1. Introduction
The more than 500 extant species of oak (
Quercus spp.) are widely distributed across the Northern Hemisphere, including Mesoamerica [
1,
2]. Oak-dominated forests are of great ecological and economical importance [
3,
4]. For instance, oaks are extensively used in soil and water conservation and restoration efforts since they have strong, adventitious root systems and exhibit a high tolerance to waterlogging [
5,
6,
7]. Additionally, oaks are well adapted to fire and hence are frequently used for constructing forest fire belts [
8]. Oaks also play an important role in maintaining biodiversity. For instance, Caprio and Ellena [
9] reported that the retention of native oaks is the key factor for the conservation of winter bird diversity in local deciduous woods. In addition to the ecological functions, oak timber is distinguished for its great strength, durability and beauty [
10]. As a high-grade material, oak is widely used in shipbuilding and in the manufacture of furniture, sports equipment and flooring [
11].
China contains approximately 51 oak species, which are widely distributed throughout the country’s mountainous regions [
12,
13]. According to the 8th Chinese National Forest Inventory (CNFI), oak-dominated forests cover 16.72 million hm
2, accounting for 10.15% of the total forest area. Stocking is 1.294 billion m
3, representing 8.76% of the total stocking in China [
14]. A total of 96.29% of the oak-dominated forests in China are degraded secondary natural forests. They have historically experienced extensive disturbance, and many are almost coppice forests of very poor quality. For instance, the average stocking per hectare of oak-dominated forests in China is about 77.39 m
3, while in Germany it is 305 m
3 per hectare [
15]. Therefore, it is urgent to manage these degraded forests in a sustainable way to improve their ecological and economic value.
The prediction of future stand development under different management scenarios is of great importance to inform forest management. Forest growth and yield models are an effective tool to provide such information [
16,
17]. Forest growth and yield models can be divided into three types, namely whole-stand growth models, size-class models and individual-tree models [
18,
19,
20,
21]. Almost all oak-dominated forests are uneven-aged mixed-species forests, and more detailed information is required to formulate management strategies for them. Under these circumstances, a whole-stand growth model, which is specifically produced for single-tree species plantations, is not appropriate [
18]. Although size class models have been documented to model forest dynamics for uneven-aged mixed-species forests, they cannot provide the information required for individual trees. In China, close-to-nature forest management with a single tree selective cutting system has been widely adopted for the management of uneven-aged mixed-species forests [
22]. This single-tree selective cutting system requires predictive information at the level of the individual trees. Fortunately, individual-tree models, which use single trees as the basic modeling unit, have the special ability to predict individual tree growth under a complex combination of species mixtures, stand structures, and silvicultural practices [
23,
24,
25].
As an important component of individual-tree models, individual-tree basal area increment models are generally expressed as a linear function of tree size, competition pressures, and site conditions [
26,
27,
28,
29]. In China, individual-tree basal area increment models have been developed for many tree species, e.g., red pine (
Pinus koraiensis Sieb. et Zucc.) [
30], China fir (
Cunninghamia lanceolata (Lamb.) Hook.) [
31], and linden (
Tilia tuan Szyszyl.) [
32]. Unfortunately, there is no individual-tree basal area increment model for oak species in China. Additionally, forest structural diversity, an important part of biological diversity, has been extensively documented to affect forest growth and productivity [
33,
34]. For instance, Lei et al. [
35] selected tree species, tree size, and height diversity indices to represent structural diversity and found that forest structural diversity had a significant positive effect on net growth and survivor growth. However, only a few studies have included forest structural diversity in forest growth and yield models.
The traditional regression analysis using ordinary least square regression (OLS) is the most commonly used method to develop individual-tree models [
36,
37]. In principle, the successful use of OLS requires that the data should satisfy the following statistical assumptions: independence of observations, and normally distributed residuals with equal variance of the residuals [
38,
39,
40]. However, forestry data usually has the characteristic of a hierarchical stochastic structure because of the repeated measurements of the same sampling units and the nested structure of the sampling units [
41,
42]. This feature of forestry data violates these assumptions and using OLS in such a situation could result in biased estimations [
42,
43].
Mixed-effects models provide an efficient means to deal with longitudinal and nested data [
44,
45]. They contain both fixed effects parameters and random effects parameters. Fixed effects parameters account for covariate or treatment effects as in traditional regression, while random effects parameters explain the different sources of stochastic variability [
46,
47,
48]. Mixed-effects models are therefore extensively used in forestry, such as diameter–height models [
49,
50], crown models [
51,
52], self-thinning models [
53,
54,
55], and growth models [
56,
57].
In this study, the main objective was to develop a linear mixed-effects individual-tree basal area increment model for oaks in a degraded natural secondary forest in Hunan Province, south-central China. We hypothesized that the introduction of forest structural diversity and random effects would significantly improve model performance and we also hope our model will contribute to forest management strategies by predicting forest dynamics under different management scenarios.
4. Discussion
In this study, we developed a linear mixed-effects individual-tree basal area increment model for oaks considering forest structural diversity in Hunan Province, south-central China, using data from 845 sample plots of CNFI measured three times each. The model was described as a stochastic process, where the fixed component explained the mean value for the basal area increment and the random part incorporated unexplained residual variability acting at the level of sample plot (e.g., moisture, soil parameters, nutrient content, etc.) [
47,
63]. Our final mixed-effects model included the variables 1/DBH, RD, NT, EL and GC in the fixed component. Additionally, the relative importance values for 1/DBH, RD, NT, EL and GC were 33.15%, 38.78%, 13.94%, 12.71% and 1.42%, respectively. From the perspective of forest management, the independent variables that have higher relative importance value should be given higher priority in forest management planning.
Consistent with results reported by Cao et al. [
73], Uzoh and Oliver [
56], Lei et al. [
74], Pokharel and Dech [
59], and Timilsina and Staudhammer [
75], 1/DBH was significantly negatively related to basal area increment. It had the second largest effect on basal area increment. RD, a distance-independent individual-tree-level competition index, was found to be positively related to basal area increment, which had more effect on basal area increment than any other variable. Similar findings were documented by Lei et al. [
74] and Yan [
32]. Both 1/DBH and RD were indicators of individual tree competition status in a forest stand and their relationship with basal area increment suggested that larger trees have stronger competitive ability for resources, especially for light which is considered as the major limiting resource for individual-tree diameter growth [
65,
76]. For instance, Ma et al. [
77] reported the nutrient use efficiency of the China fir in mature stands was almost twice that in young stands. RD has frequently been selected to represent individual tree competition effects due to its performance in models [
74,
78]. In comparison to the individual-tree-level competition indices, i.e., 1/DBH and RD, NT can be thought to represent stand-level competition, which had the third largest effect on basal area increment. In our study, a negative relationship was observed between NT and basal area increment, indicating that stand-level competition reduces individual tree growth.
There are two different types of competition, i.e., one- and two-sided competition [
18,
79]. In one-sided competition, larger trees are not affected by their smaller neighbors. In two-sided competition, by contrast, resources are shared by all trees either equally or proportionally to size [
80]. It is commonly assumed that one-sided competition is driven by the availability of aboveground resources, whereas two-sided competition is more reflective of belowground competition [
81]. Therefore, many growth and yield models consider both one- and two-sided indices of competition to more comprehensively quantify the level of competition experienced by a tree, as well as its social position within the stand [
18,
79]. Thus, we also considered one-sided (BAL, BAL/DBH, RD) and two-sided (NT, BA) competition when developing the basic model in the present study. RD and NT were included in the model to represent the comprehensive effects of aboveground and belowground competition.
Although we included three site-level independent variables, i.e., EL, SLSin, and SLCos, only EL was left in our final model. EL had the fourth largest effect on basal area growth. We observed a negative correlation between EL and basal area increment, which suggested that individual trees exhibited faster growth in the lower elevation. This negative correlation could be attributed to the shorter growing season at higher elevations [
60,
65,
82]. Additionally, many authors argued that EL could indirectly affect tree growth by altering temperature, moisture, light, soil nutrient availability and other chemical and physical agents in a forest stand [
83,
84].
GC had the smallest influence on basal area increment. The negative correlation between GC and basal area increment suggested that variation in large tree size reduces basal area increment. Similar results were also reported by Liang et al. [
34], Cordonnier and Kunstler [
85] and Bourdier et al. [
86]. Bourdier et al. [
86] attributed the negative tree size inequality effect to the reduced total light interception of the stand, which reduces light use efficiency. Their detailed explanations are as follows: for stands with a comparable basal area and mean diameter, the increase in GC indicates greater difference in the DBHs between the biggest and small trees. Because tree canopy width and depth increase asymptotically with tree size, the light interception efficiency, which is determined by the canopy characteristics, also increases asymptotically. Therefore, a decrease in the amount of light intercepted might occur when the gain in light intercepted by the bigger trees is unlikely to compensate for the loss of light intercepted by the smaller trees. In comparison, the reduced light use efficiency with increases in the GC may be attributed to the fact that large individuals already intercept more light and an increase in light only slightly improves their growth, whereas small individuals live in low light conditions where supplementary light has a stronger effect on growth. Therefore, from a forest management perspective, extremely large trees, which have significant relative dominance and absolute advantages in light competition, should be cut to reduce tree-size inequality in a stand. Consequently, the total light interception and light use efficiency of a stand might increase.
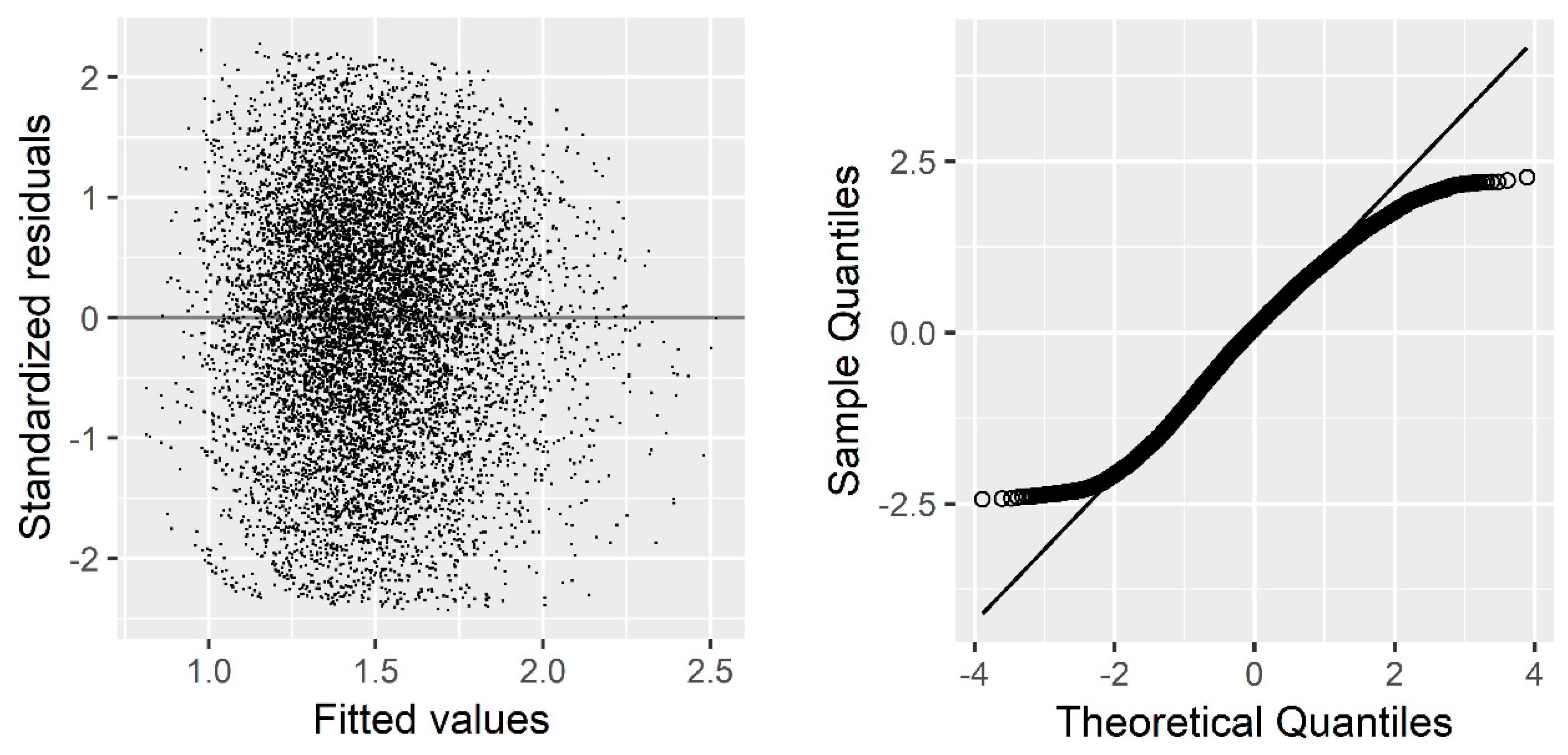
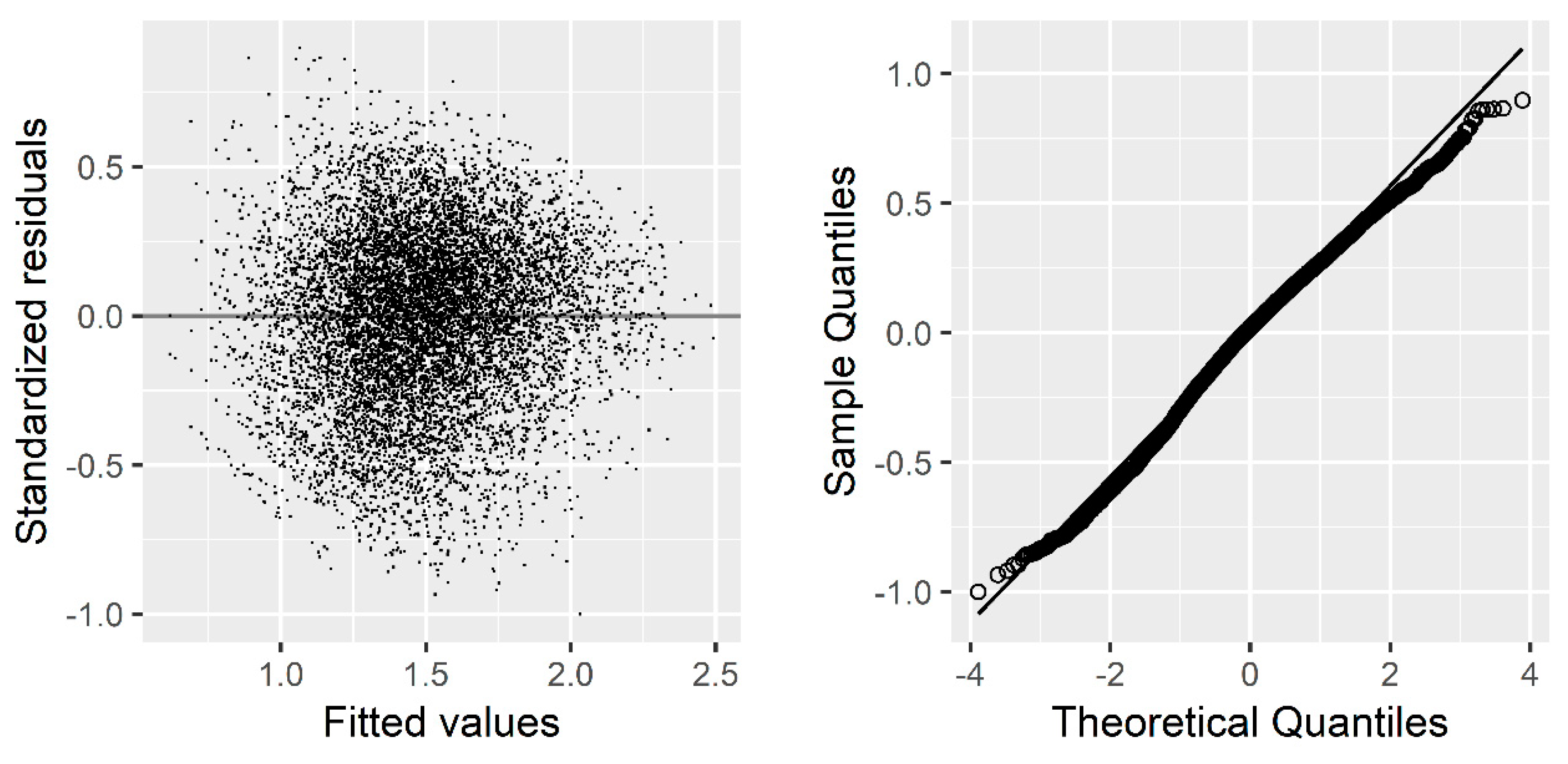
Due to the hierarchical structural of our data, we used the sample plot as a random effect to produce the individual-tree basal area increment model. Our results indicated a significant improvement in model performance after introducing the random effects. For example, the AIC dropped from 6317.446 to 4705.766, and BIC from 6367.733 to 4878.177. Similar results have been extensively documented by other authors [
65,
67,
75].
Although introducing random effects could correct or reduce autocorrelation and heteroscedasticity [
65,
68,
87,
88], our mixed-effects model still exhibited autocorrelation and heteroscedasticity. We therefore further introduced three variance functions and three correlation structures to refine our model. Finally, based on the lack-of-fit statistics, the exponent function and AR(1) were determined as the optimum option for correcting the heteroscedasticity and autocorrelation of the residuals. Similar results were also reported by Calama and Montero [
47] and Fabian et al. [
56]. Additionally, the model prediction and evaluation using validation data further supported the conclusion that the mixed-effects approach had significantly improved the model’s predictive performance, either in PA or SS in comparison to the basic model without random effects. For instance, the Bias decreased from 0.2239 to 0.1716, the RMSE decreased from 0.2776 to 0.2167 and the
R2 increased from 0.3663 to 0.6140.
Although the model performance was improved after we used the mixed-effects method to develop the individual-tree basal area increment model for oaks, there are some limitations of our final mixed-effects model. Firstly, we only considered the autocorrelation structure to correct autocorrelation. However, Zhao et al [
65] reported that modelling the autocorrelation structure alone could not completely match the possible common effects and suggested introducing random period effect to describe autocorrelation, if data is sufficient. Pokharel and Dech [
59] directly introduced random period effects to develop their diameter growth model. In the present study, unfortunately only three period observations were available for the same trees and hence it makes no sense to employ period as a random effect.
Secondly, climatic change has been widely reported to influence forest growth, productivity, tree species composition and distribution [
89,
90,
91]. For instance, Battles et al. [
92] assessed the impact of climate change on the productivity and health of the mixed-conifer forest in California and found that the productivity of stem volume increment was reduced by 19% in the most extreme climate change. Unfortunately, we did not include climatic variables due to lack of data. It is imperative to integrate climatic variables into forest growth models when data is available.
There are many individual-tree forest simulators throughout the world, for instance, the FVS, TASS, PROGNAUS in the US, the SILVA, and BWINPro in Germany and CAPSIS in France [
93,
94,
95]. For a comprehensive prediction of forest dynamics, this individual-tree simulation system not only included individual-tree basal area or diameter increment models, but also integrated an individual-tree mortality and recruitment model. Therefore, the development of an individual-tree mortality and recruitment model of oak species is strongly recommended in the future.







{kind=link}
{kind=link}
{kind=link}
{kind=link}