Deep Learning Approaches for the Mapping of Tree Species Diversity in a Tropical Wetland Using Airborne LiDAR and High-Spatial-Resolution Remote Sensing Images


Abstract
:1. Introduction
2. Study Area and Materials
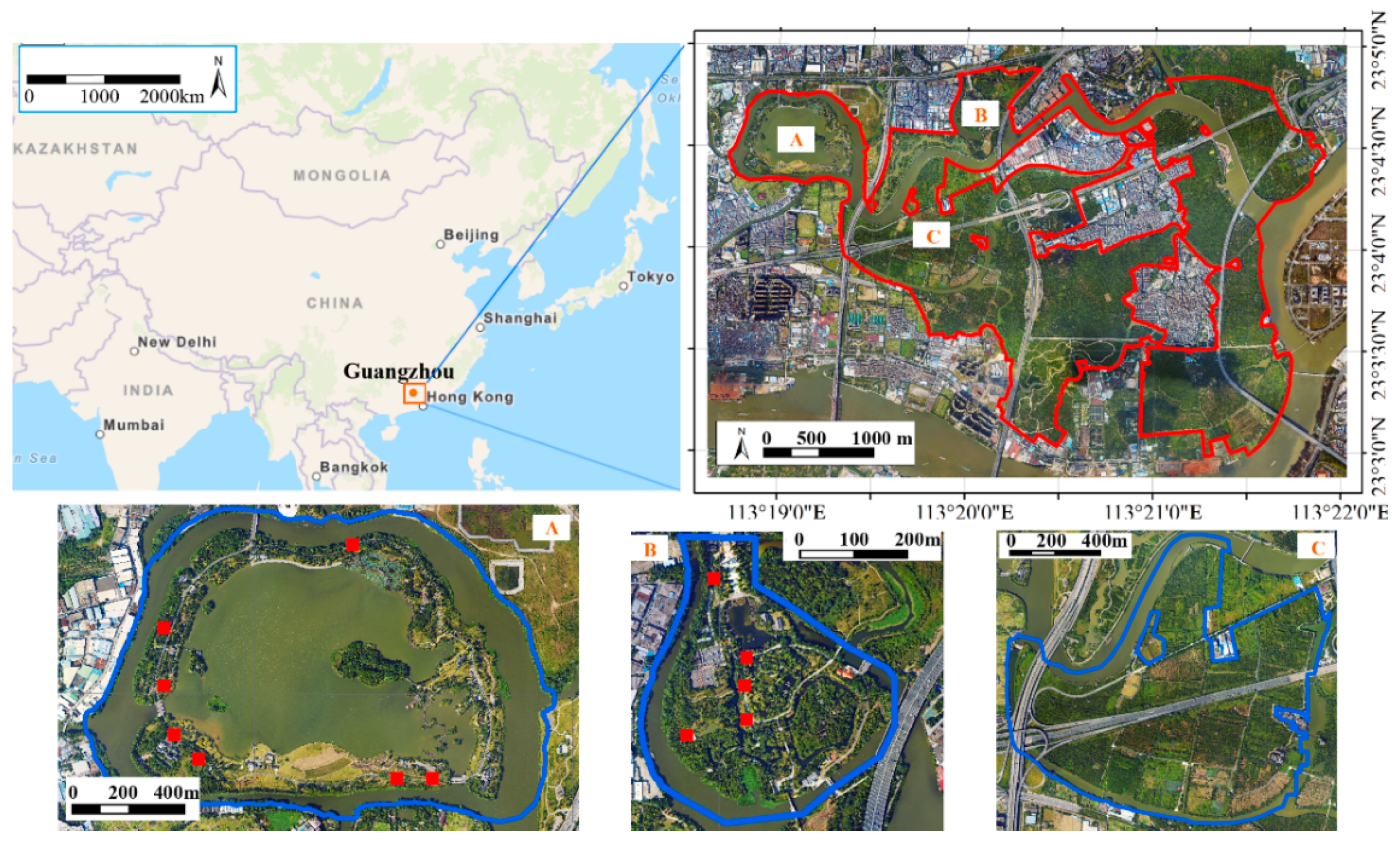
2.1. Study Area
2.2. Field Survey
2.3. Remotely Sensed Data
3. Methodology
3.1. Overview
3.2. Individual Tree Detection
3.3. Deep Learning Methods for Tree Species Classification
3.4. Forest Species Diversity Mapping
3.5. Experimental Setup
3.6. Assessment
4. Results
4.1. Individual Tree Species Classification Results: AlexNet, VGG16, and ResNet50
4.2. Forest Species Diversity Mapping
4.2.1. Visual Performance
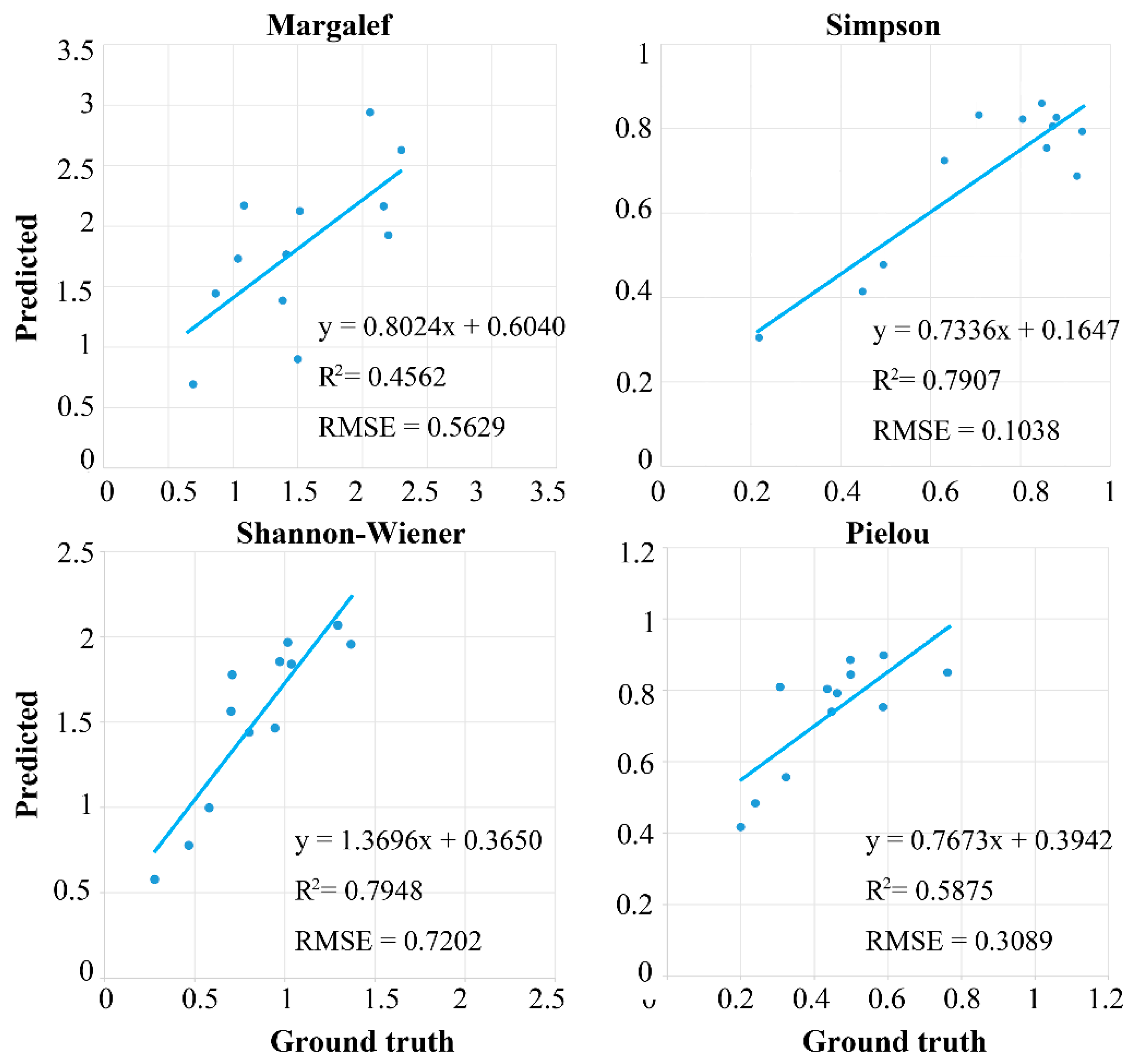
4.2.2. Accuracy Assessment
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schäfer, E.; Heiskanen, J.; Heikinheimo, V.; Pellikka, P. Mapping tree species diversity of a tropical montane forest by unsupervised clustering of airborne imaging spectroscopy data. Ecol. Indic. 2016, 64, 49–58. [Google Scholar] [CrossRef]
- Ferreira, M.P.; Zortea, M.; Zanotta, D.C.; Shimabukuro, Y.E.; de Souza Filho, C.R. Mapping tree species in tropical seasonal semi-deciduous forests with hyperspectral and multispectral data. Remote Sens. Environ. 2016, 179, 66–78. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Gillson, L.; Duffin, K. Thresholds of potential concern as benchmarks in the management of African savannahs. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 309–319. [Google Scholar] [CrossRef]
- Madonsela, S.; Cho, M.A.; Ramoelo, A.; Mutanga, O.; Naidoo, L. Estimating tree species diversity in the savannah using NDVI and woody canopy cover. Int. J. Appl. Earth Obs. Geoinf. 2018, 66, 106–115. [Google Scholar] [CrossRef]
- Jetz, W.; Cavender-Bares, J.; Pavlick, R.; Schimel, D.; Davis, F.W.; Asner, G.P.; Guralnick, R.; Kattge, J.; Latimer, A.M.; Moorcroft, P. Monitoring plant functional diversity from space. Nat. Plants 2016, 2, 16024. [Google Scholar] [CrossRef] [PubMed]
- Nagendra, H.; Lucas, R.; Honrado, J.P.; Jongman, R.H.; Tarantino, C.; Adamo, M.; Mairota, P. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Hernández, H.; Galleguillos, M.; Fassnacht, F. Comparing generalized linear models and random forest to model vascular plant species richness using LiDAR data in a natural forest in central Chile. Remote Sens. Environ. 2016, 173, 200–210. [Google Scholar] [CrossRef]
- Nagendra, H.; Gadgil, M. Satellite imagery as a tool for monitoring species diversity: An assessment. J. Appl. Ecol. 1999, 36, 388–397. [Google Scholar] [CrossRef]
- Nagendra, H. Using remote sensing to assess biodiversity. Int. J. Remote Sens. 2001, 22, 2377–2400. [Google Scholar] [CrossRef]
- Kuenzer, C.; Ottinger, M.; Wegmann, M.; Guo, H.; Wang, C.; Zhang, J.; Dech, S.; Wikelski, M. Earth observation satellite sensors for biodiversity monitoring: Potentials and bottlenecks. Int. J. Remote Sens. 2014, 35, 6599–6647. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, Y.; Zheng, Z.; Dong, W.; Zhao, D.; Wu, B.; Zhao, Q. Forest species diversity mapping using airborne LiDAR and hyperspectral data in a subtropical forest in China. Remote Sens. Environ. 2018, 213, 104–114. [Google Scholar] [CrossRef]
- Yang, C.; Everitt, J.H.; Fletcher, R.S.; Jensen, R.R.; Mausel, P.W. Evaluating AISA+ hyperspectral imagery for mapping black mangrove along the South Texas Gulf Coast. Photogramm. Eng. Remote Sens. 2009, 75, 425–435. [Google Scholar] [CrossRef]
- Nevalainen, O.; Honkavaara, E.; Tuominen, S.; Viljanen, N.; Hakala, T.; Yu, X.; Hyyppä, J.; Saari, H.; Pölönen, I.; Imai, N. Individual tree detection and classification with UAV-based photogrammetric point clouds and hyperspectral imaging. Remote Sens. 2017, 9, 185. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Tree species classification in the Southern Alps based on the fusion of very high geometrical resolution multispectral/hyperspectral images and LiDAR data. Remote Sens. Environ. 2012, 123, 258–270. [Google Scholar] [CrossRef]
- Goodenough, D.G.; Dyk, A.; Niemann, K.O.; Pearlman, J.S.; Chen, H.; Han, T.; Murdoch, M.; West, C. Processing Hyperion and ALI for forest classification. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1321–1331. [Google Scholar] [CrossRef]
- Féret, J.-B.; Asner, G.P. Semi-supervised methods to identify individual crowns of lowland tropical canopy species using imaging spectroscopy and LiDAR. Remote Sens. 2012, 4, 2457–2476. [Google Scholar] [CrossRef]
- Hakkenberg, C.; Zhu, K.; Peet, R.; Song, C. Mapping multi-scale vascular plant richness in a forest landscape with integrated Li DAR and hyperspectral remote-sensing. Ecology 2018, 99, 474–487. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Schöning, I. Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles. Methods Ecol. Evolut. 2012, 3, 397–404. [Google Scholar] [CrossRef]
- Carleer, A.; Wolff, E. Exploitation of very high resolution satellite data for tree species identification. Photogramm. Eng. Remote Sens. 2004, 70, 135–140. [Google Scholar] [CrossRef]
- van Lier, O.R.; Fournier, R.A.; Bradley, R.L.; Thiffault, N. A multi-resolution satellite imagery approach for large area mapping of ericaceous shrubs in Northern Quebec, Canada. Int. J. Appl. Earth Obs. Geoinform. 2009, 11, 334–343. [Google Scholar] [CrossRef]
- Wang, L.; Sousa, W.P.; Gong, P.; Biging, G.S. Comparison of IKONOS and QuickBird images for mapping mangrove species on the Caribbean coast of Panama. Remote Sens. Environ. 2004, 91, 432–440. [Google Scholar] [CrossRef]
- Sasaki, T.; Imanishi, J.; Ioki, K.; Morimoto, Y.; Kitada, K. Object-based classification of land cover and tree species by integrating airborne LiDAR and high spatial resolution imagery data. Lands. Ecol. Eng. 2012, 8, 157–171. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Clark, D.B. Hyperspectral discrimination of tropical rain forest tree species at leaf to crown scales. Remote Sens. Environ. 2005, 96, 375–398. [Google Scholar] [CrossRef]
- Rosenfeld, A.; Zemel, R.; Tsotsos, J.K. The elephant in the room. arXiv 2018, arXiv:1808.03305. [Google Scholar]
- Ma, L.; Liu, Y.; Zhang, X.; Ye, Y.; Yin, G.; Johnson, B.A. Deep learning in remote sensing applications: A meta-analysis and review. ISPRS J. Photogramm. Remote Sens. 2019, 152, 166–177. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, X.; Xin, Q.; Huang, J. Developing a multi-filter convolutional neural network for semantic segmentation using high-resolution aerial imagery and LiDAR data. ISPRS J. Photogramm. Remote Sens. 2018, 143, 3–14. [Google Scholar] [CrossRef]
- Xu, G.; Zhu, X.; Fu, D.; Dong, J.; Xiao, X. Automatic land cover classification of geo-tagged field photos by deep learning. Environ. Model. Softw. 2017, 91, 127–134. [Google Scholar] [CrossRef]
- Sun, Z.; Zhao, X.; Wu, M.; Wang, C. Extracting Urban Impervious Surface from WorldView-2 and Airborne LiDAR Data Using 3D Convolutional Neural Networks. J. Indian Soc. Remote Sens. 2019, 47, 401–412. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, X.; Xin, Q.; Sun, Y.; Zhang, P. Automatic building extraction from high-resolution aerial images and LiDAR data using gated residual refinement network. ISPRS J. Photogramm. Remote Sens. 2019, 151, 91–105. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, Z. Rbnet: A deep neural network for unified road and road boundary detection. Int. Conf. Neural Inf. Process. 2017, 677–687. [Google Scholar] [CrossRef]
- Du, Z.; Yang, J.; Ou, C.; Zhang, T. Smallholder Crop Area Mapped with a Semantic Segmentation Deep Learning Method. Remote Sens. 2019, 11, 888. [Google Scholar] [CrossRef]
- Persello, C.; Tolpekin, V.; Bergado, J.; de By, R. Delineation of agricultural fields in smallholder farms from satellite images using fully convolutional networks and combinatorial grouping. Remote Sens. Environ. 2019, 231, 111253. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, J.-D.; Drolet, G.; Brown, N. Mapping dead forest cover using a deep convolutional neural network and digital aerial photography. ISPRS J. Photogramm. Remote Sens. 2019, 156, 14–26. [Google Scholar] [CrossRef]
- Flood, N.; Watson, F.; Collett, L. Using a U-net convolutional neural network to map woody vegetation extent from high resolution satellite imagery across Queensland, Australia. Int. J. Appl. Earth Obs. Geoinform. 2019, 82, 101897. [Google Scholar] [CrossRef]
- Li, W.; Fu, H.; Yu, L.; Cracknell, A. Deep learning based oil palm tree detection and counting for high-resolution remote sensing images. Remote Sens. 2016, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Popescu, S.C.; Wynne, R.H.; Nelson, R.F. Estimating plot-level tree heights with lidar: Local filtering with a canopy-height based variable window size. Comput. Electron. Agric. 2002, 37, 71–95. [Google Scholar] [CrossRef]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. Imagenet classification with deep convolutional neural networks. Adv. Neural Inf. Process. Syst. 2012, 25, 1097–1105. [Google Scholar] [CrossRef]
- Simonyan, K.; Zisserman, A. Very deep convolutional networks for large-scale image recognition. arXiv 2014, arXiv:1409.1556. Available online: https://arxiv.org/abs/1409.1556 (accessed on 10 April 2015).
- Szegedy, C.; Liu, W.; Jia, Y.; Sermanet, P.; Reed, S.; Anguelov, D.; Erhan, D.; Vanhoucke, V.; Rabinovich, A. Going deeper with convolutions. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Boston, MA, USA, 7–12 June 2015; pp. 1–9. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Rocchini, D.; Hernández-Stefanoni, J.L.; He, K.S. Advancing species diversity estimate by remotely sensed proxies: A conceptual review. Ecol. Inf. 2015, 25, 22–28. [Google Scholar] [CrossRef]
- Margalef, D. Information theory in ecology, General systems. Transl. Mem. Real Acad. Cienc. Artes Barcelona 1958, 32, 373–449. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.; Weiner, W. The Mathematical Theory of Communication; Urban University Illinois Press: Champaign, IL, USA, 1963; 125p. [Google Scholar]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Jia, Y.; Shelhamer, E.; Donahue, J.; Karayev, S.; Long, J.; Girshick, R.B.; Guadarrama, S.; Darrell, T. Caffe: Convolutional Architecture for Fast Feature Embedding. ACM Multimed. 2014, 675–678. [Google Scholar] [CrossRef]
- Kingma, D.P.; Ba, J. Adam: A Method for Stochastic Optimization. In Proceedings of the International Conference on Learning Representations, San Diego, CA, USA, 7–9 May 2015. [Google Scholar]
- Martin, D.R.; Fowlkes, C.C.; Malik, J. Learning to detect natural image boundaries using local brightness, color, and texture cues. IEEE Trans. Pattern Anal. Mach. Intell. 2004, 26, 530–549. [Google Scholar] [CrossRef]
- Jung, H.; Choi, M.-K.; Jung, J.; Lee, J.-H.; Kwon, S.; Young Jung, W. ResNet-based vehicle classification and localization in traffic surveillance systems. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition Workshops, Honolulu, HI, USA, 21–26 July 2017; pp. 61–67. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Indices and Description | Definition | Remarks |
|---|---|---|
| Margalef Richness Index: An index to measure the number of species in a certain region. | : number of species; the total number of all individuals. | |
| Simpson Diversity Index: An index that takes into account the number of species, as well as the relative abundance of each species. | the total number of species ; the total number of all individuals; the number of species. | |
| Shannon–Wiener Diversity Index: An index that indicates the relationship between species and community complexity. | : the total number of species ; the total number of all individuals; the number of species. | |
| Pielou Evenness Index 1: An index that refers to the distribution of the total number of species and the number of individuals in a community. | the result of the Shannon–Wiener diversity index; the number of species. |
| Tree Type | VGG16 (140,000) | ResNet50 (110,000) | AlexNet (100,000) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| UA (%) | PA (%) | F1-Score | UA (%) | PA (%) | F1-Score | UA (%) | PA (%) | F1-Score | |
| Silk floss tree | 30.61 | 55.56 | 39.47 | 33.33 | 44.44 | 38.10 | 28.57 | 44.44 | 34.78 |
| Banyan tree | 59.77 | 76.47 | 67.10 | 54.63 | 86.76 | 67.05 | 53.19 | 73.53 | 61.73 |
| Flame tree | 80.70 | 90.20 | 85.19 | 83.64 | 90.20 | 86.79 | 76.79 | 84.31 | 80.37 |
| Longan | 40.38 | 80.77 | 53.85 | 47.92 | 88.46 | 62.16 | 36.00 | 69.23 | 47.37 |
| Banana | 93.75 | 100.00 | 96.77 | 91.84 | 100.00 | 95.74 | 87.23 | 91.11 | 89.13 |
| Papaya | 100.00 | 100.00 | 100.00 | 95.83 | 100.00 | 97.87 | 92.00 | 100.00 | 95.83 |
| Bauhinia | 77.17 | 81.61 | 79.33 | 75.26 | 83.91 | 79.35 | 72.94 | 71.26 | 72.09 |
| Eucalyptus trees | 88.00 | 100.00 | 93.62 | 84.62 | 100.00 | 91.67 | 78.18 | 97.73 | 86.87 |
| Carambola | 86.67 | 76.47 | 81.25 | 70.00 | 82.35 | 75.68 | 63.64 | 82.35 | 71.79 |
| Sakura tree | 100.00 | 100.00 | 100.00 | 96.88 | 96.88 | 96.88 | 96.97 | 100.00 | 98.46 |
| Pond cypress | 88.89 | 100.00 | 94.12 | 83.33 | 83.33 | 83.33 | 68.75 | 91.67 | 78.57 |
| Alstonia scholaris | 71.43 | 83.33 | 76.92 | 68.63 | 83.33 | 75.27 | 80.00 | 66.67 | 72.72 |
| Bischofia javanica | 66.00 | 89.19 | 75.86 | 59.62 | 83.78 | 69.66 | 68.18 | 81.08 | 74.07 |
| Hibiscus tiliaceus | 76.92 | 100.00 | 86.96 | 86.36 | 95.00 | 90.48 | 83.33 | 100.00 | 90.91 |
| Litchi | 50.00 | 15.00 | 23.08 | 80.00 | 40.00 | 53.33 | 33.33 | 15.00 | 20.69 |
| Mango tree | 60.00 | 28.57 | 38.71 | 80.00 | 38.10 | 51.61 | 38.46 | 23.81 | 29.41 |
| Camphor tree | 44.44 | 27.59 | 34.04 | 33.33 | 24.14 | 28.00 | 48.00 | 41.38 | 44.44 |
| Others | 79.11 | 59.14 | 67.68 | 83.5 | 55.48 | 66.67 | 76.15 | 55.15 | 63.97 |
| OA = 73.25% Kappa = 69.76% | OA = 72.93% Kappa = 69.62% | OA = 68.53% Kappa = 64.52% | |||||||
| Tree Species | Area A | Area B | Area C |
|---|---|---|---|
| Others | 2212 (28.98%) | 900 (30.19%) | 6186 (20.79%) |
| Silk floss tree | 696 (9.12%) | 364 (12.21%) | 2099 (7.05%) |
| Banyan tree | 1049 (13.74%) | 359 (12.04%) | 1508 (5.07%) |
| Flame tree | 505 (6.62%) | 115 (3.86%) | 651 (2.19%) |
| Longan | 570 (7.47%) | 303 (10.16%) | 10735 (36.07%) |
| Banana | 49 (0.64%) | 178 (5.97%) | 2497 (8.39%) |
| Papaya | 20 (0.26%) | 2 (0.07%) | 155 (0.52%) |
| Bauhinia | 741 (9.71%) | 234 (7.85%) | 1352 (4.54%) |
| Eucalyptus trees | 382 (5.00%) | 53 (1.78%) | 555 (1.87%) |
| Carambola | 89 (1.17%) | 137 (4.60%) | 1141 (3.83%) |
| Sakura tree | 38 (0.50%) | 2 (0.07%) | 475 (1.60%) |
| Pond cypress | 150 (1.96%) | 19 (0.64%) | 123 (0.41%) |
| Alstonia scholaris | 336 (4.40%) | 36 (1.21%) | 227 (0.76%) |
| Bischofia javanica | 191 (2.50%) | 57 (1.91%) | 278 (0.93%) |
| Hibiscus tiliaceus | 136 (1.78%) | 6 (0.20%) | 500 (1.68%) |
| Litchi | 94 (1.23%) | 95 (3.19%) | 951 (3.20%) |
| Mango tree | 71 (0.93%) | 67 (2.25%) | 128 (0.43%) |
| Camphor tree | 305 (4.00%) | 54 (1.81%) | 197 (0.66%) |
| Species Richness | Plot Number | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| Ground truth | 8 | 9 | 4 | 6 | 7 | 3 | 5 | 5 | 4 | 6 | 8 | 5 |
| Prediction | 9 | 8 | 6 | 4 | 7 | 3 | 5 | 9 | 6 | 8 | 11 | 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Huang, J.; Ao, Z.; Lao, D.; Xin, Q. Deep Learning Approaches for the Mapping of Tree Species Diversity in a Tropical Wetland Using Airborne LiDAR and High-Spatial-Resolution Remote Sensing Images. Forests 2019, 10, 1047. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111047
Sun Y, Huang J, Ao Z, Lao D, Xin Q. Deep Learning Approaches for the Mapping of Tree Species Diversity in a Tropical Wetland Using Airborne LiDAR and High-Spatial-Resolution Remote Sensing Images. Forests. 2019; 10(11):1047. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111047
Chicago/Turabian StyleSun, Ying, Jianfeng Huang, Zurui Ao, Dazhao Lao, and Qinchuan Xin. 2019. "Deep Learning Approaches for the Mapping of Tree Species Diversity in a Tropical Wetland Using Airborne LiDAR and High-Spatial-Resolution Remote Sensing Images" Forests 10, no. 11: 1047. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111047