Effects of Phosphate-Solubilizing Bacteria and N2-fixing Bacteria on Nutrient Uptake, Plant Growth, and Bioactive Compound Accumulation in Cyclocarya paliurus (Batal.) Iljinskaja


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seedlings, Growth Media, and Microorganism’s Preparation
2.2. Site Description and Experimental Design
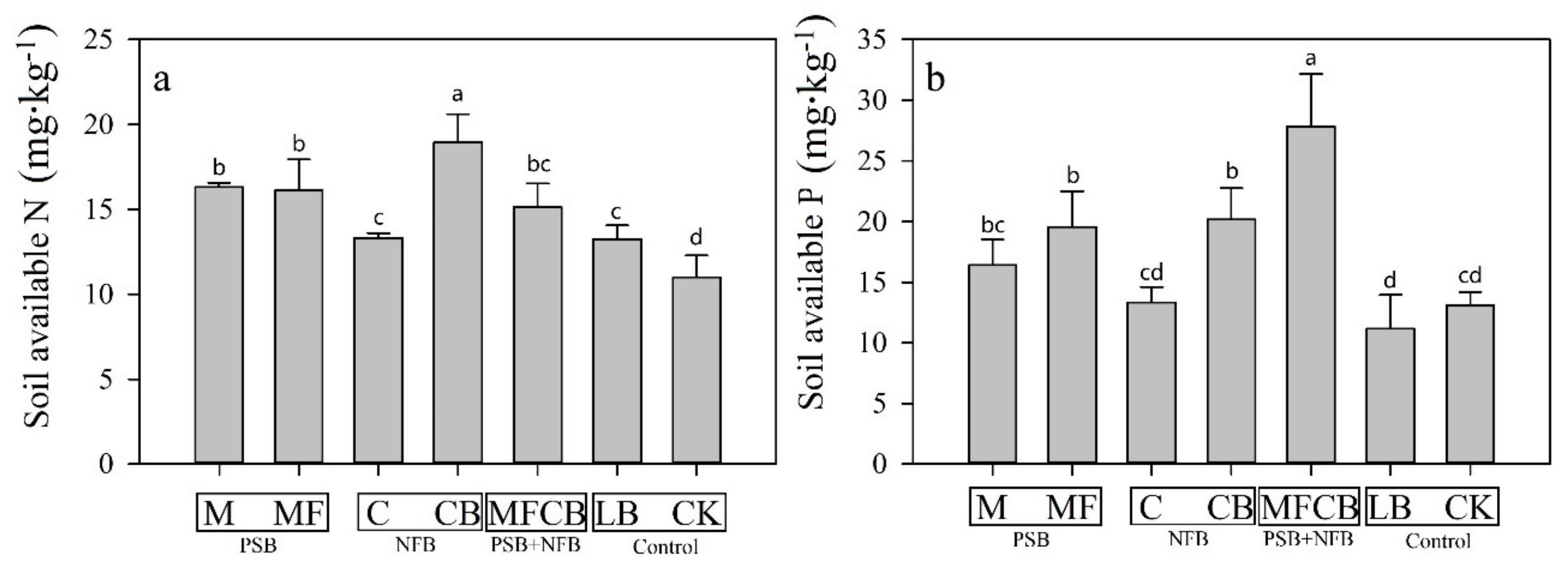
2.3. Measurement of Soil Available Nitrogen and Phosphorus
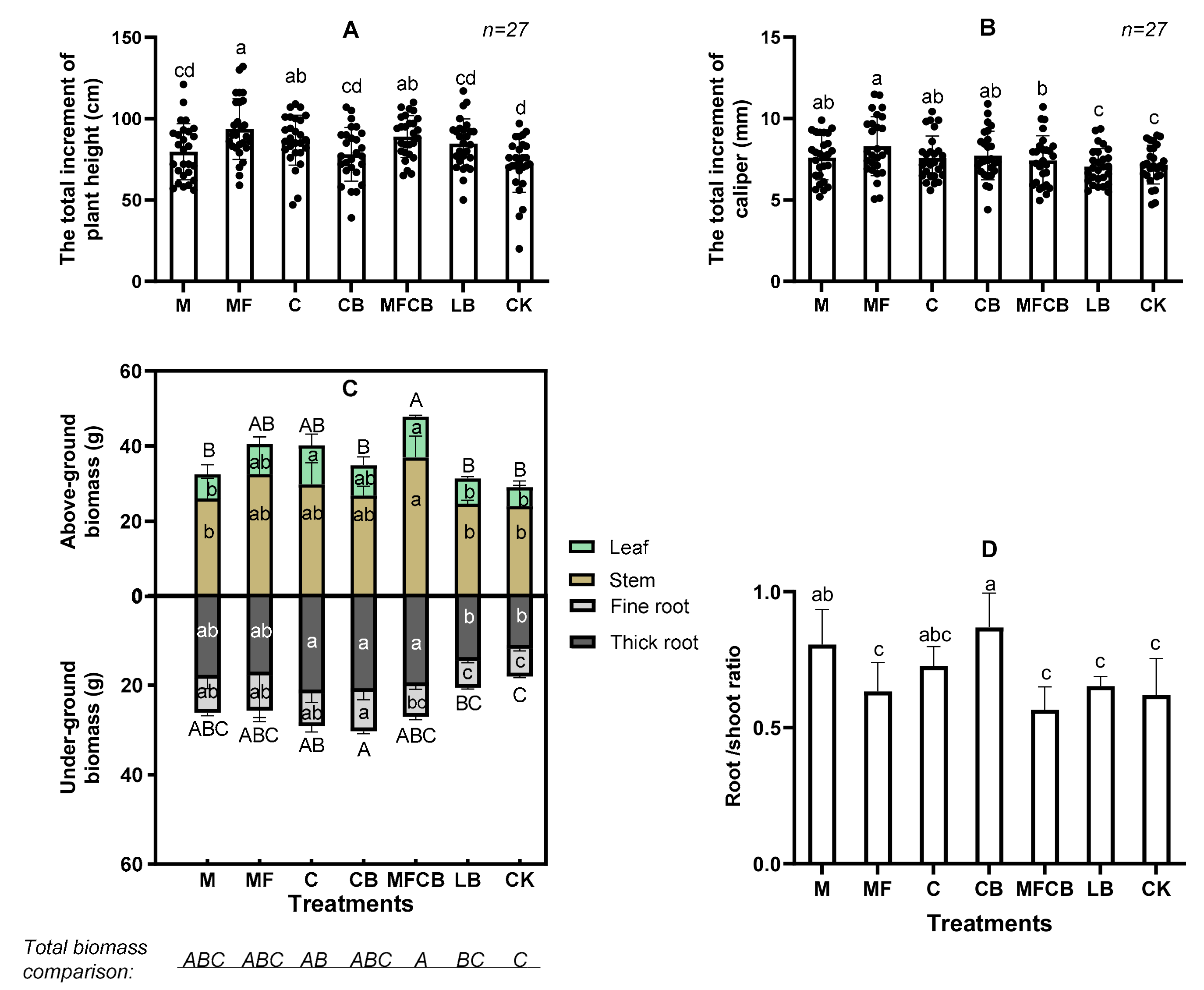
2.4. Plant Growth and Leaf Harvest
2.5. Measurement of Total Carbon, Nitrogen, and Phosphorus in Leaves
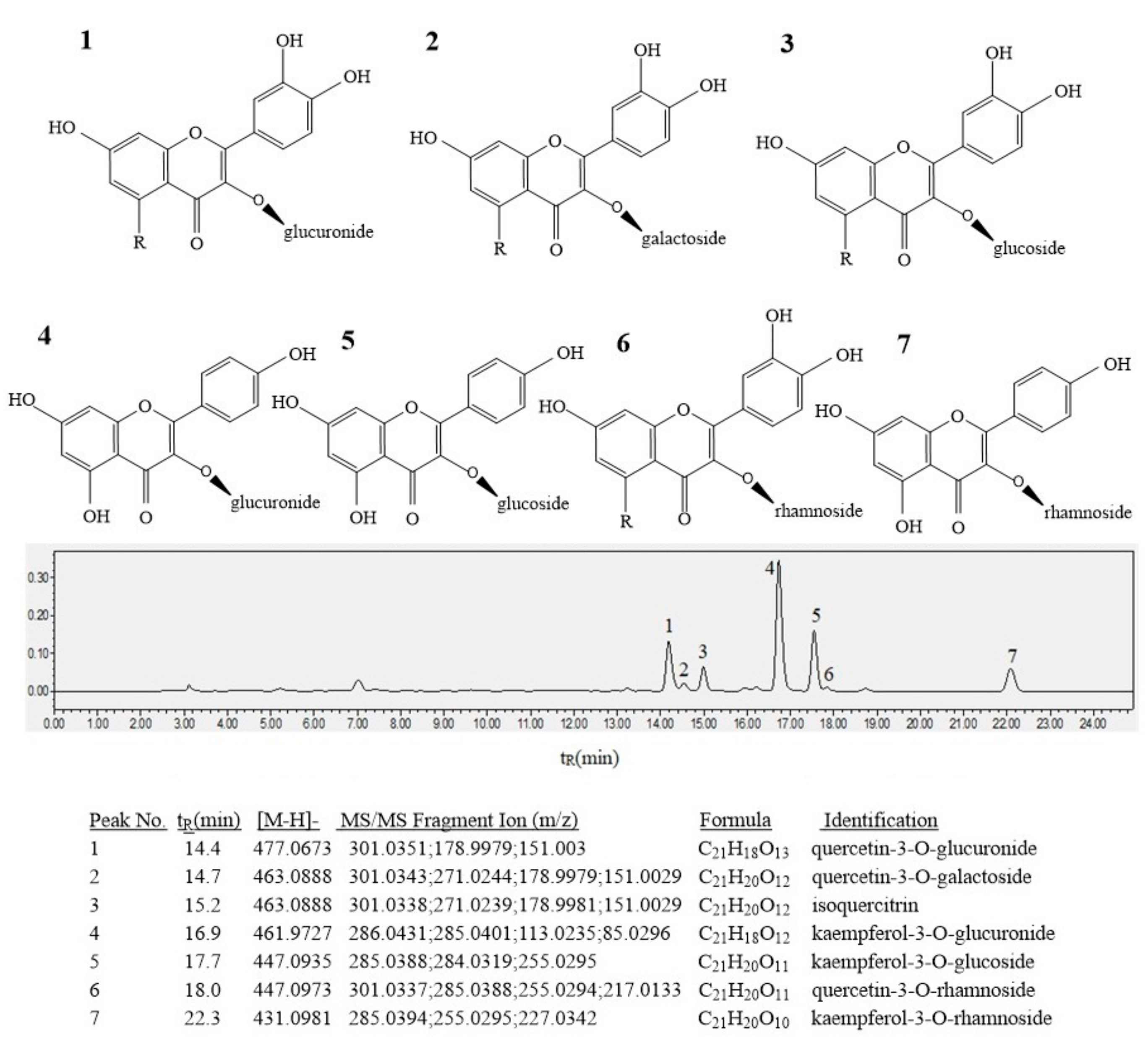
2.6. Extraction and Determination of Bioactive Compounds
2.7. Statistical Analysis
3. Results
3.1. Soil Available N and P
3.2. Plant Growth and Biomass
3.3. C:N:P Stoichiometry in C. paliurus Leaves
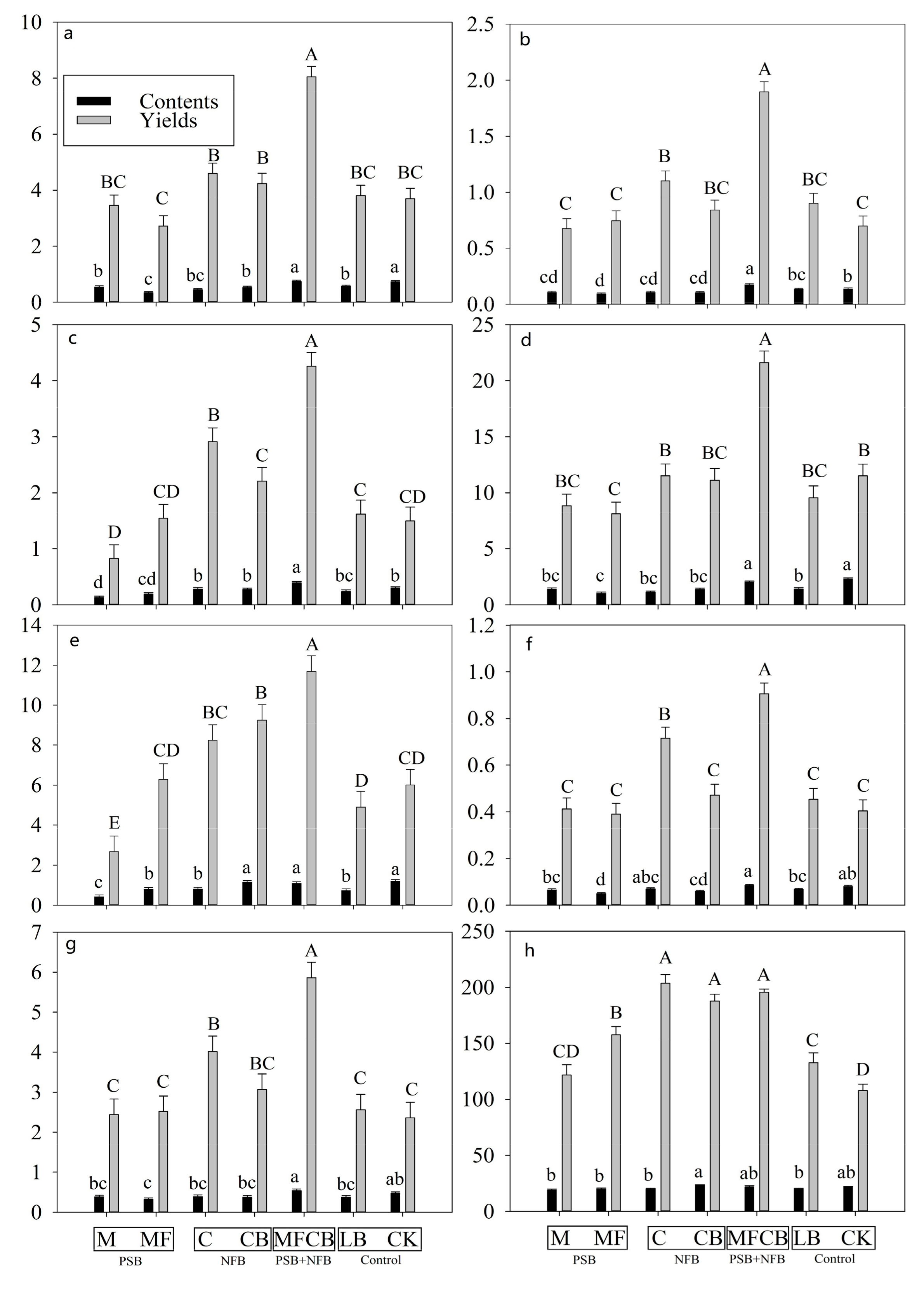
3.4. Flavonoids
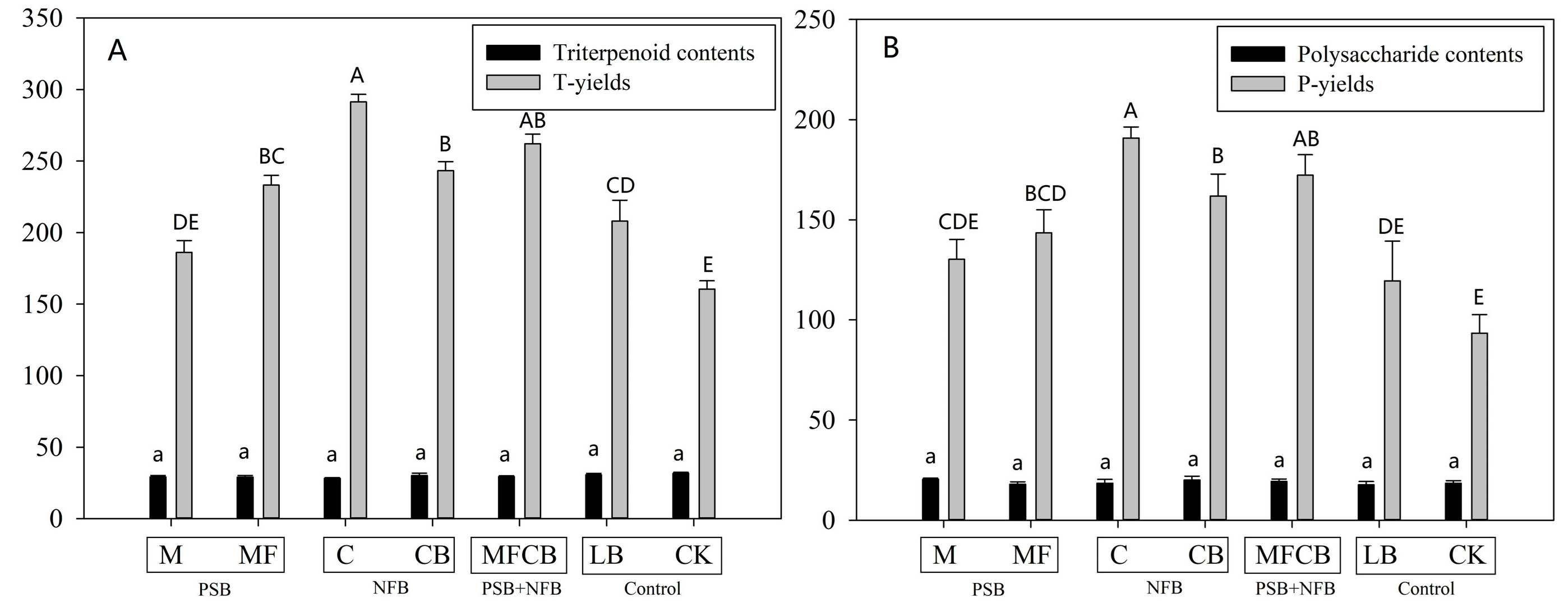
3.5. Water-Soluble Polysaccharide and Triterpenoid
4. Discussion
4.1. PGPR Symbiosis Increased Nutrients in Soil Which Improved Plant Growth
4.2. PGPR Additions Improved the BC Output Mainly by Increasing the Leaf Biomass rather than Concentrations
4.3. Selections of PGPR Could be Considered for Multiple Purposes of C. paliurus Plantation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fang, S.Z.; Yang, W.X.; Chu, X.L.; Shang, X.L.; She, C.Q.; Fu, X.X. Provenance and temporal variations in selected flavonoids in leaves of Cyclocarya paliurus. Food Chem. 2011, 124, 1382–1386. [Google Scholar] [CrossRef]
- Kennelly, E.J.; Cai, L.; Long, L.; Shamon, L.; Zaw, K.; Zhou, B.N.; Pezzuto, J.M.; Kinghorn, A.D. Novel Highly Sweet Secodammarane Glycosides from Pterocarya-Paliurus. J. Agric. Food Chem. 1995, 43, 2602–2607. [Google Scholar] [CrossRef]
- Xie, J.H.; Dong, C.J.; Nie, S.P.; Li, F.; Wang, Z.J.; Shen, M.Y.; Xie, M.Y. Extraction, chemical composition and antioxidant activity of flavonoids from Cyclocarya paliurus (Batal.) Iljinskaja leaves. Food Chem. 2015, 186, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shen, Q.; Lu, J.C.; Li, J.Y.; Liu, W.Y.; Yang, J.J.; Li, J.; Xiao, K. Phenolic compounds from the leaves of Cyclocarya paliurus (Batal.) Ijinskaja and their inhibitory activity against PTP1B. Food Chem. 2010, 119, 1491–1496. [Google Scholar] [CrossRef]
- Wu, Z.F.; Meng, F.C.; Cao, L.J.; Jiang, C.H.; Zhao, M.G.; Shang, X.L.; Fang, S.Z.; Ye, W.C.; Zhang, Q.W.; Zhang, J.; et al. Triterpenoids from Cyclocarya paliurus and their inhibitory effect on the secretion of apoliprotein B48 in Caco-2 cells. Phytochemistry 2017, 142, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Q.; Chang, S.; Clinton, P.W.; Johnson, C.E.; Wang, H.L.; Xu, Z.H. Preface to the special issue for the 8th International Symposium on Forest Soils: Linking Soil Processes to Forest Productivity and Water Protection under Global Change. J. Soil Sediment 2017, 17, 2215–2217. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Z.; Sun, J.; Ren, C.J.; Kang, D.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 2015, 5, 10195. [Google Scholar]
- Blaise, D.; Singh, J.V.; Bonde, A.N.; Tekale, K.U.; Mayee, C.D. Effects of farmyard manure and fertilizers on yield, fibre quality and nutrient balance of rainfed cotton (Gossypium hirsutum). Bioresour. Technol. 2005, 96, 345–349. [Google Scholar] [CrossRef]
- Deng, B.; Shang, X.L.; Fang, S.Z.; Li, Q.Q.; Fu, X.X.; Su, J. Integrated Effects of Light Intensity and Fertilization on Growth and Flavonoid Accumulation in Cyclocarya paliurus. J. Agric. Food Chem. 2012, 60, 6286–6292. [Google Scholar] [CrossRef]
- Kumar, R.; Meena, S.S.; Kakani, R.K.; Mehta, R.S.; Meena, N.K. Response of crop geometry, fertilizer levels and genotypes on productivity of fenugreek (Trigonella foenum-graecum L.). Int. J. Seed Spices 2015, 5, 63–67. [Google Scholar]
- Gyaneshwar, P.; Kumar, G.N.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Kaur, G.; Reddy, M.S. Effects of Phosphate-Solubilizing Bacteria, Rock Phosphate and Chemical Fertilizers on Maize-Wheat Cropping Cycle and Economics. Pedosphere 2015, 25, 428–437. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Kizilkaya, R. Yield response and nitrogen concentrations of spring wheat (Triticum aestivum) inoculated with Azotobacter Chroococcum strains. Ecol. Eng. 2008, 33, 150–156. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; You, Y.H.; Joo, G.J.; Lee, I.J.; Lee, K.E.; Kim, J.H. Phosphate Solubilizing Bacillus megaterium mj1212 Regulates Endogenous Plant Carbohydrates and Amino Acids Contents to Promote Mustard Plant Growth. Indian J. Microbiol. 2014, 54, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Dadrasan, M.; Chaichi, M.R.; Pourbabaee, A.A.; Yazdani, D.; Keshavarz-Afshar, R. Deficit irrigation and biological fertilizer influence on yield and trigonelline production of fenugreek. Ind. Crop. Prod. 2015, 77, 156–162. [Google Scholar] [CrossRef]
- Xie, W.; Hao, Z.P.; Zhou, X.F.; Jiang, X.L.; Xu, L.J.; Wu, S.L.; Zhao, A.H.; Zhang, X.; Chen, B.D. Arbuscular mycorrhiza facilitates the accumulation of glycyrrhizin and liquiritin in Glycyrrhiza uralensis under drought stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef]
- Yu, X.; Liu, X.; Zhu, T.H.; Liu, G.H.; Mao, C. Co-inoculation with phosphate-solubilzing and nitrogen-fixing bacteria on solubilization of rock phosphate and their effect on growth promotion and nutrient uptake by walnut. Eur. J. Soil Biol. 2012, 50, 112–117. [Google Scholar] [CrossRef]
- Cao, Y.N.; Fang, S.Z.; Yin, Z.Q.; Fu, X.X.; Shang, X.L.; Yang, W.X.; Yang, H.M. Chemical Fingerprint and Multicomponent Quantitative Analysis for the Quality Evaluation of Cyclocarya paliurus Leaves by HPLC-Q-TOF-MS. Molecules 2017, 22, 1927. [Google Scholar] [CrossRef]
- Rajendran, P.; Rengarajan, T.; Nandakumar, N.; Palaniswami, R.; Nishigaki, Y.; Nishigaki, I. Kaempferol, a potential cytostatic and cure for inflammatory disorders. Eur. J. Med. Chem. 2014, 86, 103–112. [Google Scholar] [CrossRef]
- Spencer, J.P.E.; El Mohsen, M.M.A.; Rice-Evans, C. Cellular uptake and metabolism of flavonoids and their metabolites: implications for their bioactivity. Arch. Biochem. Biophys. 2004, 423, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fang, S.Z.; Zhou, M.M.; Shang, X.L.; Yang, W.X.; Fu, X.X. Geographic variation in water-soluble polysaccharide content and antioxidant activities of Cyclocarya paliurus leaves. Ind. Crop. Prod. 2018, 121, 180–186. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Fu, X. Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency. Int. J. Environ. Res. Public Health 2019, 16, 2442. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, X.; Zhu, T.H.; Liu, G.H.; Mao, C. Isolation and characterization of phosphate-solubilizing bacteria from walnut and their effect on growth and phosphorus mobilization. Biol. Fert. Soils 2011, 47, 437–446. [Google Scholar] [CrossRef]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: a greenhouse trial. Geoderma 2005, 125, 155–166. [Google Scholar] [CrossRef]
- Kim, C.; Kecskes, M.L.; Deaker, R.J.; Gilchrist, K.; New, P.B.; Kennedy, I.R.; Kim, S.; Sa, T. Wheat root colonization and nitrogenase activity by Azospirillum isolates from crop plants in Korea. Can. J. Microbiol. 2005, 51, 948–956. [Google Scholar] [CrossRef] [Green Version]
- Sanders, E.R. Aseptic Laboratory Techniques: Plating Methods. J. Vis. Exp. 2012, 63, e3064. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Misc. Pap. Inst. Agric. Res. 1954, 939, 1–19. [Google Scholar]
- Bao, J.; Cai, Y.; Sun, M.; Wang, G.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef]
- Fu, X.X.; Zhou, X.D.; Deng, B.; Shang, X.L.; Fang, S.Z. Seasonal and genotypic variation of water-soluble polysaccharide content in leaves of Cyclocarya paliurus. South For. 2015, 77, 231–236. [Google Scholar] [CrossRef]
- Fan, J.P.; He, C.H. Simultaneous quantification of three major bioactive triterpene acids in the leaves of Diospyros kaki by high-performance liquid chromatography method. J. Pharm. Biomed. Anal. 2006, 41, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Fang, S.Z.; Shang, X.L.; Fu, X.X.; Li, Y. Influence of provenance and shade on biomass production and triterpenoid accumulation in Cyclocarya paliurus. Agrofor. Syst. 2019, 93, 483–492. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.; Yang, W.; Shang, X.; Fu, X. Light quality affects flavonoid production and related gene expression in Cyclocarya paliurus. J. Photochem. Photobiol. B 2018, 179, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Li, Y.Y.; Lei, G.; Liu, G.H. Effects of nitrogen availability on mineral nutrient balance and flavonoid accumulation in Cyclocarya paliurus. Plant Physiol. Biochem. 2019, 135, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Megali, L.; Glauser, G.; Rasmann, S. Fertilization with beneficial microorganisms decreases tomato defenses against insect pests. Agron. Sustain. Dev. 2014, 34, 649–656. [Google Scholar] [CrossRef]
- Singh, S.; Kapoor, K.K. Effects of inoculation of phosphate-solubilizing microorganisms and an arbuscular mycorrhizal fungus on mungbean grown under natural soil conditions. Mycorrhiza 1998, 7, 249–253. [Google Scholar] [CrossRef]
- Gull, M.; Hafeez, F.Y.; Saleem, M.; Malik, K.A. Phosphorus uptake and growth promotion of chickpea by co-inoculation of mineral phosphate solubilising bacteria and a mixed rhizobial culture. Aust. J. Exp. Agric. 2004, 44, 623–628. [Google Scholar] [CrossRef]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.F.; Peng, Z.P.; Zeng, X.T.; Houx, J.H. Effects of nitrogen-phosphorus imbalance on plant biomass production: a global perspective. Plant Soil 2019, 436, 245–252. [Google Scholar] [CrossRef]
- Gigolashvili, T.; Kopriva, S. Transporters in plant sulfur metabolism. Front. Plant Sci. 2014, 5, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, F.M.; Canas, R.A.; de la Torre, F.N.; Pascual, M.B.; Castro-Rodriguez, V.; Avila, C. Nitrogen Metabolism and Biomass Production in Forest Trees. Front. Plant Sci. 2018, 9, 1449. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Hao, Z.; Yu, M.; Wu, Z.; Zhao, A.; Li, J.; Zhang, X.; Chen, B. Improved phosphorus nutrition by arbuscular mycorrhizal symbiosis as a key factor facilitating glycyrrhizin and liquiritin accumulation in Glycyrrhiza uralensis. Plant Soil 2018, 439, 243–257. [Google Scholar] [CrossRef]
- Dastmalchi, M.; Chapman, P.; Yu, J.; Austin, R.S.; Dhaubhadel, S. Transcriptomic evidence for the control of soybean root isoflavonoid content by regulation of overlapping phenylpropanoid pathways. BMC Genom. 2017, 18, 70. [Google Scholar] [CrossRef]
- Salla, T.D.; da Silva, R.; Astarita, L.V.; Santarem, E.R. Streptomyces rhizobacteria modulate the secondary metabolism of Eucalyptus plants. Plant Physiol. Biochem. 2014, 85, 14–20. [Google Scholar] [CrossRef]
- Iwashina, T.; Kitajima, J. Chalcone and flavonol glycosides from Asarum canadense (Aristolochiaceae). Phytochemistry 2000, 55, 971–974. [Google Scholar] [CrossRef]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Genotypic and climatic influences on the concentration and composition of flavonoids in kale (Brassica oleracea var. sabellica). Food Chem. 2010, 119, 1293–1299. [Google Scholar] [CrossRef]
- Tavarini, S.; Passera, B.; Martini, A.; Avio, L.; Sbrana, C.; Giovannetti, M.; Angelini, L.G. Plant growth, steviol glycosides and nutrient uptake as affected by arbuscular mycorrhizal fungi and phosphorous fertilization in Stevia rebaudiana Bert. Ind. Crop. Prod. 2018, 111, 899–907. [Google Scholar] [CrossRef]
- Cai, Z.Q.; Wang, W.H.; Yang, J.; Cai, C.T. Growth, photosynthesis and root reserpine concentrations of two Rauvolfia species in response to a light gradient. Ind. Crop. Prod. 2009, 30, 220–226. [Google Scholar] [CrossRef]
- Afshar, R.K.; Chaichi, M.R.; Jovini, M.A.; Jahanzad, E.; Hashemi, M. Accumulation of silymarin in milk thistle seeds under drought stress. Planta 2015, 242, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Islam, M.T. Co-inoculation with Enterobacter and Rhizobacteria on Yield and Nutrient Uptake by Wheat (Triticum aestivum L.) in the Alluvial Soil Under Indo-Gangetic Plain of India. J. Plant Growth Regul. 2017, 36, 608–617. [Google Scholar] [CrossRef]
- Shanmugam, S.R.; Chaganti, S.R.; Lalman, J.A.; Heath, D.D. Effect of inhibitors on hydrogen consumption and microbial population dynamics in mixed anaerobic cultures. Int. J. Hydrog. Energ. 2014, 39, 249–257. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inoculant Type | Treatment | M: Bacillus megaterium | F: Pseudomonas fluorescens | C: Azotobacter chroococcum | B: Azospirillum brasilence | LB | Water |
|---|---|---|---|---|---|---|---|
| PSB 1 | M | 50 | 0 | 0 | 0 | 0 | 0 |
| MF | 25 | 25 | 0 | 0 | 0 | 0 | |
| NFB 2 | C | 0 | 0 | 50 | 0 | 0 | 0 |
| CB | 0 | 0 | 25 | 25 | 0 | 0 | |
| PSB + NFB 3 | MFCB | 12.5 | 12.5 | 12.5 | 12.5 | 0 | 0 |
| Control 4 | LB | 0 | 0 | 0 | 0 | 50 | 0 |
| CK | 0 | 0 | 0 | 0 | 0 | 50 |
| Inoculant Type | Treatment | Carbon (g·kg−1) | Nitrogen (g·kg−1) | Phosphorus (g·kg−1) | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|
| PSB | M | 464.28a 1 | 23.04d | 1.86b | 20.18b | 250.14cd | 12.41d |
| MF | 455.90a | 23.82cd | 1.76c | 19.16bc | 258.50bc | 13.5ab | |
| NFB | C | 464.19a | 25.83b | 1.91ab | 17.97c | 243.42de | 13.54ab |
| CB | 459.50a | 25.08bc | 1.73c | 18.33c | 265.69b | 14.50a | |
| PSB + NFB | MFCB | 464.23a | 27.81a | 1.95a | 16.70d | 237.63e | 14.23ab |
| Control | LB | 465.78a | 21.44e | 1.62d | 21.73a | 287.04a | 13.23cd |
| CK | 462.08a | 21.00e | 1.57d | 22.00a | 295.26a | 13.42cd |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Chen, Z.; Xu, Z.; Fu, X. Effects of Phosphate-Solubilizing Bacteria and N2-fixing Bacteria on Nutrient Uptake, Plant Growth, and Bioactive Compound Accumulation in Cyclocarya paliurus (Batal.) Iljinskaja. Forests 2019, 10, 772. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090772
Wang Z, Chen Z, Xu Z, Fu X. Effects of Phosphate-Solubilizing Bacteria and N2-fixing Bacteria on Nutrient Uptake, Plant Growth, and Bioactive Compound Accumulation in Cyclocarya paliurus (Batal.) Iljinskaja. Forests. 2019; 10(9):772. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090772
Chicago/Turabian StyleWang, Zhikang, Ziyun Chen, Ziheng Xu, and Xiangxiang Fu. 2019. "Effects of Phosphate-Solubilizing Bacteria and N2-fixing Bacteria on Nutrient Uptake, Plant Growth, and Bioactive Compound Accumulation in Cyclocarya paliurus (Batal.) Iljinskaja" Forests 10, no. 9: 772. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090772