Mite Communities (Acari, Mesostigmata) in the Initially Decomposed ‘Litter Islands’ of 11 Tree Species in Scots Pine (Pinus sylvestris L.) Forest


Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Soil Sampling and Litterbag Experiment Design
2.3. Mite Extraction and Identification
2.4. Data Analysis
3. Results
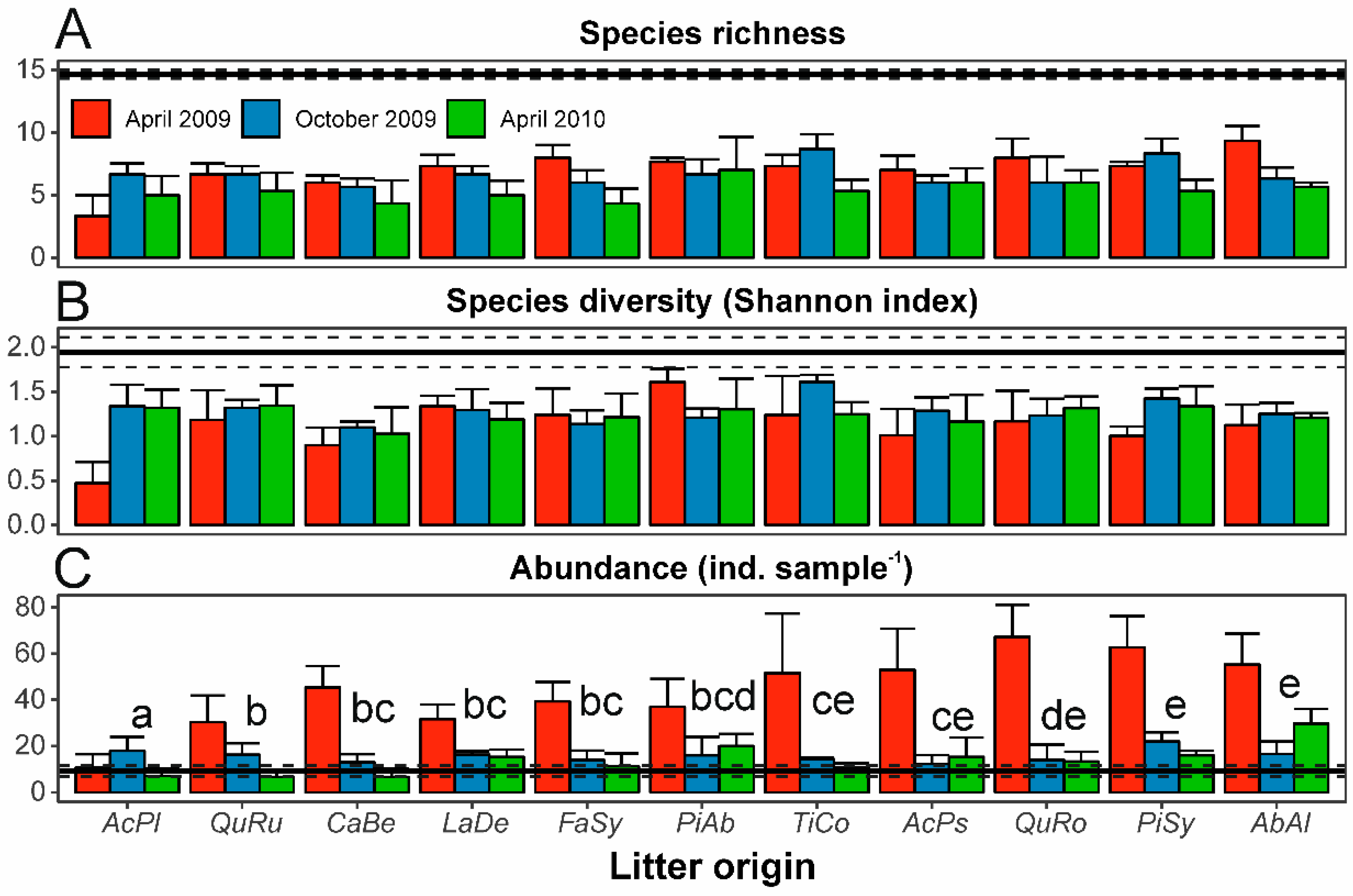
3.1. Mite Abundance, Species Richness, and Diversity
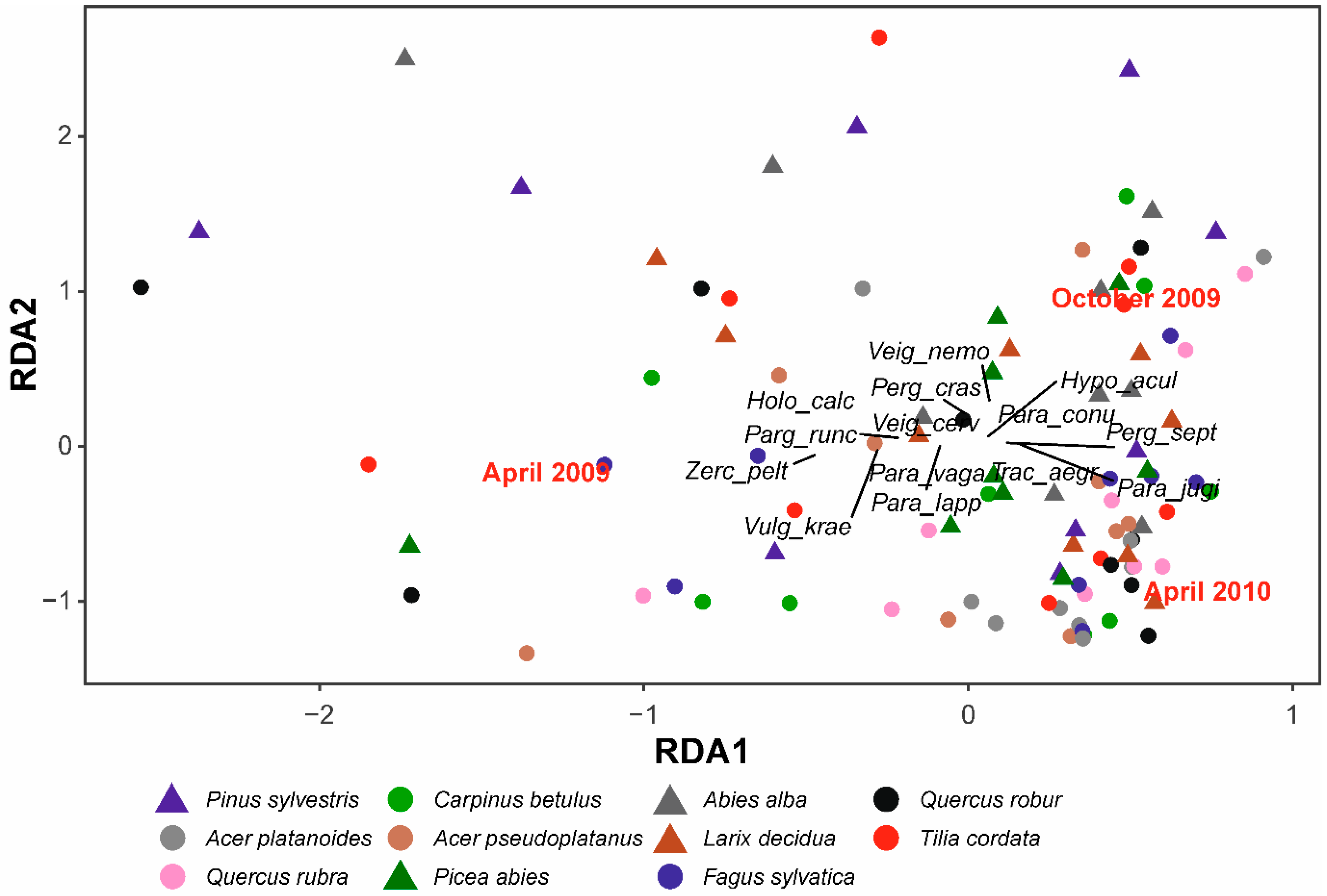
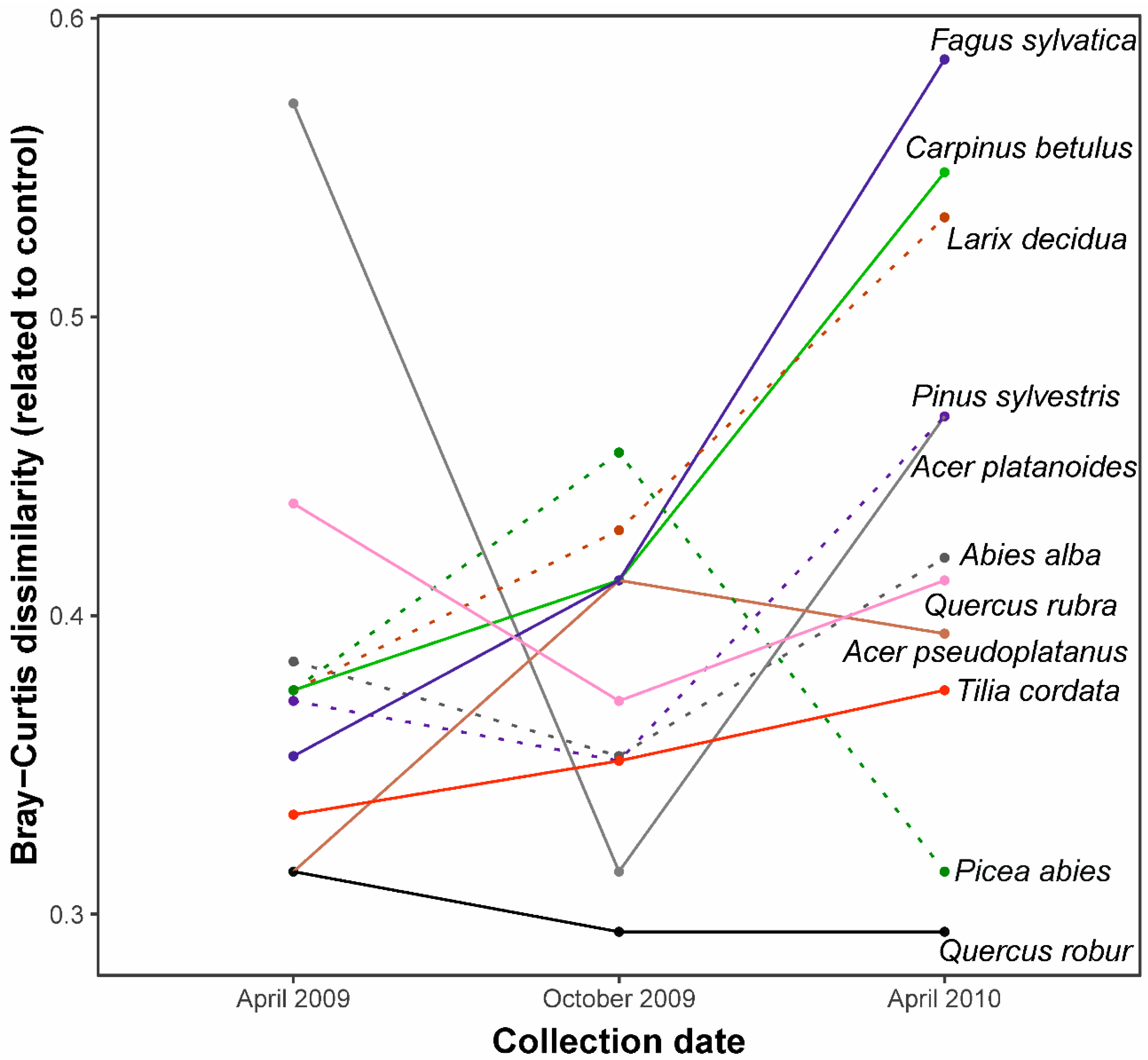
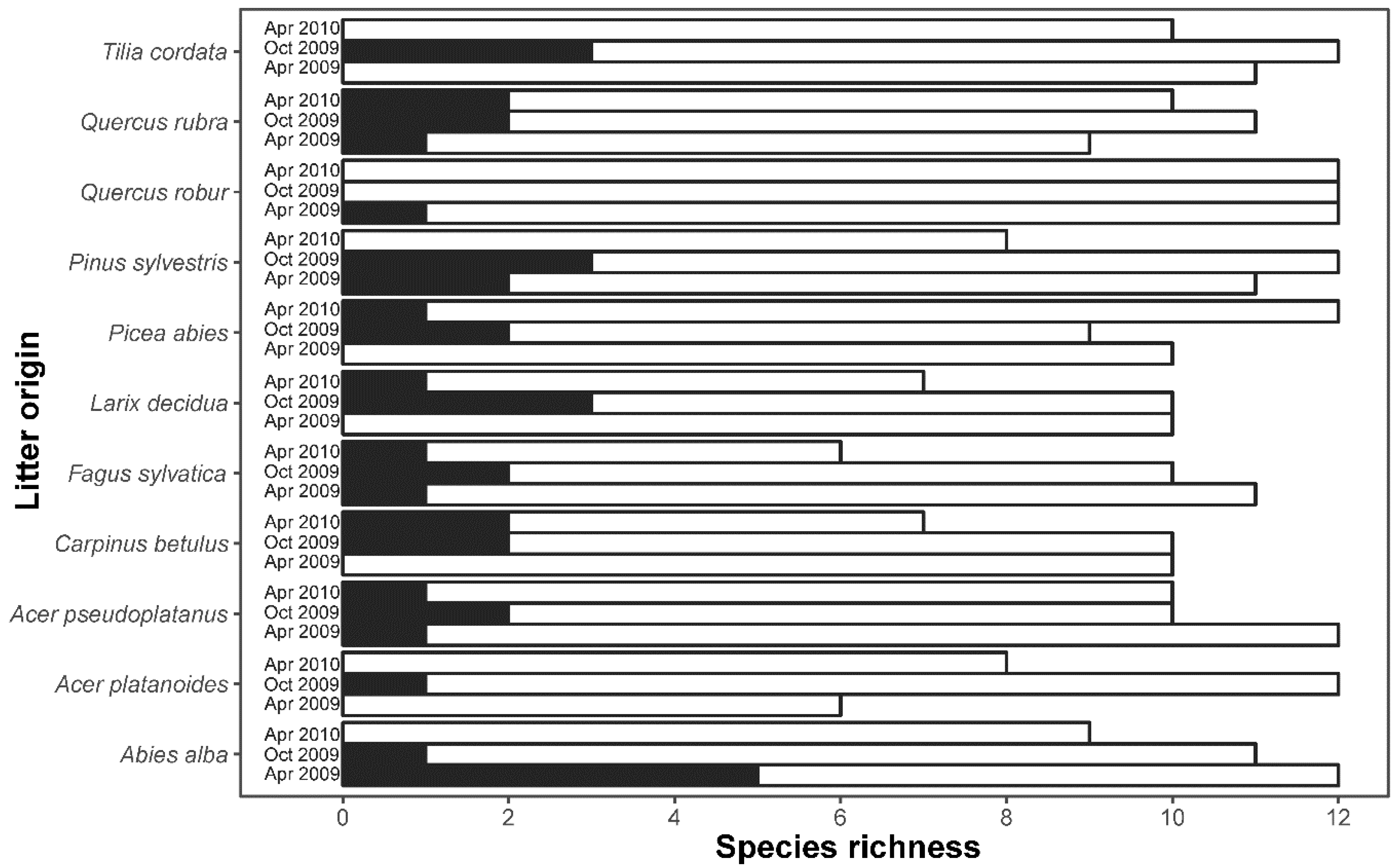
3.2. Similarity between Litterbag and Soil Mite Pool Community
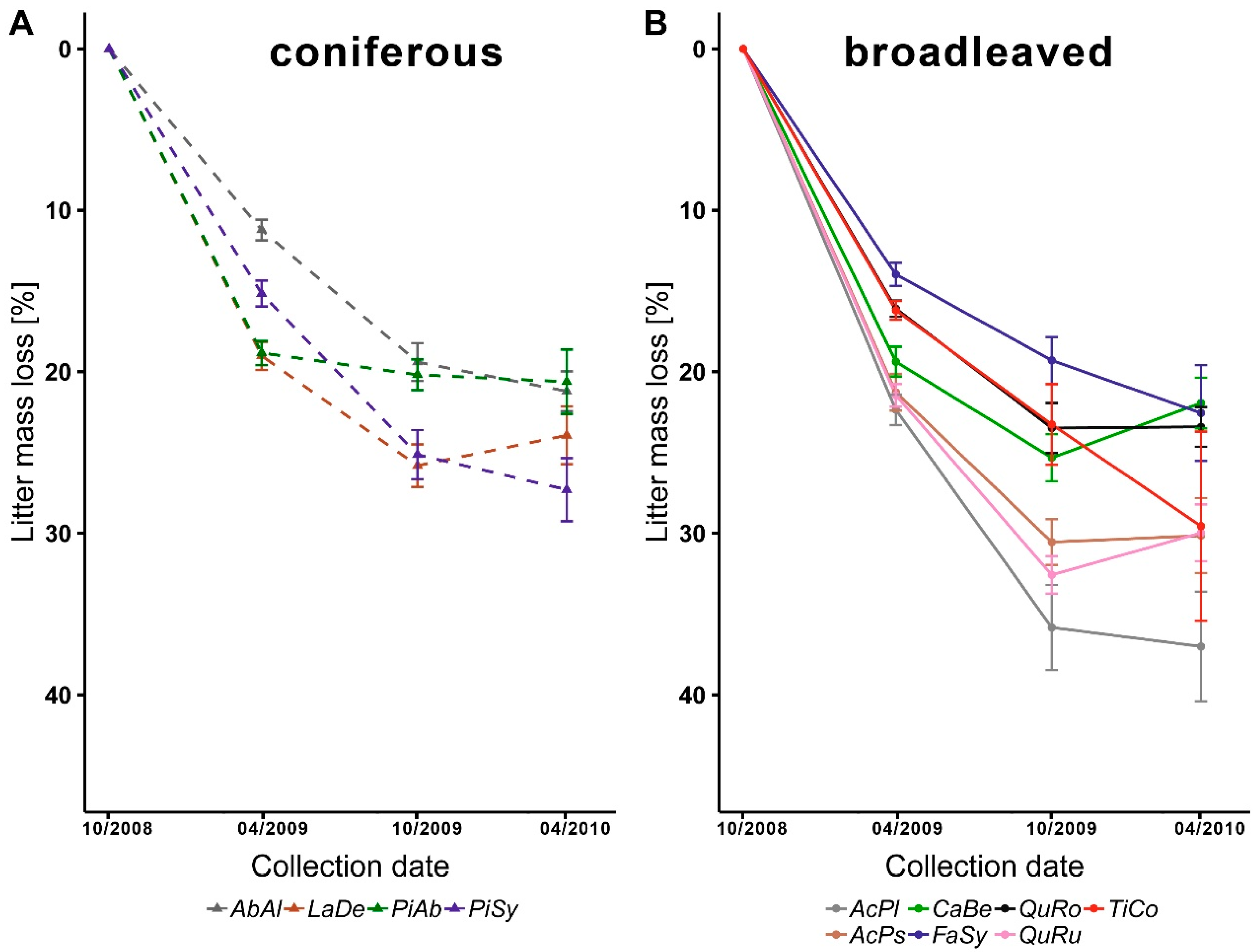
3.3. Litter Decomposition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month and Year | Mean Temperature [°C] | Precipitation Sum [mm] |
|---|---|---|
| October 2008 * | 8.3 | 46.9 |
| November 2008 | 4.9 | 35.7 |
| December 2008 | 0.8 | 25.2 |
| January 2009 | −3.5 | 11.5 |
| February 2009 | −1.3 | 20.1 |
| March 2009 | 3.5 | 14.8 |
| April 2009 ** | 12.0 | 9.4 |
| May 2009 | 13.6 | 96.5 |
| June 2009 | 15.3 | 169.3 |
| July 2009 | 19.4 | 224.6 |
| August 2009 | 19.0 | 84.3 |
| September 2009 | 15.1 | 56.9 |
| October 2009 ** | 6.9 | 146.6 |
| November 2009 | 5.6 | 66.4 |
| December 2009 | −0.8 | 82.3 |
| January 2010 | −6.6 | 61.3 |
| February 2010 | −1.6 | 22.9 |
| March 2010 | 3.4 | 50.7 |
| April 2010 ** | 8.7 | 70.7 |
| No. | Mite Species | Soil Pool | Litterbags with Various Litter Types | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abies alba | Acer platanoides | Acer pseudoplatanus | Carpinus betulus | Fagus sylvatica | Larix decidua | Picea abies | Pinus sylvestris | Quercus robur | Quercus rubra | Tilia cordata | |||
| 1 | Alliphis halleri (G. & R. Canestrini, 1881) | - | - | - | - | - | - | - | - | - | - | + | - |
| 2 | Amblyseius sp. | + | - | - | - | - | - | - | + | - | + | - | - |
| 3 | Arctoseius cetratus (Sellnick, 1940) | - | - | - | - | - | - | - | - | - | - | - | + |
| 4 | Arctoseius semiscissus (Berlese, 1892) | - | - | - | - | - | - | - | - | + | - | - | - |
| 5 | Asca aphidioides (Linnaeus, 1758) | - | + | - | + | + | + | + | - | + | - | + | + |
| 6 | Gamasellodes bicolor Berlese, 1918 | + | + | - | - | - | - | - | + | + | - | + | + |
| 7 | Holoparasitus calcaratus (C.L. Koch, 1839) | ++ | + | + | + | + | + | + | + | + | + | - | + |
| 8 | Hypoaspis aculeifer (Canestrini, 1883) | + | + | + | + | + | + | + | + | + | + | - | + |
| 9 | Hypoaspis praesternalis Willmann, 1949 | - | + | - | - | - | - | - | - | - | - | - | - |
| 10 | Hypoaspis procera Karg 1965 | - | + | - | - | - | - | - | + | + | - | - | - |
| 11 | Hypoaspis vacua (Michael, 1891) | - | + | - | - | + | - | ++ | - | - | - | + | - |
| 12 | Laelapsis astronomica (Koch, 1839) | - | + | - | - | - | - | - | - | - | - | - | - |
| 13 | Oodinychus ovalis C.L. Koch, 1839 | + | - | - | - | + | - | - | - | - | + | + | + |
| 14 | Paragamasus conus Karg, 1971 | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | + | + | ++ |
| 15 | Paragamasus jugincola (Athas-Henriot, 1967) | +++ | +++ | ++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | ++ | ++ |
| 16 | Paragamasus lapponicus Tragardh, 1910 | + | + | + | + | + | + | - | + | + | ++ | - | - |
| 17 | Paragamasus vagabundus (Karg, 1968) | ++ | + | + | + | + | ++ | + | ++ | + | ++ | + | + |
| 18 | Pargamasus runcatellus (Berlese, 1903) | +++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| 19 | Pergamasus crassipes Linnaeus, 1758 | + | + | + | + | + | + | + | + | + | + | - | + |
| 20 | Pergamasus mediocris Berlese, 1904 | + | - | - | - | - | - | - | - | - | - | - | + |
| 21 | Pergamasus septentrionalis Oudemans, 1902 | - | + | + | + | + | + | + | + | + | + | + | + |
| 22 | Rhodacarellus silesiacus Willmann, 1936 | + | - | - | - | - | - | - | - | + | - | - | - |
| 23 | Rhodacarus coronatus Berlese, 1921 | ++ | - | + | - | - | - | + | - | - | + | + | + |
| 24 | Trachytes aegrota (C. L. Koch, 1841) | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | + |
| 25 | Trichouropoda obscura (C.L. Koch, 1836) | + | - | - | - | - | - | - | - | - | - | - | - |
| 26 | Veigaia cervus (Kramer, 1876) | ++ | + | + | + | - | ++ | + | + | + | + | + | - |
| 27 | Veigaia nemorensis (C. L. Koch, 1839) | ++ | +++ | ++ | ++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ | +++ |
| 28 | Vulgarogamasus kraepelini (Berlese, 1904) | + | +++ | + | ++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | ++ |
| 29 | Zercon peltatus C.L. Koch, 1836 | ++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ |
| 30 | Zercon triangularis C.L. Koch, 1836 | + | - | - | - | + | + | - | - | - | + | + | + |
| 31 | Zercon zelawaiensis Sellnick, 1944 | + | - | - | - | - | - | + | - | - | - | - | - |
References
- Vanbergen, A.J.; Woodcock, B.A.; Watt, A.D.; Niemela, J. Effect of land-use heterogeneity on carabid communities at the landscape scale. Ecography 2005, 28, 3–16. [Google Scholar] [CrossRef]
- Hunter, M.L. Maintaining Biodiversity in Forest Ecosystems; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Young, R.; Giese, R. Introduction to Forest Ecosystem Science and Management, 3rd ed.; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Götmark, F.; Friedman, J.; Kempe, G.; Nord, B. Broadleaved tree species in conifer-dominated forestry: Regeneration and limitation of saplings in southern Sweden. For. Ecol. Manag. 2005, 214, 142–157. [Google Scholar] [CrossRef]
- Alem, S.; Pavlis, J.; Urban, J.; Kucera, J. Pure and mixed plantations of Eucalyptus camaldulensis and Cupressus lusitanica: Their growth interactions and effect on diversity and density of undergrowth woody plants in relation to light. Open J. For. 2015, 5, 375. [Google Scholar]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, 1–13. [Google Scholar] [CrossRef]
- Migge, S.; Maraun, M.; Scheu, S.; Schaefer, M. The oribatid mite community (Acarina) of pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) of different age. Appl. Soil Ecol. 1998, 9, 115–121. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, H.; Wang, B. Effects of soil microarthropods on plant litter decomposition across an elevation gradient in the Wuyi Mountains. Soil Biol. Biochem. 2009, 41, 891–897. [Google Scholar] [CrossRef]
- Fujii, S.; Takeda, H. Succession of soil microarthropod communities during the aboveground and belowground litter decomposition processes. Soil Biol. Biochem. 2017, 110, 95–102. [Google Scholar]
- Franca, M.; Sandmann, D.; Krashevska, V.; Maraun, M.; Scheu, S. Altitude and decomposition stage rather than litter origin structure soil microarthropod communities in tropical montane rainforests. Soil Biol. Biochem. 2018, 263–274. [Google Scholar]
- Urbanowski, C.K.; Horodecki, P.; Kamczyc, J.; Skorupski, M.; Jagodziński, A.M. Succession of mite assemblages (Acari, Mesostigmata) during decomposition of tree leaves in forest stands growing on reclaimed post-mining spoil heap and adjacent forest habitats. Forests 2018, 9, 718. [Google Scholar] [CrossRef]
- Petersen, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 288–388. [Google Scholar] [CrossRef]
- Ruf, A.; Beck, L. The use of predatory soil mites in ecological soil classification and assessment concepts, with perspectives for oribatid mites. Ecotoxicol. Environ. Saf. 2005, 62, 290–299. [Google Scholar] [CrossRef]
- Koehler, H.H. Predatory mites (Gamasina, Mesostigmata). Agric. Ecosyst. Environ. 1999, 74, 395–410. [Google Scholar] [CrossRef]
- Gulvik, M. Mites (Acari) as indicators of soil biodiversity and land use monitoring: A review. Pol. J. Ecol. 2007, 415–440. [Google Scholar]
- Gan, H.; Zak, D.R.; Hunter, M.D. Chronic nitrogen deposition alters the structure and function of detrital food webs in a northern hardwood ecosystem. Ecol. Appl. 2013, 23, 1311–1321. [Google Scholar] [CrossRef]
- Berg, M.P.; Kniese, J.P.; Bedaux, J.J.M.; Verhoef, H.A. Dynamics and stratification of functional groups of micro- and mesoarthropods in the organic layer of a Scots pine forest. Biol. Fertil. Soils 1998, 26, 268–284. [Google Scholar] [CrossRef]
- Magcale-Macandog, D.B.; Manlubatan, M.B.T.; Javier, J.M.; Edrial, J.D.; Mago, K.S.; De Luna, J.E.I.; Nayoos, J.; Porcioncula, R.P. Leaf litter decomposition and diversity of arthropod decomposers in tropical Muyong forest in Banaue, Philippines. Paddy Water Environ. 2018, 16, 265–277. [Google Scholar] [CrossRef]
- Vasconcelos, H.L.; Laurance, W.F. Influence of habitat, litter type, and soil invertebrates on leaf-litter decomposition in a fragmented Amazonian landscape. Oecologia 2005, 144, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Elkins, N.Z.; Whitford, W.G. The role of microarthropods and nematodes in decomposition in a semi-arid ecosystem. Oecologia 1982, 55, 303–310. [Google Scholar] [CrossRef]
- González, G.; Seastedt, T.R.; Donato, Z. Earthworms, arthropods and plant litter decomposition in aspen (Populus tremuloides) and lodgepole pine (Pinus contorta) forests in Colorado, USA. Pedobiologia 2003, 47, 863–869. [Google Scholar] [CrossRef]
- Heneghan, L.; Coleman, D.C.; Zou, X.; Crossley, D.A.; Haines, B.L. Soil microarthropod contributions to decomposition dynamics: Tropical–temperate comparisons of a single substrate. Ecology 1999, 80, 1873–1882. [Google Scholar]
- Lindo, Z.; Winchester, N.N. Oribatid mite communities and foliar litter decomposition in canopy suspended soils and forest floor habitats of western redcedar forests, Vancouver Island, Canada. Soil Biol. Biochem. 2007, 39, 2957–2966. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I.; et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.; Crossley, D.A.; Callaham, L.C. Effects of litter quality and microarthropods on N dynamics and retention of exogenous 15N in decomposing litter. Biol. Fertil. Soils 1992, 12, 241–252. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A. Nutrients in forest litter treated with naphthalene and simulated throughfall: A field microcosm study. Soil Biol. Biochem. 1983, 15, 159–165. [Google Scholar] [CrossRef]
- Horodecki, P.; Nowiński, M.; Jagodziński, A.M. Advantages of mixed tree stands in restoration of upper soil layers on postmining sites: A five-year leaf litter decomposition experiment. Land Degrad. Dev. 2019, 30, 3–13. [Google Scholar] [CrossRef]
- Gholz, H.L. Long-term dynamics of pine and hardwood litter in contrasting environments: Toward a global model of decomposition. Glob. Change Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Freschet, G.T.; Aerts, R.; Cornelissen, J.H.C. Multiple mechanisms for trait effects on litter decomposition: Moving beyond home-field advantage with a new hypothesis. J. Ecol. 2012, 100, 619–630. [Google Scholar] [CrossRef]
- Keiser, A.D.; Strickland, M.S.; Fierer, N.; Bradford, M.A. The effect of resource history on the functioning of soil microbial communities is maintained across time. Biogeosciences 2011, 8, 1477–1486. [Google Scholar] [CrossRef] [Green Version]
- Gergócs, V.; Hufnagel, L. The effect of microarthropods on litter decomposition depends on litter quality. Eur. J. Soil Biol. 2016, 75, 24–30. [Google Scholar] [CrossRef]
- Ceitel, J. Zmiany mikroklimatu przygruntowej warstwy powietrza oraz morfologii drzew ze wzrostem upraw sosnowych założonych w różnych więźbach początkowych; Katedra Hodowli Lasu, Akademia Rolnicza w Poznaniu. Ph.D. Thesis, Department of Silviculture, Agricultural University, Poznań, Poland, 1982. [Google Scholar]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Oleksyn, J. Ecological consequences of silviculture at variable stand densities. I. Stand growth and development. Sylwan 2009, 153, 75–85. [Google Scholar]
- Jagodziński, A.M.; Oleksyn, J. Ecological consequences of silviculture at variable stand densities. II. Biomass production and allocation, nutrient retention. Sylwan 2009, 153, 147–157. [Google Scholar]
- Jagodziński, A.M.; Oleksyn, J. Ecological consequences of silviculture at variable stand densities. III. Stand stability, phytoclimate and biodiversity. Sylwan 2009, 153, 219–230. [Google Scholar]
- Skorupski, M.; Dalik, J.; Ceitel, J. Species composition of mites from the order Mesostigmata in experimental pine and spruce tree stands planted in various initial spacing. Sci. Pap. Agric. Univ. Poznan For. 2003, 49–56. [Google Scholar]
- Crossley, D.A.; Blair, J.M. Proceedings of the International Workshop on Modern Techniques in Soil Ecology Relevant to Organic Matter Breakdown, Nutrient Cycling and Soil Biological Processes A high-efficiency, “low-technology” Tullgren-type extractor for soil microarthropods. Agric. Ecosyst. Environ. 1991, 34, 187–192. [Google Scholar] [CrossRef]
- Edwards, C.A. The assessment of populations of soil-inhabiting invertebrates. Agric. Ecosyst. Environ. 1991, 34, 145–176. [Google Scholar] [CrossRef]
- Karg, W. Acari (Acarina), Milben Parasitiformes (Anactinochaeta), Cohors Gamasina Leach. Raubmilben. Die Tierwelt Deutschlands; VEB Gustav Fischer Verlag: Jena, Germany, 1993.
- Ghilarov, M.C.; Bregetova, N.G. Opredelitel obitajuscich v Pocve Klescej-Mesostigmata (Key to the Soil Mites—Mesostigmata); Nauka: Leningrad, Russia, 1977. [Google Scholar]
- Micherdziński, W. Die Familie Parasitidae Oudemans, 1901 “(Acarina, Mesostigmata)”; Państwowe Wydawnictwo Naukowe: Kraków, Poland, 1969. [Google Scholar]
- Oksanen, J.; Blanchet, F.; Legendre, P.; Michin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Henry, M.; Stevens, H.; Wagner, H. “vegan” 2.3.3.–Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 8 May 2019).
- Wieder, R.K.; Lang, G.E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 14 March 2019).
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Seastedt, T.R. The role of microarthropods in decomposition and mineralization processes. Annu. Rev. Entomol. 1984, 29, 25–46. [Google Scholar] [CrossRef]
- Kaneko, N.; Salamanca, E. Mixed leaf litter effects on decomposition rates and soil microarthropod communities in an oak–pine stand in Japan. Ecol. Res. 1999, 14, 131–138. [Google Scholar] [CrossRef]
- González, G.; Seastedt, T.R. Comparison of the abundance and composition of litter fauna in tropical and subalpine forests. Pedobiologia 2000, 44, 545–555. [Google Scholar] [CrossRef]
- Gergócs, V.; Rétháti, G.; Hufnagel, L. Litter quality indirectly influences community composition, reproductive mode and trophic structure of oribatid mite communities: A microcosm. Exp. Appl. Acarol. 2015, 67, 335–356. [Google Scholar] [CrossRef]
- Kampichler, C.; Bruckner, A. The role of microarthropods in terrestrial decomposition: A meta-analysis of 40 years of litterbag studies. Biol. Rev. 2009, 84, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Usher, M.B. Studies on a wood-feeding termite community in Ghana, West Africa. Biotropica 1975, 7, 217–233. [Google Scholar] [CrossRef]
- Minor, M.A.; Cianciolo, J.M. Diversity of soil mites (Acari: Oribatida, Mesostigmata) along a gradient of land use types in New York. Appl. Soil Ecol. 2007, 35, 140–153. [Google Scholar] [CrossRef]
- Walter, D.E.; Proctor, H. Mites: Ecology, Evolution & Behaviour–Life at a Microscale|David Evans Walter, Springer; CABI Publishing: New York, NY, USA, 1999. [Google Scholar]
- Martikainen, E.; Huhta, V. Interactions between nematodes and predatory mites in raw humus soil: A microcosm experiment. Rev. DÉcologie Biol. Sol 1990, 27, 13–20. [Google Scholar]
- Hasegawa, M.; Okabe, K.; Fukuyama, K.; Makino, S.; Okochi, I.; Tanaka, H.; Goto, H.; Mizoguchi, T.; Sakata, T. Community structures of Mesostigmata, Prostigmata and Oribatida in broad-leaved regeneration forests and conifer plantations of various ages. Exp. Appl. Acarol. 2013, 59, 391–408. [Google Scholar] [CrossRef] [PubMed]





| Response | Term | Estimate | SE | z | Pr (>|z|) |
|---|---|---|---|---|---|
| Abundance | (Intercept) | 4.0945 | 0.0770 | 53.1450 | <0.0001 |
| Litter origin—Acer platanoides | −1.0376 | 0.1125 | −9.2250 | <0.0001 | |
| Litter origin—Quercus rubra | −0.6353 | 0.0978 | −6.4980 | <0.0001 | |
| Litter origin—Carpinus betulus | −0.4374 | 0.0918 | −4.7630 | <0.0001 | |
| Litter origin—Acer pseudoplatanus | −0.2215 | 0.0863 | −2.5680 | 0.0102 | |
| Litter origin—Picea abies | −0.3214 | 0.0887 | −3.6220 | 0.0003 | |
| Litter origin—Abies alba | 0.0099 | 0.0812 | 0.1220 | 0.9031 | |
| Litter origin—Larix decidua | −0.4634 | 0.0926 | −5.0060 | <0.0001 | |
| Litter origin—Fagus sylvatica | −0.4426 | 0.0920 | −4.8110 | <0.0001 | |
| Litter origin—Quercus robur | −0.0615 | 0.0826 | −0.7440 | 0.4571 | |
| Litter origin—Tilia cordata | −0.2680 | 0.0874 | −3.0670 | 0.0022 | |
| Date—October 2009 | −1.0295 | 0.0511 | −20.1360 | <0.0001 | |
| Date—April 2010 | −1.1567 | 0.0536 | −21.5720 | <0.0001 | |
| Random effect—plot | Variance | 0.0070 | SD | 0.0839 | |
| Richness | (Intercept) | 2.2331 | 0.2102 | 10.6240 | <0.0001 |
| Decomposition constant (k) | −1.1424 | 0.7384 | −1.5470 | 0.1218 | |
| Date—October 2009 | −0.0572 | 0.0937 | −0.6100 | 0.5420 | |
| Date—April 2010 | −0.2735 | 0.0994 | −2.7520 | 0.0059 | |
| Random effect—plot | Variance | 0.0197 | SD | 0.1405 | |
| Shannon | (Intercept) | 1.3827 | 0.1961 | 7.0500 | <0.0001 |
| Decomposition constant (k) | −0.6634 | 0.5996 | −1.1060 | 0.2710 | |
| Random effect—plot | Variance | 0.0444 | SD | 0.2107 |
| Term | df | Variance | F | Pr (>F) |
|---|---|---|---|---|
| Collection date | 2 | 0.06917 | 5.4696 | 0.001 |
| Residual | 96 | 0.60690 | - | - |
| AIC | −44.43 | AIC0 | −37.75 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamczyc, J.; Dyderski, M.K.; Horodecki, P.; Jagodziński, A.M. Mite Communities (Acari, Mesostigmata) in the Initially Decomposed ‘Litter Islands’ of 11 Tree Species in Scots Pine (Pinus sylvestris L.) Forest. Forests 2019, 10, 403. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050403
Kamczyc J, Dyderski MK, Horodecki P, Jagodziński AM. Mite Communities (Acari, Mesostigmata) in the Initially Decomposed ‘Litter Islands’ of 11 Tree Species in Scots Pine (Pinus sylvestris L.) Forest. Forests. 2019; 10(5):403. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050403
Chicago/Turabian StyleKamczyc, Jacek, Marcin K. Dyderski, Paweł Horodecki, and Andrzej M. Jagodziński. 2019. "Mite Communities (Acari, Mesostigmata) in the Initially Decomposed ‘Litter Islands’ of 11 Tree Species in Scots Pine (Pinus sylvestris L.) Forest" Forests 10, no. 5: 403. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050403