurrence; Gnomoniopsis was subsequently proposed as a synonym of Gnomonia [22]. Sogonov et al. reevaluated concepts of the leaf-inhabiting genera in Gnomoniaceae based on the DNA sequence data of these genera, and restricted the genus Gnomoniopsis to G. chamaemori Berl. (type), G. comari Sogonov, G. fructicola Sogonov, G. macounii Sogonov, G. paraclavulata Sogonov, G. racemula Sogonov, and G. toementillae Sogonov [21]. Subsequently, nine additional species were added to this genus [23]. Gnomoniopsis castaneae and G. smithogilvyi were described independently from Europe and Australia, but Shuttleworth et al. proved that both names refer to a single species based on a comparative morphological analysis and five-marker phylogenetic analysis [18].
During the surveys of chestnut rot conducted in Anhui and Hubei provinces in China, typical brown rot symptoms were observed (Figure 1). Our aim in this study was to identify pathogens associated with chestnut brown rot in China. We conducted pathogenicity tests on healthy nuts to assess their pathogenicity.
2. Materials and Methods
2.1. Sample Collection and Isolation
Anhui and Hubei provinces are two important chestnut production bases in China. Samples were randomly collected in local storehouses from different chestnut plantations after harvest, then packed in paper bags, and posted to the laboratory for further study. Rotted chestnuts were surface-sterilized for 1 min in 75% ethanol, 3 min in 1.25% sodium hypochlorite, and 1 min in 75% ethanol, then rinsed for 2 min in sterile water and blotted on dry sterile filter paper. Infected nut tissues were cut into small pieces (0.2 cm × 0.2 cm) using a sterile scalpel and transferred onto the surface of malt extract agar (MEA; 30 g malt extract, 5 g peptone, 15 g agar/L; Aobox Company Limited, Beijing, China). After inoculation, agar plates were left at 25 °C in the dark for 2 days. Then, single hyphal strands were transferred to fresh medium plates under a dissecting stereomicroscope with a sterile needle. Specimen of the new species was deposited in the Museum of Beijing Forestry University, Beijing, China (BJFC). The ex-type culture was maintained in the China Forestry Culture Collection Center, Beijing, China (CFCC).
2.2. DNA Extraction and Phylogenetic Analysis
Genomic DNA was extracted from 15-day-old mycelium grown on MEA using the CTAB (cetyltrimethylammonium bromide) method [24]. DNA sequences were generated for the internal transcribed spacer (ITS) regions including the 5.8S gene of the ribosomal RNA operon amplified with primers ITS1/ITS4 [25], the translation elongation factor 1a (tef1) amplified with primers EF1-728F/EF1-1567R [26], and the b-tubulin gene 2 (tub2) amplified with primers T1/Bt2b [27]. The PCR conditions were: initial denaturation step of 5 min at 94 °C, followed by 35 cycles of 30 s at 94 °C, 50 s at 48 °C (ITS) or 54 °C (tef1) or 52 °C (tub2), and 1 min at 72 °C, and a final elongation step of 7 min at 72 °C. The PCR amplification products were scored visually by electrophoresis in 2% agarose gel. The DNA sequencing was performed using an ABI Prism 3730xl DNA Analyzer (ABI, Foster City, CA, USA) with Big-Dye Terminator kit v.3.1 (Invitrogen, Beijing, China) at Shanghai Invitrogen Biological Technology Co. Ltd. (Beijing, China).
Sequences of the three individual loci (ITS, tef1 and tub2) were aligned and edited manually using MEGA6 (Table 1). Maximum likelihood (ML) analysis was used for phylogenetic inferences of the concatenated alignments. ML analysis was implemented on the CIPRES Science Gateway portal using RAxML-HPC BlackBox v. 8.2.10 based on single ITS and combined sequences of ITS, tef1, and tub2 [28]. The resulting trees were plotted using FigTree v. 1.4.2.
2.3. Morphological Identification and Characterization
Morphological descriptions of the new Gnomoniopsis species were based on cultures sporulating on MEA in the dark at 25 °C after a month. Micromorphological features were observed under a Leica compound microscope (DM 2500, Leica, Wetzlar, Germany). More than 50 conidia were randomly selected for measurement. Cultural characteristics of isolates incubated on MEA were observed and recorded, including colony color and texture.
2.4. Pathogenicity Trials
Four isolates representing Gnomoniopsis daii were selected for inoculations, viz., CMF002A (ex-type from Anhui province), CMF002B (from Anhui province), CMF095 (from Hubei province), and CMF098 (from Hubei province). Isolates were allowed to grow on MEA for one week at 25 °C before the tests. We collected 160 asymptomatic nuts from a chestnut orchard in Anhui province, and 10 of them were randomly chosen and dissected to confirm healthy status. The remaining 150 chestnuts were surface-sterilized for 1 min in 75% ethanol, 3 min in 1.25% sodium hypochlorite, and 1 min in 75% ethanol, then rinsed for 2 min in sterile water and blotted on dry sterile filter paper. Using a cork borer (7 mm diameter), we wounded the nuts by removing the seed coat to expose the seed. Same-sized gar discs were removed from the actively growing margins of cultures and placed into the wounds with the mycelium facing the exposed seed. Sterile MEA discs were used for the negative controls. Wounds with the inoculated mycelium or sterile MEA were covered with masking tape to prevent contamination and desiccation. We ran 30 replicates for each strain and negative control. These inoculated nuts were maintained in a greenhouse at 25 °C. After 15 days, all the replicates were examined for disease, and re-isolations were conducted for all the symptomatic nuts.
3. Results
3.1. Fungal Isolation and Identification
Most of the pieces from infected nut tissue yielded a fungus, and 125 isolates were obtained. The isolates were primarily identified based on the morphology of conidia formed on the plates and ITS sequences. As a result, four isolates were Alternaria, six isolates were Botryosphaeria, 53 isolates were Colletotrichum, seven isolates were Diaporthe, 42 isolates were Gnomoniopsis, 11 isolates were Fusarium, and two isolates were Penicillium. Only one fungus was obtained from one rotted chestnut. For the first time in China, Gnomoniopsis isolates were obtained from rotted chestnut. Hence, detailed studies on them were conducted during the present study.
3.2. Phylogeny
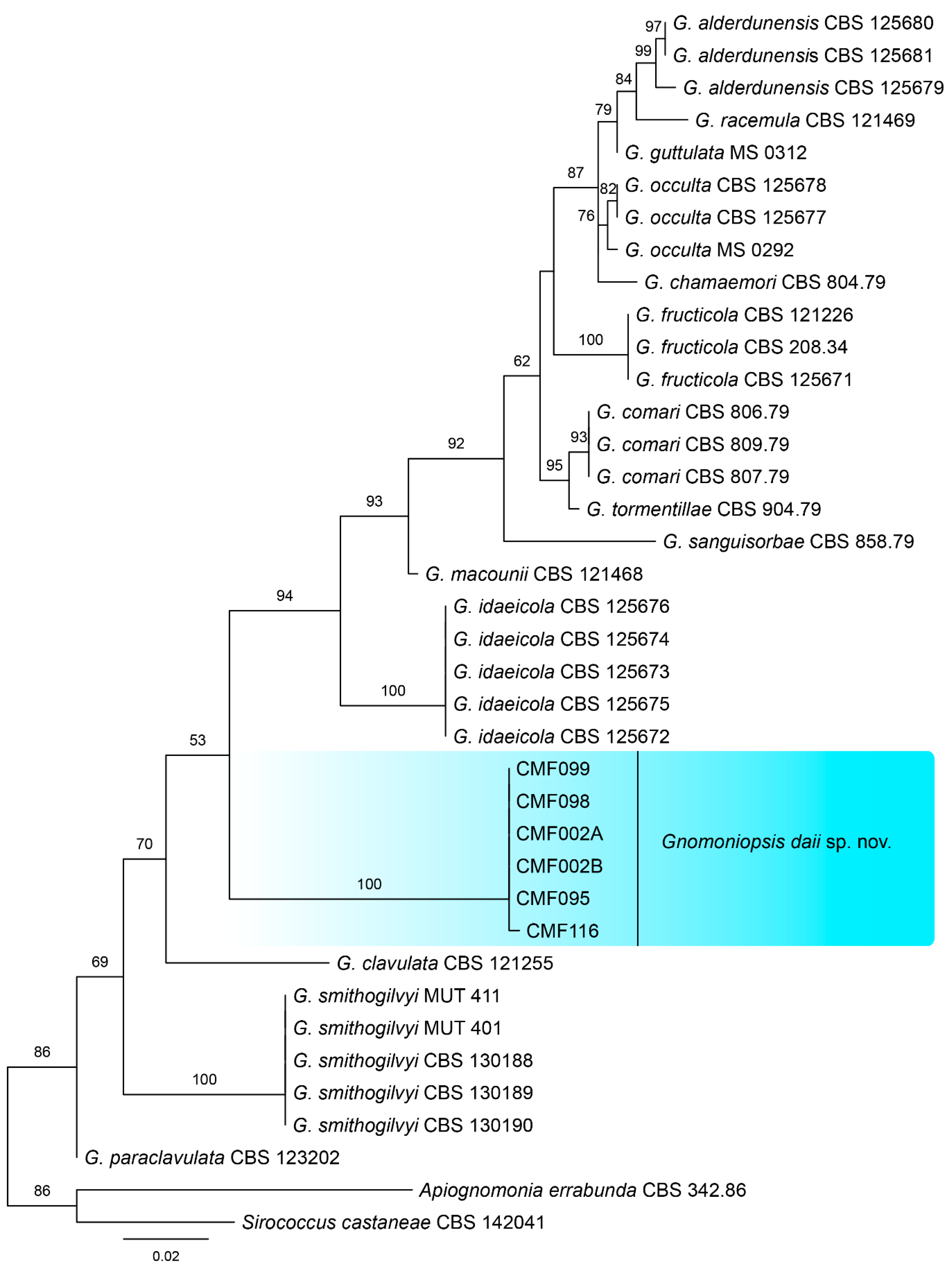
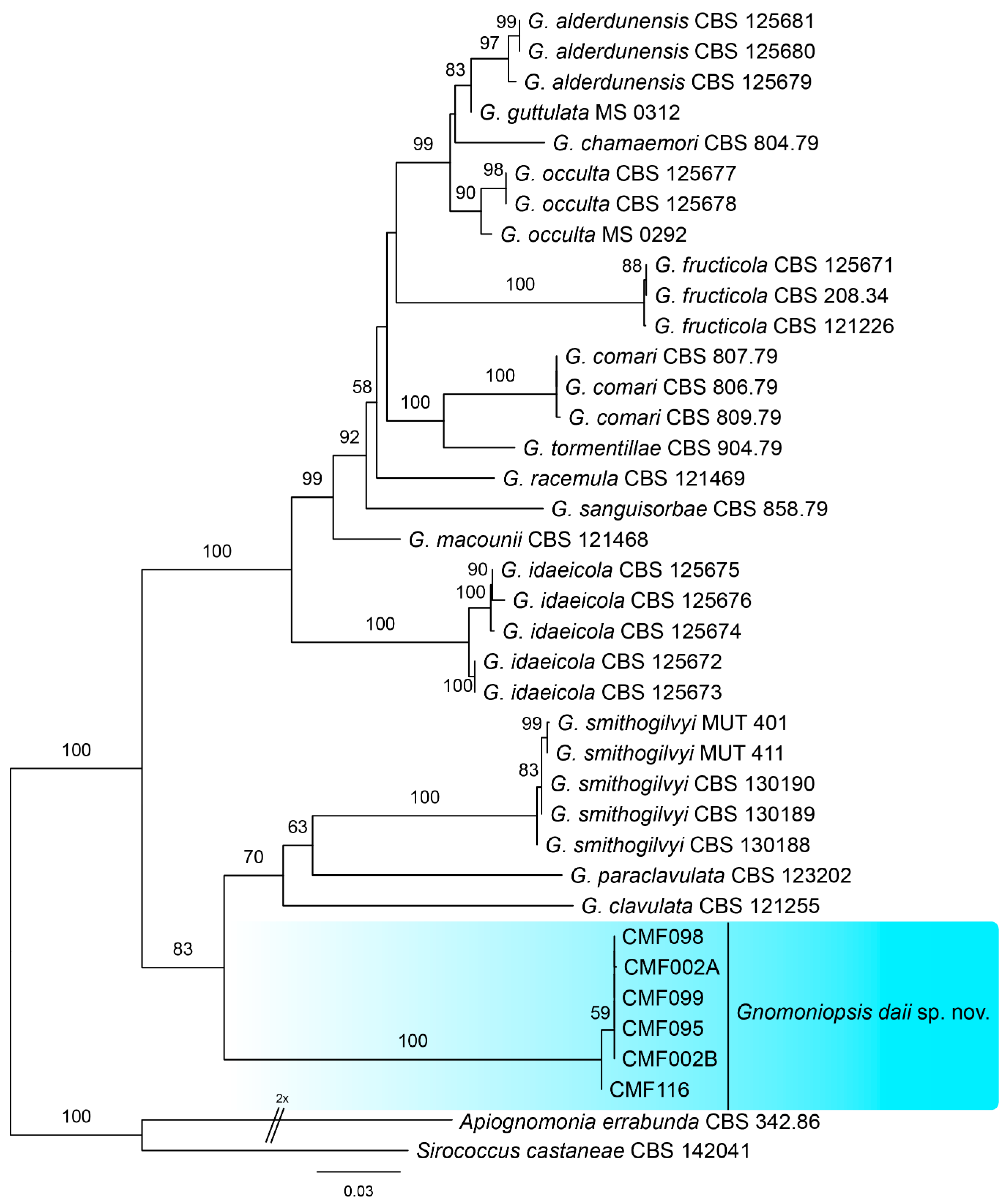
To identify the phylogenetic position of our isolates within Gnomoniopsis, phylogenetic analyses were performed based on ITS and combined ITS, tef1, and tub2 sequence data. The ITS alignment contained 38 sequences (including one outgroup) with 542 characters including alignment gaps. Of these, 428 characters were constant, 34 were variable and parsimony-uninformative, and 80 were parsimony-informative. The six Gnomoniopsis strains from this study form a well-supported clade distinguished from known species (Figure 2). The combined ITS, tef1, and tub2 alignment contained 38 sequences (including one outgroup) and 1685 characters including alignment gaps; 960 of these were parsimony-informative, 174 were variable and parsimony-uninformative, and 551 were constant. A similar phylogram was obtained from multi-genes to single ITS (Figure 3), which indicated strains from this study as a new Gnomoniopsis species. All 42 Gnomoniopsis isolates were identical in our primary comparison; hence, the six are shown in Figure 2 and Figure 3.
3.3. Morphology and Taxonomy
MycoBank MB 833088
Holotype: BJFC-C005
Etymology: in honor of Fanglan Dai, who is one of the most famous Chinese taxonomists.
Host/Distribution: on rotted Castanea mollissima fruits in China.
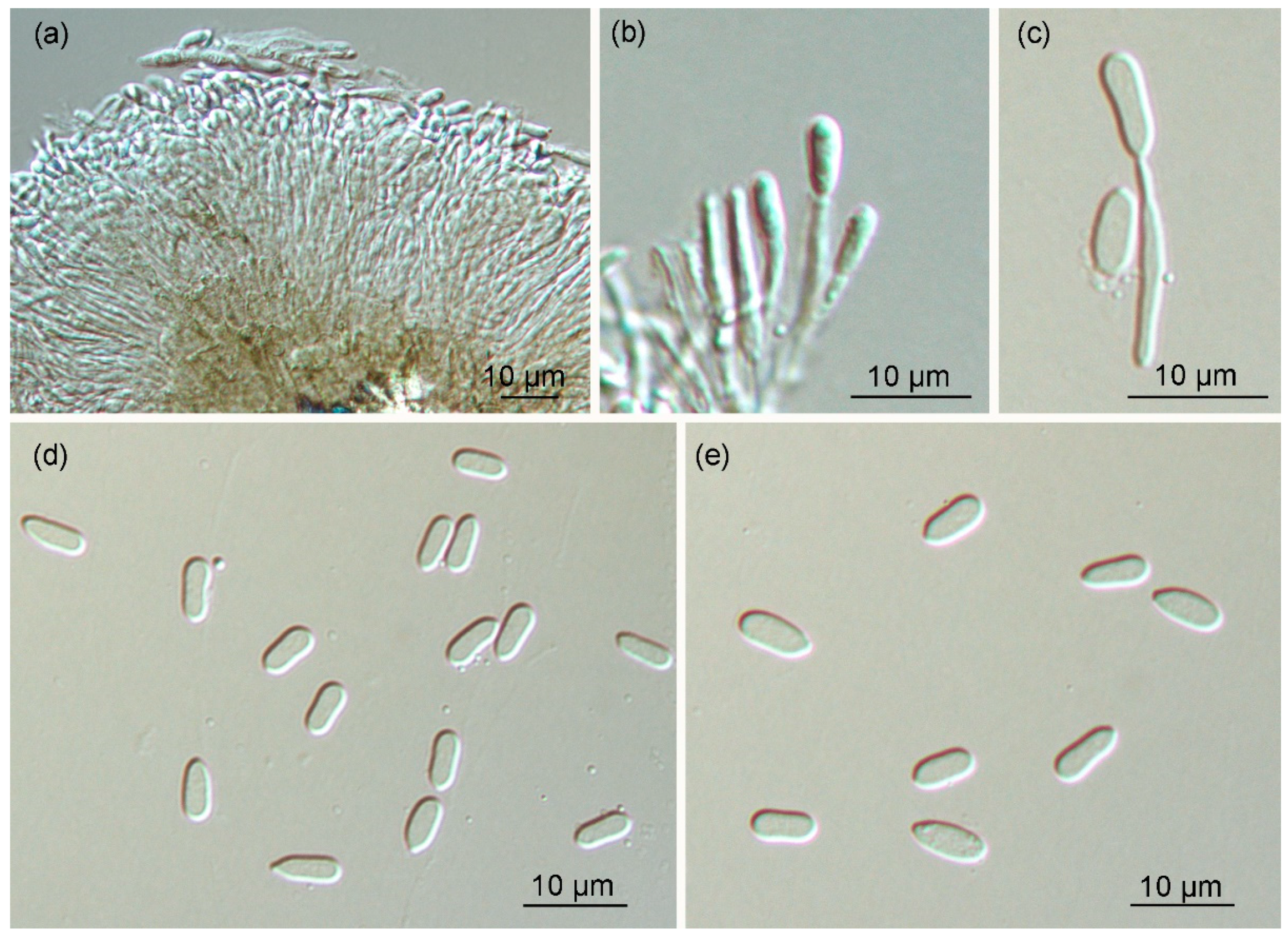
Original description: Colonies on MEA attaining 60 mm in one week at 25 °C, with undulate margin, whitish; after one month at 25 °C, light orange to white conidiomata distributed irregularly on the surface. Pycnidia globose to oval, solitary or confluent, light orange to white, 150–950 μm diameter; conidiophores indistinct, often reduced to conidiogenous cells. Conidiogenous cells oval, hyaline, one-celled, 5–18 µm. Conidia oval, oblate, fusiform, straight to slightly curved, hyaline, finely guttulate or not, (5.0–)5.5–7.0(–8.0) × 2.0–3.5 µm.
Material examined: CHINA, Anhui province, Liuan city, on rotted fruits of Castanea mollissima, Ning Jiang and Chengming Tian, 7 October 2017 (BJFC-C005 holotype; ex-type culture, CMF002A = CFCC 54043); Liuan city, on rotted fruits of Castanea mollissima, Ning Jiang and Chengming Tian, 7 October 2017 (living culture, CMF002B). Hubei province, Huanggang city, on rotted fruits of Castanea mollissima, Ning Jiang and Chengming Tian, 20 September 2019 (living culture, CMF095, CMF098, CMF099, CMF116).
Notes: Gnomoniopsis daii has similar conidia to G. smithogilvyi (5.0–8.0 × 2.0–3.5 µm in Gnomoniopsis daii vs. 6.1–9.8 × 2.4–4.9 µm in G. smithogilvyi), but they are different in host species and distribution (Gnomoniopsis daii on Castanea mollissima in China vs. G. smithogilvyi on Castanea sativa in Europe) [18]. They are obviously separated in the phylogram base on ITS (Figure 2) and combined ITS, tef1, and tub2 (Figure 3).
3.4. Pathogenicity Trials
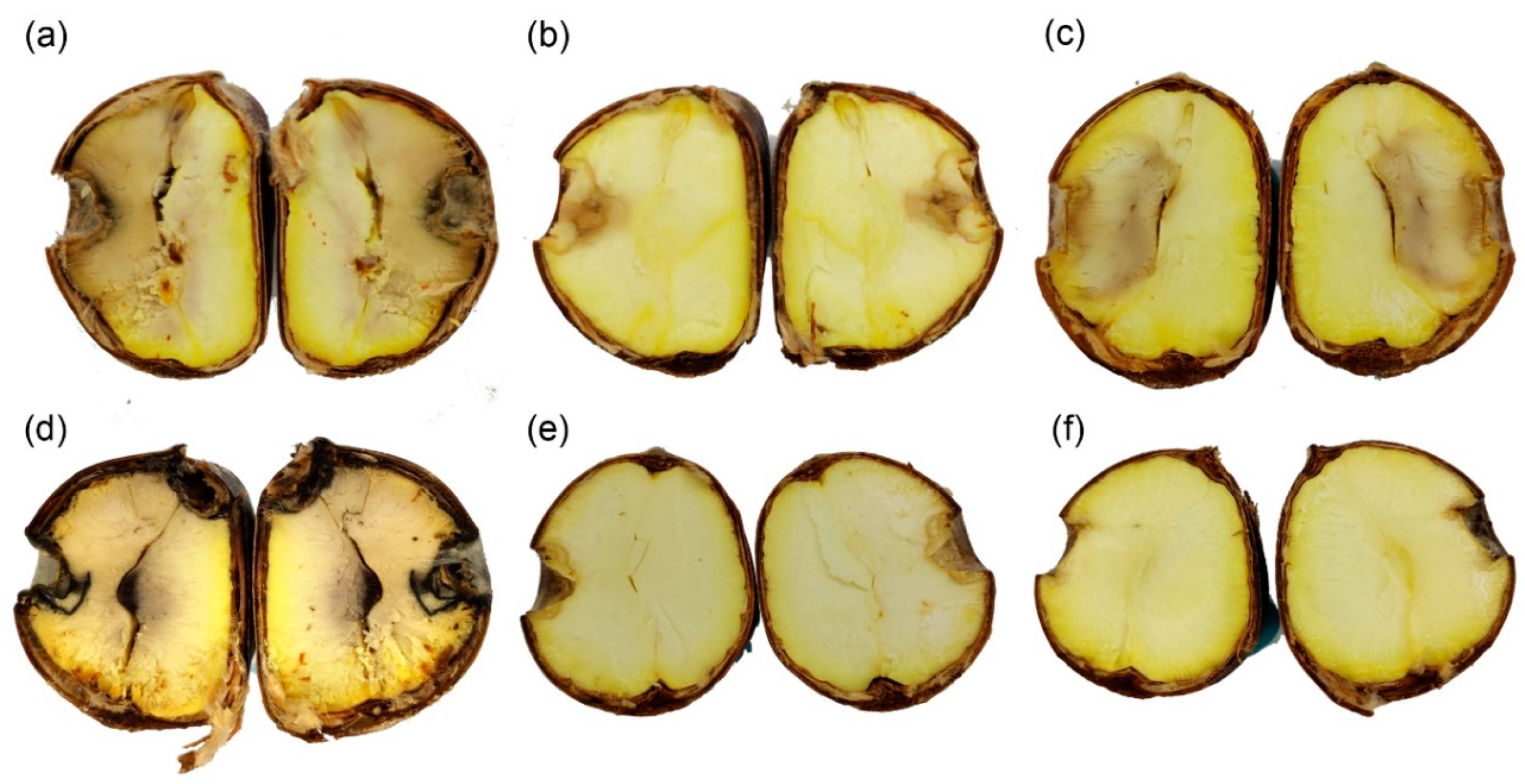
All 10 nuts assayed to test their health were found to be intact. The four tested strains showed brown rot symptoms and were detected in 83% of the artificially infected nuts (Figure 6). No obvious differences were found among the four strains (Table 2). Re-isolates were obtained from affected nuts and identified based on ITS sequence, which were all Gnomoniopsis daii. The asymptomatic nuts and negative controls did not show any symptoms.
4. Discussion
In this paper, Gnomoniopsis daii is introduced as a new species in the genus Gnomoniopsis. This species was found to be an emerging causal agent of chestnut brown rot in Anhui and Hubei provinces in China. Identification for describing the fungus as a new taxon was based on the results of phylogenetic analyses of sequence data for combined ITS and tef1 and tub2 genes, as well as the morphological characteristics. However, only the asexual state of Gnomoniopsis daii was discovered from rotted seeds of chestnut trees.
The related species of Gnomoniopsis daii, G. smithogilvyi, has been reported to cause serious disease on Castanea sativa and C. crenata × C. sativa hybrids in Europe and Oceania [13,19]. The infection process and cycle of chestnut disease has been demonstrated, and ascospores of G. smithogilvyi from chestnut buds are key to causing fruit rot [11,13]. The pathogen was later isolated from cankers on stems and branches [11,13]. However, we did not discover the sexual morph of Gnomoniopsis daii on the chestnut bud during this study.
Gnomoniopsis species inhabited three families of host, viz., Fagaceae, Rosaceae, and Onagraceae [18,20,23]. Gnomoniopsis daii, G. smithogilvyi, G. clavulata, and G. paraclavulata were discovered from Fagaceae trees and formed a close phylogenetic relationship differing from other species (Figure 2 and Figure 3). Gnomoniopsis smithogilvyi was first reported as a nut rot pathogen [8]. Subsequently, the authors isolated this fungus from chestnut branches [13]. Gnomoniopsis daii was described as a novel pathogen of chestnut rot disease in China depending on its asexual state. Gnomoniopsis clavulata and G. paraclavulata were collected from overwintered leaves belonging to Quercus species in the form of sexual states [21]. These two species were only reported in the USA [21]. Conidial size can only barely separate these four close species (5.0–8.0 × 2.0–4.0 µm in G. clavulata vs. 5.0–8.0 × 2.0–3.5 µm in G. daii vs. 6.0–9.5 × 2.0–3.5 µm in G. paraclavulata vs. 4.9–9.8 × 2.9–4.9 µm in G. smithogilvyi) [8,20,23], but the combined evidence of host species, distribution, and molecular data (ITS, tef1, and tub2) clearly distinguishes these related species.
During our pathogenicity test, we confirmed that Gnomoniopsis daii also causes chestnut brown rot. Hence, this Gnomoniopsis species represents the second species in this genus infecting Castanea hosts. Castanea is an important plant genus worldwide, so it is necessary to further research the fundamental aspects of the relationship between the pathogen genus Gnomoniopsis and host genus Castanea.
Accurate identification and diagnostics of fungal pathogens are important for determining the disease cycle and route of transmission. As an emergent disease agent in chestnut orchards in China, chestnut tree loss appears to be closely associated with Gnomoniopsis nut rot. Further studies should focus on methods to prevent increased damage to this valuable crop tree.
5. Conclusions
A novel fungal species, Gnomoniopsis daii, is an emerging pathogen causing Chinese sweet chestnut brown rot in China.
Author Contributions
Experiments: N.J.; writing—original draft preparation: N.J.; writing—review and editing: N.J. and C.T.
Funding
This research was funded by the National Natural Science Committee, grant number 31670647.
Acknowledgments
We are grateful for the assistance of Yingmei Liang (Museum of Beijing Forestry University), Chungen Piao and Minwei Guo (China Forestry Culture Collection Center) during this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yi, S.J. Situation and development strategy of chestnut industry in China. J. West China For. Sci. 2017, 46, 132–149. [Google Scholar] [CrossRef]
- Lu, C.; Guo, S.J. Analysis on the nutritional characters and comprehensive evaluation of 16 chestnut germplasm resources. Sci. Technol. Food Ind. 2017, 37, 357–376. [Google Scholar] [CrossRef]
- Wang, J.R.; Liu, M.C.; Yang, L.; Min, Q.W. Factors affecting the willingness of farmers to accept eco-compensation in the Qianxi chestnut agroforestry system, Hebei. J. Resour. Ecol. 2018, 9, 407–415. [Google Scholar] [CrossRef]
- He, W.; Shen, R.X.; Wang, X.J. Pathogenicity of pathogens contributing to dry rot of Chinese chestnut and their infection process. J. Beijing For. Univ. 2001, 23, 36–39. [Google Scholar]
- He, W.; Yin, W.L.; Shen, R.X.; Wang, X.J. Latent infection of fungal pathogens and pathogenesis of Chinese chestnut seed rot. Sci. Silvae Sin. 2004, 40, 96–102. [Google Scholar]
- Hou, B.L.; Zhang, Z.M.; Yang, X.M.; Shang, J.L.; Xu, F.S.; Ma, C.F. The study on the kernel spot diseases of Chinese chestnut in Hebei. J. Hebei Agric. Univ. 1988, 11, 11–22. [Google Scholar]
- Zhu, J.R.; Lian, Y.Y.; Hua, Z.Y. Preliminary exploration into the cause of Chinese chestnut putrity in storage. J. Zhejiang For. Technol. 1992, 12, 44–46. [Google Scholar]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Decock, C.A.; Dreyer, L.L.; Granke, L.L.; Guest, D.I.; Hardy, G.E.St.J.; Hausbeck, M.K.; et al. Fungal Planet Description Sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Dennert, F.G.; Broggini, G.A.; Gessler, C.; Storari, M. Gnomoniopsis castanea is the main agent of chestnut nut rot in Switzerland. Phytopathol. Mediterr. 2015, 54, 199–211. [Google Scholar]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Alves, A.; Abdollahzadeh, J.; Phillips, A.J.L. Phylogeny, morphology and pathogenicity of Botryosphaeriaceae, Diatrypaceae and Gnomoniaceae associated with branch diseases of hazelnut in Sardinia (Italy). Eur. J. Plant. Pathol. 2016, 146, 259–279. [Google Scholar] [CrossRef]
- Pasche, S.; Calmin, G.; Auderset, G.; Crovadore, J.; Pelleteret, P.; Mauch-Mani, B.; Barja, F.; Paul, B.; Jermini, M.; Lefort, F. Gnomoniopsis smithogilvyi causes chestnut canker symptoms in Castanea sativa shoots in Switzerland. Fungal Genet. Biol. 2016, 87, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Seddaiu, S.; Cerboneschi, A.; Sechi, C.; Mello, A. Gnomoniopsis castaneae associated with Dryocosmus kuriphilus galls in chestnut stands in Sardinia (Italy). IFOREST 2017, 10, 440–445. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Guest, D.I. The infection process of chestnut rot, an important disease caused by Gnomoniopsis smithogilvyi (Gnomoniaceae, Diaporthales) in Oceania and Europe. Australas. Plant. Pathol. 2017, 46, 397–405. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Liew, E.C.Y.; Guest, D.I. Survey of the incidence of chestnut rot in south-eastern Australia. Australas. Plant. Pathol. 2013, 42, 63–72. [Google Scholar] [CrossRef]
- Şimşek, S.A.; Katircioğlu, Y.Z.; Serçe, Ç.U.; Çakar, D.; Rigling, D.; Maden, S. Phytophthora species associated with dieback of sweet chestnut in Western Turkey. For. Pathol. 2019, 49, e12533. [Google Scholar] [CrossRef]
- Meyer, J.B.; Trapiello, E.; Senn-Irlet, B.; Sieber, T.N.; Cornejo, C.; Aghayeva, D.; González, A.J.; Prosperoa, S. Phylogenetic and phenotypic characterisation of Sirococcus castaneae comb. nov. (synonym Diplodina castaneae), a fungal endophyte of European chestnut. Fungal Biol. UK 2017, 121, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant. Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Walker, D.M.; Guest, D.I. The chestnut pathogen Gnomoniopsis smithogilvyi (Gnomoniaceae, Diaporthales) and its synonyms. Mycotaxon 2016, 130, 929–940. [Google Scholar] [CrossRef]
- Trapiello, E.; Feito, I.; González, A.J. First Report of Gnomoniopsis castaneae Causing Canker on Hybrid Plants of Castanea sativa× C. crenata in Spain. Plant. Dis. 2018, 102, 1040. [Google Scholar] [CrossRef]
- Visentin, I.; Gentile, S.; Valentino, D.; Gonthier, P.; Tamietti, G.; Cardinale, F. Gnomoniopsis castanea sp. nov.(Gnomoniaceae, Diaporthales) as the causal agent of nut rot in sweet chestnut. J. Plant. Pathol. 2012, 94, 411–419. [Google Scholar]
- Sogonov, M.V.; Castlebury, L.A.; Rossman, A.Y.; Mejía, L.C.; White, J.F. Leaf-inhabiting genera of the Gnomoniaceae, Diaporthales. Stud. Mycol. 2008, 62, 1–77. [Google Scholar] [CrossRef] [PubMed]
- Barr, M.E. The Diaporthales in North America with emphasis on Gnomonia and its segregates. Mycol. Mem. 1978, 7, 1–232. [Google Scholar]
- Walker, D.M.; Castlebury, L.A.; Rossman, A.Y.; Sogonov, M.V.; White, J.F. Systematics of genus Gnomoniopsis (Gnomoniaceae, Diaporthales) based on a three gene phylogeny, host associations and morphology. Mycologia 2010, 102, 1479–1496. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
Figure 1.
Symptoms of chestnut brown rot: (a,b) diseased nuts and (c) discolored kernels.

Figure 2.
Consensus tree resulting from a RAxML analysis of ITS sequence alignment for species of Gnomoniopsis. The scale bar represents the expected number of changes per site.
Figure 2.
Consensus tree resulting from a RAxML analysis of ITS sequence alignment for species of Gnomoniopsis. The scale bar represents the expected number of changes per site.

Figure 3.
Consensus tree resulting from a RAxML analysis of combined ITS, tef1, and tub2 sequence alignment for species of Gnomoniopsis. The scale bar represents the expected number of changes per site.
Figure 3.
Consensus tree resulting from a RAxML analysis of combined ITS, tef1, and tub2 sequence alignment for species of Gnomoniopsis. The scale bar represents the expected number of changes per site.

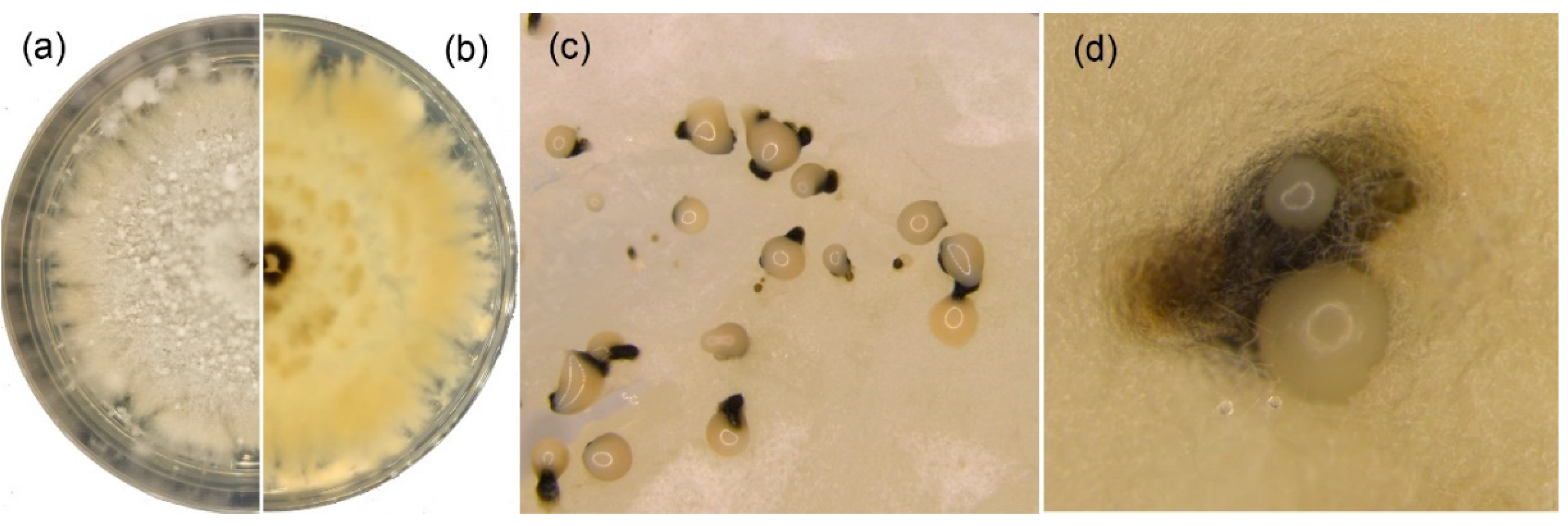
Figure 4.
Gnomoniopsis daii (BJFC-C005 from CMF002A) cultures on MEA. (a,b) Colony on MEA after 15 days at 25 °C; (c,d) conidiomata formed on MEA.
Figure 4.
Gnomoniopsis daii (BJFC-C005 from CMF002A) cultures on MEA. (a,b) Colony on MEA after 15 days at 25 °C; (c,d) conidiomata formed on MEA.

Figure 5.
Morphology of Gnomoniopsis daii (BJFC-C005 from CMF002A): (a–c) conidiogenous cells with attached conidia and (d,e) conidia.
Figure 5.
Morphology of Gnomoniopsis daii (BJFC-C005 from CMF002A): (a–c) conidiogenous cells with attached conidia and (d,e) conidia.

Figure 6.
Results of pathogenicity trials after 15 days: (a–d) Mild to severe symptoms caused by CMF002A (ex-type), (e) asymptomatic chestnut after incubation, and (f) negative control.
Figure 6.
Results of pathogenicity trials after 15 days: (a–d) Mild to severe symptoms caused by CMF002A (ex-type), (e) asymptomatic chestnut after incubation, and (f) negative control.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Isolates and GenBank accession numbers used in this study.
| Species | Country | Host | Strain | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| Internal Transcribed Spacer (ITS) | tub2 | tef1 | ||||
| Apiognomonia veneta | France | Platanus occidentalis | CBS 342.86 | DQ313531 | EU219235 | DQ318036 |
| Gnomoniopsis alderdunensis | USA | Rubus pedatus | CBS 125679 | GU320826 | GU320788 | GU320813 |
| Gnomoniopsis alderdunensis | USA | Rubus parviflorus | CBS 125680 | GU320825 | GU320787 | GU320801 |
| Gnomoniopsis alderdunensis | USA | Rubus parviflorus | CBS 125681 | GU320827 | GU320789 | GU320802 |
| Gnomoniopsis chamaemori | Finland | Rubus chamaemorus | CBS 804.79 | GU320817 | GU320777 | GU320809 |
| Gnomoniopsis clavulata | USA | Quercus falcata | CBS 121255 | EU254818 | EU219211 | GU320807 |
| Gnomoniopsis comari | Finland | Comarum palustre | CBS 806.79 | EU254821 | EU219156 | GU320810 |
| Gnomoniopsis comari | Finland | Comarum palustre | CBS 807.79 | EU254822 | GU320779 | GU320814 |
| Gnomoniopsis comari | Switzerland | Comarum palustre | CBS 809.79 | EU254823 | GU320778 | GU320794 |
| Gnomoniopsis daii | China | Castanea mollissima | CMF002A | MN598671 | MN605519 | MN605517 |
| Gnomoniopsis daii | China | Castanea mollissima | CMF002B | MN598672 | MN605520 | MN605518 |
| Gnomoniopsis daii | China | Castanea mollissima | CMF095 | MN598673 | NA | NA |
| Gnomoniopsis daii | China | Castanea mollissima | CMF098 | MN598674 | NA | NA |
| Gnomoniopsis daii | China | Castanea mollissima | CMF099 | MN598675 | NA | NA |
| Gnomoniopsis daii | China | Castanea mollissima | CMF116 | MN598676 | NA | NA |
| Gnomoniopsis fructicola | USA | Fragaria vesca | CBS 121226 | EU254824 | EU219144 | GU320792 |
| Gnomoniopsis fructicola | France | Fragaria sp. | CBS 208.34 | EU254826 | EU219149 | GU320808 |
| Gnomoniopsis fructicola | USA | Fragaria sp. | CBS 125671 | GU320816 | GU320776 | GU320793 |
| Gnomoniopsis guttulata | Bulgaria | Agrimonia eupatoria | NA | EU254812 | NA | NA |
| Gnomoniopsis idaeicola | USA | Rubus sp. | CBS 125672 | GU320823 | GU320781 | GU320797 |
| Gnomoniopsis idaeicola | USA | Rubus pedatus | CBS 125673 | GU320824 | GU320782 | GU320798 |
| Gnomoniopsis idaeicola | France | Rubus sp. | CBS 125674 | GU320820 | GU320780 | GU320796 |
| Gnomoniopsis idaeicola | USA | Rubus procerus | CBS 125675 | GU320822 | GU320783 | GU320799 |
| Gnomoniopsis idaeicola | USA | Rubus procerus | CBS 125676 | GU320821 | GU320784 | GU320811 |
| Gnomoniopsis macounii | USA | Spiraea sp. | CBS 121468 | EU254762 | EU219126 | GU320804 |
| Gnomoniopsis occulta | USA | Potentilla sp. | CBS 125677 | GU320828 | GU320785 | GU320812 |
| Gnomoniopsis occulta | USA | Potentilla sp. | CBS 125678 | GU320829 | GU320786 | GU320800 |
| Gnomoniopsis occulta | Russia | Potentilla anserina | NA | EU254811 | NA | NA |
| Gnomoniopsis paraclavulata | USA | Quercus alba | CBS 123202 | GU320830 | GU320775 | GU320815 |
| Gnomoniopsis racemula | USA | Chamerion angustifolium | CBS 121469 | EU254841 | EU219125 | GU320803 |
| Gnomoniopsis sanguisorbae | Switzerland | Sanguisorba minor | CBS 858.79 | GU320818 | GU320790 | GU320805 |
| Gnomoniopsis smithogilvyi | Australia | Castanea sp. | CBS 130190 | JQ910642 | JQ910639 | KR072534 |
| Gnomoniopsis smithogilvyi | Australia | Castanea sp. | CBS 130189 | JQ910644 | JQ910641 | KR072535 |
| Gnomoniopsis smithogilvyi | Australia | Castanea sp. | CBS 130188 | JQ910643 | JQ910640 | KR072536 |
| Gnomoniopsis smithogilvyi | Italy | Castanea sativa | MUT 401 | HM142946 | KR072532 | KR072537 |
| Gnomoniopsis smithogilvyi | New Zealand | Castanea sativa | MUT 411 | HM142948 | KR072533 | KR072538 |
| Gnomoniopsis tormentillae | Switzerland | Potentilla sp. | CBS 904.79 | EU254856 | EU219165 | GU320795 |
| Sirococcus castaneae | Switzerland | Castanea sativa | CBS 142041 | KX929744 | KX958443 | KX929710 |
Note: NA, not applicable. Strains in this study are identified in bold.
Table 2.
Results of pathogenicity trials.
| Strain | No. of Affected Nuts | No. of Asymptomatic Nuts |
|---|---|---|
| CMF002A | 26 | 4 |
| CMF002B | 24 | 6 |
| CMF095 | 22 | 8 |
| CMF098 | 23 | 7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jiang, N.; Tian, C. An Emerging Pathogen from Rotted Chestnut in China: Gnomoniopsis daii sp. nov. Forests 2019, 10, 1016. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111016
AMA Style
Jiang N, Tian C. An Emerging Pathogen from Rotted Chestnut in China: Gnomoniopsis daii sp. nov. Forests. 2019; 10(11):1016. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111016
Chicago/Turabian StyleJiang, Ning, and Chengming Tian. 2019. "An Emerging Pathogen from Rotted Chestnut in China: Gnomoniopsis daii sp. nov." Forests 10, no. 11: 1016. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.