Drought-Induced Reductions and Limited Recovery in the Radial Growth, Transpiration, and Canopy Stomatal Conductance of Mongolian Scots Pine (Pinus sylvestris var. mongolica Litv): A Five-Year Observation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Layout
2.3. Meteorological Data
2.4. Measurements of Tree Diameter
2.5. Soil Moisture Measurements
2.6. Sap Flux Density Measurements and Canopy Transpiration Estimation
2.7. Calculation of Canopy Stomatal Conductance
2.8. Statistical Analyses
3. Results
3.1. Climate Factors and Soil Moisture
3.2. Tree Radial Growth
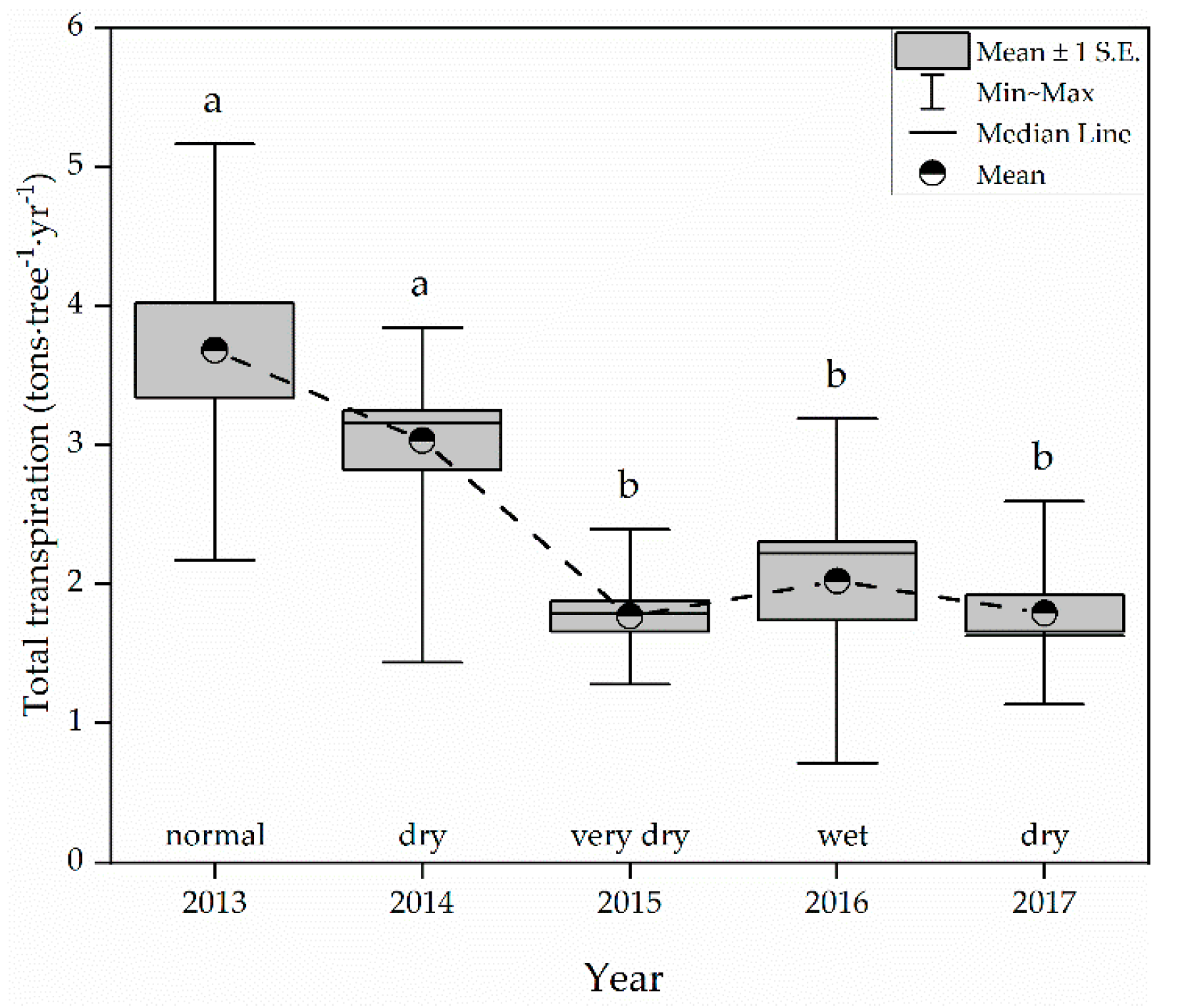
3.3. Transpiration Levels in Different Years
3.4. Seasonal Dynamics in Tree Transpiration
3.5. Seasonal Dynamics in Canopy Stomatal Conductance
3.6. Diurnal Courses of Qt and Gs in Different Years and Different Parts of the Growing Season
4. Discussion
4.1. Drought-Induced Tree Radial Growth Decline
4.2. Drought-Induced Reduction of Water Use
4.3. Limited Recovery after a Prolonged Drought Relief
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ryan, M.G. Tree responses to drought. Tree Physiol. 2011, 31, 237–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaltonen, H.; Lindén, A.; Heinonsalo, J.; Biasi, C.; Pumpanen, J. Effects of prolonged drought stress on Scots pine seedling carbon allocation. Tree Physiol. 2017, 37, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetter, S. Drought, change and resilience in South Africa’s arid and semi-arid rangelands. S. Afr. J. Sci. 2009, 105, 29–33. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; López, B.C.; Loepfe, L.; Lloret, F. Stand-and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 2012, 168, 877–888. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an inciting mortality factor in Scots pine stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Linton, M.; Sperry, J.; Williams, D. Limits to water transport in Juniperus osteosperma and Pinus edulis: Implications for drought tolerance and regulation of transpiration. Funct. Ecol. 1998, 12, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Piñol, J.; Sala, A. Ecological implications of xylem cavitation for several Pinaceae in the Pacific Northern USA. Funct. Ecol. 2000, 14, 538–545. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Hereş, A.-M.; Voltas, J.; López, B.C.; Martínez-Vilalta, J. Drought-induced mortality selectively affects Scots pine trees that show limited intrinsic water-use efficiency responsiveness to raising atmospheric CO2. Funct. Plant Biol. 2014, 41, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Prat, E.; Oliveras, I.; Piñol, J. Xylem hydraulic properties of roots and stems of nine Mediterranean woody species. Oecologia 2002, 133, 19–29. [Google Scholar] [CrossRef]
- Rebetez, M.; Dobbertin, M. Climate change may already threaten Scots pine stands in the Swiss Alps. Theor. Appl. Climatol. 2004, 79, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.-M.; Kellomäki, S.; Peltola, H.; Zhou, X.; Wang, K.-Y.; Väisänen, H. Impacts of changing climate on the productivity of Norway spruce dominant stands with a mixture of Scots pine and birch in relation to water availability in southern and northern Finland. Tree Physiol. 2011, 31, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Timofeeva, G.; Treydte, K.; Bugmann, H.; Rigling, A.; Schaub, M.; Siegwolf, R.; Saurer, M. Long-term effects of drought on tree-ring growth and carbon isotope variability in Scots pine in a dry environment. Tree Physiol. 2017, 37, 1028–1041. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Hoffmann, W.A.; Meinzer, F.C.; Franco, A.C.; Giambelluca, T.; Miralles-Wilhelm, F. Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agric. For. Meteorol. 2008, 148, 839–849. [Google Scholar] [CrossRef]
- Verbeeck, H.; Steppe, K.; Nadezhdina, N.; de Beeck, M.O.; Deckmyn, G.; Meiresonne, L.; Lemeur, R.; Čermák, J.; Ceulemans, R.; Janssens, I.A. Stored water use and transpiration in Scots pine: A modeling analysis with ANAFORE. Tree Physiol. 2007, 27, 1671–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hereş, A.-M.; Camarero, J.J.; López, B.C.; Martínez-Vilalta, J. Declining hydraulic performances and low carbon investments in tree rings predate Scots pine drought-induced mortality. Trees 2014, 28, 1737–1750. [Google Scholar] [CrossRef]
- Børja, I.; Svĕtlík, J.; Nadezhdin, V.; Čermák, J.; Rosner, S.; Nadezhdina, N. Sap flux—A real time assessment of health status in Norway spruce. Scand. J. For. Res. 2016, 31, 450–457. [Google Scholar] [CrossRef]
- Peters, M.P.; Iverson, L.R.; Matthews, S.N. Long-term droughtiness and drought tolerance of eastern US forests over five decades. For. Ecol. Manag. 2015, 345, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Macinnis-Ng, C.; Wyse, S.; Veale, A.; Schwendenmann, L.; Clearwater, M. Sap flow of the southern conifer, Agathis australis during wet and dry summers. Trees 2016, 30, 19–33. [Google Scholar] [CrossRef]
- Granier, A.; Breda, N.; Biron, P.; Villette, S. A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecol. Model. 1999, 116, 269–283. [Google Scholar] [CrossRef]
- Nadezhdina, N. Sap flow index as an indicator of plant water status. Tree Physiol. 1999, 19, 885–891. [Google Scholar] [CrossRef]
- Bovard, B.; Curtis, P.; Vogel, C.; Su, H.-B.; Schmid, H. Environmental controls on sap flow in a northern hardwood forest. Tree Physiol. 2005, 25, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Santana, V.; Rodriguez-Dominguez, C.M.; Fernández, J.E.; Diaz-Espejo, A. Role of leaf hydraulic conductance in the regulation of stomatal conductance in almond and olive in response to water stress. Tree Physiol. 2016, 36, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Poyatos, R.; Aguadé, D.; Galiano, L.; Mencuccini, M.; Martínez-Vilalta, J. Drought-induced defoliation and long periods of near-zero gas exchange play a key role in accentuating metabolic decline of Scots pine. New Phytol. 2013, 200, 388–401. [Google Scholar] [CrossRef]
- Meir, P.; Mencuccini, M.; Dewar, R.C. Drought-related tree mortality: Addressing the gaps in understanding and prediction. New Phytol. 2015, 207, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Tatarinov, F.; Rotenberg, E.; Maseyk, K.; Ogée, J.; Klein, T.; Yakir, D. Resilience to seasonal heat wave episodes in a Mediterranean pine forest. New Phytol. 2016, 210, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.; Konings, A.G.; Trugman, A.T.; Yu, K.-L.; Bowling, D.R.; Gabbitas, R.; Karp, D.S.; Pacala, S.; Sperry, J.S.; Sulman, B.N.; et al. Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature 2018, 561, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; Lopez, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Zeppel, M.J.; Anderegg, W.R.; Bloemen, J.; De Kauwe, M.G.; Hudson, P.; Ruehr, N.K.; Powell, T.L.; von Arx, G.; Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: Processes and trade-offs. Ecol. Res. 2018, 33, 839–855. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Walter, J.; Nagy, L.; Hein, R.; Rascher, U.; Beierkuhnlein, C.; Willner, E.; Jentsch, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot. 2011, 71, 34–40. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress memory and the inevitable effects of drought: A physiological perspective. Front. Plant Sci. 2016, 7, 143. [Google Scholar] [CrossRef] [Green Version]
- Peltier, D.M.; Fell, M.; Ogle, K. Legacy effects of drought in the southwestern United States: A multi-species synthesis. Ecol. Monogr. 2016, 86, 312–326. [Google Scholar] [CrossRef]
- Wu, X.-C.; Liu, H.-Y.; Li, X.-Y.; Ciais, P.; Babst, F.; Guo, W.; Zhang, C.; Magliulo, V.; Pavelka, M.; Liu, S.; et al. Differentiating drought legacy effects on vegetation growth over the temperate Northern Hemisphere. Glob. Chang. Biol. 2018, 24, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.-R. Report on the causes of the early decline of Pinus slyvestris var. mongolica shelterbelt and its preventative and control measures in Zhanggutai of Liaoning Province. Sci. Silvae Sin. 2001, 37, 131–138. [Google Scholar]
- Zhu, J.-J.; Zeng, D.-H.; Kang, H. Decline of Pinus Sylvestris var. Mongolica Plantations on Sandy Land; Chinese Forestry Press: Beijing, China, 2005. [Google Scholar]
- Zhu, J.; Li, F.; Xu, M.; Kang, H.; Wu, X. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China: An experimental approach. Ann. For. Sci. 2008, 65, 304. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Zhu, J.; Yan, Q.; Song, L. Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land, Northeast China. Agric. Water Manag. 2012, 109, 94–106. [Google Scholar] [CrossRef]
- Jiang, F.-Q.; Cao, C.-Y.; Zeng, D.-H. Degradation and Restoration of Ecosystems on Keerqin Sandy Land; Chinese Forestry Press: Beijing, China, 2002. [Google Scholar]
- Song, L.-N.; Zhu, J.-J.; Yan, Q.-L.; Li, M.-C.; Yu, G.-Q. Comparison of intrinsic water use efficiency between different aged Pinus sylvestris var. mongolica wide windbreaks in semiarid sandy land of northern China. Agrofor. Syst. 2015, 89, 477–489. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Zhang, J.; Zheng, X.; Wang, K. Canopy transpiration of Pinus sylvestris var. mongolica in a sparse wood grassland in the semiarid sandy region of Northeast China. Agric. For. Meteorol. 2018, 250, 192–201. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Yu, Z. Water utilization of Pinus sylvestris var. mongolica in a sparse wood grassland in the semiarid sandy region of Northeast China. Trees 2014, 28, 971–982. [Google Scholar] [CrossRef]
- Köstner, B.; Biron, P.; Siegwolf, R.; Granier, A. Estimates of water vapor flux and canopy conductance of Scots pine at the tree level utilizing different xylem sap flow methods. Theor. Appl. Climatol. 1996, 53, 105–113. [Google Scholar] [CrossRef]
- Kume, T.; Onozawa, Y.; Komatsu, H.; Tsuruta, K.; Shinohara, Y.; Umebayashi, T.; Otsuki, K. Stand-scale transpiration estimates in a Moso bamboo forest:(I) Applicability of sap flux measurements. For. Ecol. Manag. 2010, 260, 1287–1294. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed.; Springer-Verlag: New York, NY, USA, 1998; pp. 37–50. [Google Scholar]
- Bernier, P.; Bréda, N.; Granier, A.; Raulier, F.; Mathieu, F. Validation of a canopy gas exchange model and derivation of a soil water modifier for transpiration for sugar maple (Acer saccharum Marsh.) using sap flow density measurements. For. Ecol. Manag. 2002, 163, 185–196. [Google Scholar] [CrossRef]
- Poyatos, R.; Llorens, P.; Gallart, F. Transpiration of montane Pinus sylvestris L. and Quercus pubescens Willd. forest stands measured with sap flow sensors in NE Spain. Hydrol. Earth Syst. Sci. 2005, 2, 1011–1046. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.W.; Bond, B.J.; Jones, J.A.; Meinzer, F.C. Thermal-dissipation sap flow sensors may not yield consistent sap-flux estimates over multiple years. Trees 2010, 24, 165–174. [Google Scholar] [CrossRef]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Urban, L.; Zhao, P. Granier’s thermal dissipation probe (TDP) method for measuring sap flow in trees: Theory and practice. Aata Bot. Sin.Engl. Ed. 2004, 46, 631–646. [Google Scholar]
- Nadezhdina, N.; Čermák, J.; Ceulemans, R. Radial patterns of sap flow in woody stems of dominant and understory species: Scaling errors associated with positioning of sensors. Tree Physiol. 2002, 22, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Bai, X.-F.; Xu, G.-J.; Zhang, B.-X.; You, G.-C.; Liu, S.; Gu, Y.; Shang, S. Others Trunk sap flux density of Pinus sylvestris var. mongolica in Zhanggutai area. J. Northeast For. Univ. 2013, 41, 27–82. [Google Scholar]
- Bartkowiak, S.M.; Samuelson, L.J.; McGuire, M.A.; Teskey, R.O. Fertilization increases sensitivity of canopy stomatal conductance and transpiration to throughfall reduction in an 8-year-old loblolly pine plantation. For. Ecol. Manag. 2015, 354, 87–96. [Google Scholar] [CrossRef]
- Jiao, S.-R. Structure and Function of Forestry Ecosystem for Sand-Fixation in Zhanggutai Area; Liaoning Science and Technology Press: Shenyang, China, 1989. [Google Scholar]
- Ewers, B.E.; Oren, R. Analyses of assumptions and errors in the calculation of stomatal conductance from sap flux measurements. Tree Physiol. 2000, 20, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- Pichler, P.; Oberhuber, W. Radial growth response of coniferous forest trees in an inner Alpine environment to heat-wave in 2003. For. Ecol. Manag. 2007, 242, 688–699. [Google Scholar] [CrossRef]
- Llorens, P.; Poyatos, R.; Latron, J.; Delgado, J.; Oliveras, I.; Gallart, F. A multi-year study of rainfall and soil water controls on Scots pine transpiration under Mediterranean mountain conditions. Hydrol. Process. 2010, 24, 3053–3064. [Google Scholar] [CrossRef]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guada, G.; Camarero, J.J.; Sánchez-Salguero, R.; Cerrillo, R.M.N. Limited growth recovery after drought-induced forest dieback in very defoliated trees of two pine species. Front. Plant Sci. 2016, 7, 418. [Google Scholar] [CrossRef] [PubMed]
- Eilmann, B.; Dobbertin, M.; Rigling, A. Growth response of Scots pine with different crown transparency status to drought release. Ann. For. Sci. 2013, 70, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Irvine, J.; Perks, M.P.; Magnani, F.; Grace, J. The response of Pinus sylvestris to drought: Stomatal control of transpiration and hydraulic conductance. Tree Physiol. 1998, 18, 393–402. [Google Scholar] [CrossRef]
- Fogel, R. Root turnover and productivity of coniferous forests. In Tree Root Systems and Their Mycorrhizas; Springer-Verlag: New York, NY, USA, 1983; pp. 75–85. [Google Scholar]
- Vanguelova, E.; Nortcliff, S.; Moffat, A.; Kennedy, F. Morphology, biomass and nutrient status of fine roots of Scots pine (Pinus sylvestris) as influenced by seasonal fluctuations in soil moisture and soil solution chemistry. Plant Soil 2005, 270, 233–247. [Google Scholar] [CrossRef]
- Palmroth, S.; Berninger, F.; Nikinmaa, E.; Lloyd, J.; Pulkkinen, P.; Hari, P. Structural adaptation rather than water conservation was observed in Scots pine over a range of wet to dry climates. Oecologia 1999, 121, 302–309. [Google Scholar] [CrossRef]
- Poyatos, R.; Llorens, P.; Piñol, J.; Rubio, C. Response of Scots pine (Pinus sylvestris L.) and pubescent oak (Quercus pubescens Willd.) to soil and atmospheric water deficits under Mediterranean mountain climate. Ann. For. Sci. 2008, 65, 306. [Google Scholar] [CrossRef]
- Morán-López, T.; Poyatos, R.; Llorens, P.; Sabaté, S. Effects of past growth trends and current water use strategies on Scots pine and pubescent oak drought sensitivity. Eur. J. For. Res. 2014, 133, 369–382. [Google Scholar] [CrossRef]
- Wieser, G.; Gruber, A.; Oberhuber, W. Sap flow characteristics and whole-tree water use of Pinus cembra across the treeline ecotone of the central Tyrolean Alps. Eur. J. For. Res. 2014, 133, 287–295. [Google Scholar] [CrossRef]
- Jones, H.G. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 3rd ed.; Cambridge University Press: Cambridge, UK, 2014; pp. 122–152. [Google Scholar]
- Addington, R.N.; Mitchell, R.J.; Oren, R.; Donovan, L.A. Stomatal sensitivity to vapor pressure deficit and its relationship to hydraulic conductance in Pinus palustris. Tree Physiol. 2004, 24, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeppel, M.J.; Murray, B.R.; Barton, C.; Eamus, D. Seasonal responses of xylem sap velocity to VPD and solar radiation during drought in a stand of native trees in temperate Australia. Funct. Plant Biol. 2004, 31, 461–470. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | DBH (cm) | H (m) | Hb (m) | As (cm2) |
|---|---|---|---|---|
| 1 | 14.3 | 9.1 | 4.14 | 127 |
| 2 | 15.1 | 9.8 | 3.89 | 141 |
| 3 | 16.1 | 9.3 | 3.91 | 159 |
| 4 | 16.9 | 8.2 | 3.96 | 175 |
| 5 | 17.7 | 9.6 | 4.00 | 192 |
| 6 | 17.7 | 9.7 | 3.80 | 192 |
| 7 | 17.8 | 9.7 | 4.01 | 194 |
| 8 | 18.1 | 9.7 | 4.01 | 200 |
| 9 | 18.2 | 9.7 | 3.73 | 202 |
| 10 | 18.6 | 9.8 | 4.04 | 211 |
| 11 | 19.1 | 9.9 | 4.09 | 222 |
| Mean | 17.2 | 9.5 | 3.96 | 183 |
| Standard Error (S.E.) | 0.45 | 0.15 | 0.04 | 8.9 |
| Year | Mean Daily Transpiration (kg·tree−1·d−1) | Mean Daily Canopy Stomatal Conductance (cm·s−1) | ||||
|---|---|---|---|---|---|---|
| Early Season 1 | Mid-Season | Late Season | Early Season | Mid-Season | Late Season | |
| 2013 | 22.8 ± 2.3 Aa | 23.9 ± 2.6 Ac | 13.8 ± 1.2 Ba | 0.30 ± 0.03 Aa | 0.39 ± 0.04 Ac | 0.31 ± 0.03 Ab |
| 2014 | 20.8 ± 1.6 Aa | 16.7 ± 1.2 Aa | 11.9 ± 0.9 Bab | 0.26 ± 0.02 Aa | 0.26 ± 0.02 Aa | 0.19 ± 0.01 Ba |
| 2015 | 11.6 ± 0.8 Bb | 8.5 ± 0.5 Ab | 9.2 ± 0.6 Abc | 0.14 ± 0.01 ABb | 0.11 ± 0.01 Bb | 0.16 ± 0.01 Aa |
| 2016 | 12.6 ± 2.3 Ab | 12.6 ± 2.0 Aab | 7.7 ± 1.0 Ac | 0.15 ± 0.03 Ab | 0.18 ± 0.03 Aab | 0.19 ± 0.03 Aa |
| 2017 | 11.5 ± 0.9 Ab | 11.2 ± 0.8 Aab | 7.6 ± 0.6 Bc | 0.13 ± 0.01 Ab | 0.19 ± 0.02 Bab | 0.17 ± 0.01 ABa |
| mean | 16.1 ± 1.01 A | 13.7 ± 1.07 A | 10.4 ± 0.51 B | 0.20 ± 0.01 A | 0.23 ± 0.02 A | 0.20 ± 0.01 A |
| pyears | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| ptrees | 0.29 | 0.53 | 0.24 | 0.35 | 0.15 | 0.51 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, H.; Lu, P.; Yang, W.; Han, H.; Zhang, J. Drought-Induced Reductions and Limited Recovery in the Radial Growth, Transpiration, and Canopy Stomatal Conductance of Mongolian Scots Pine (Pinus sylvestris var. mongolica Litv): A Five-Year Observation. Forests 2019, 10, 1143. https://0-doi-org.brum.beds.ac.uk/10.3390/f10121143
Dang H, Lu P, Yang W, Han H, Zhang J. Drought-Induced Reductions and Limited Recovery in the Radial Growth, Transpiration, and Canopy Stomatal Conductance of Mongolian Scots Pine (Pinus sylvestris var. mongolica Litv): A Five-Year Observation. Forests. 2019; 10(12):1143. https://0-doi-org.brum.beds.ac.uk/10.3390/f10121143
Chicago/Turabian StyleDang, Hongzhong, Ping Lu, Wenbin Yang, Hui Han, and Jun Zhang. 2019. "Drought-Induced Reductions and Limited Recovery in the Radial Growth, Transpiration, and Canopy Stomatal Conductance of Mongolian Scots Pine (Pinus sylvestris var. mongolica Litv): A Five-Year Observation" Forests 10, no. 12: 1143. https://0-doi-org.brum.beds.ac.uk/10.3390/f10121143