Forest Gap Size Alters the Functional Diversity of Soil Nematode Communities in Alpine Forest Ecosystems

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Soil Sampling
2.3. Soil Nematode Extraction
2.4. Data Calculations and Statistical Analyses
3. Results
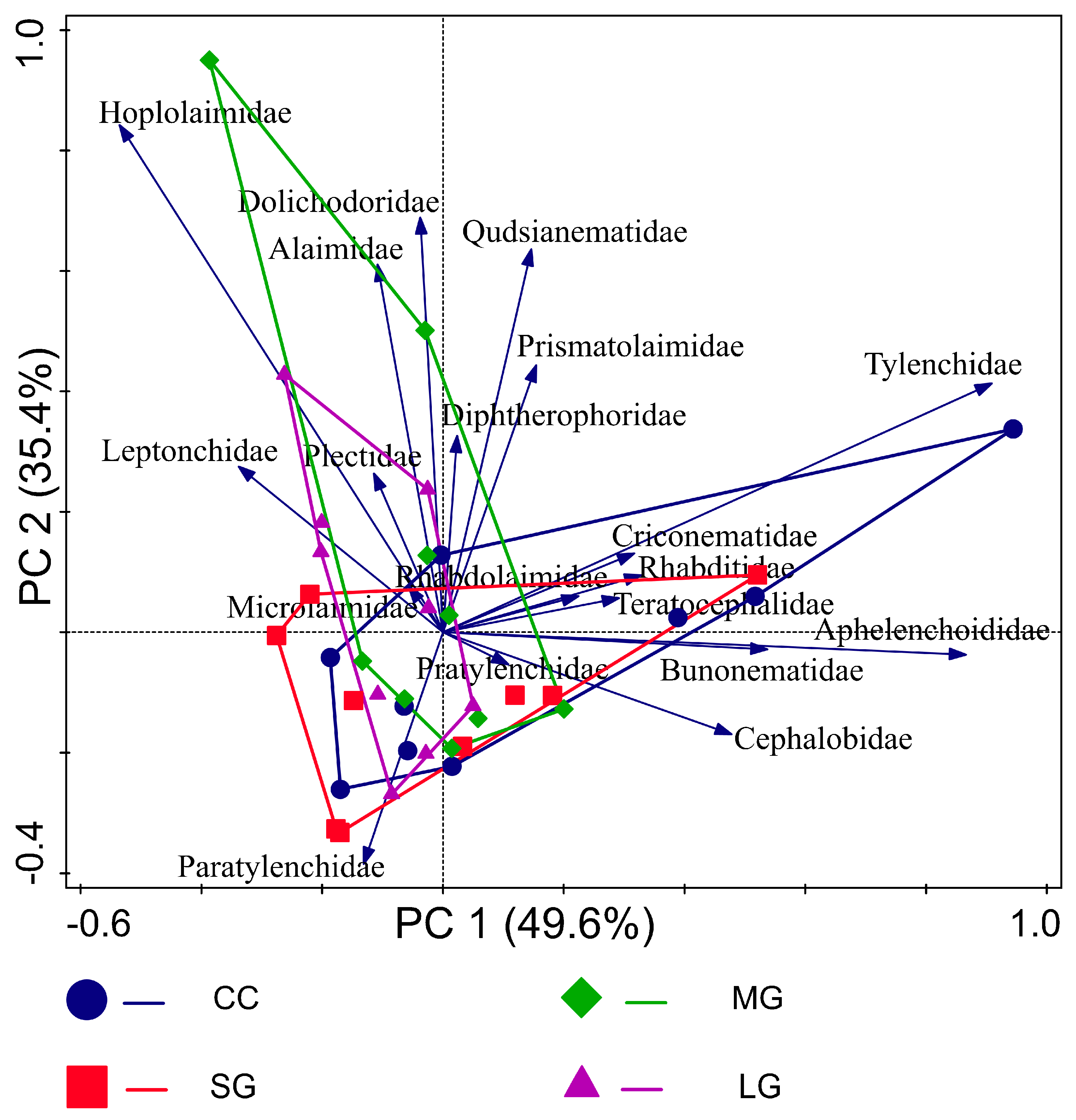
3.1. Soil Nematode Composition
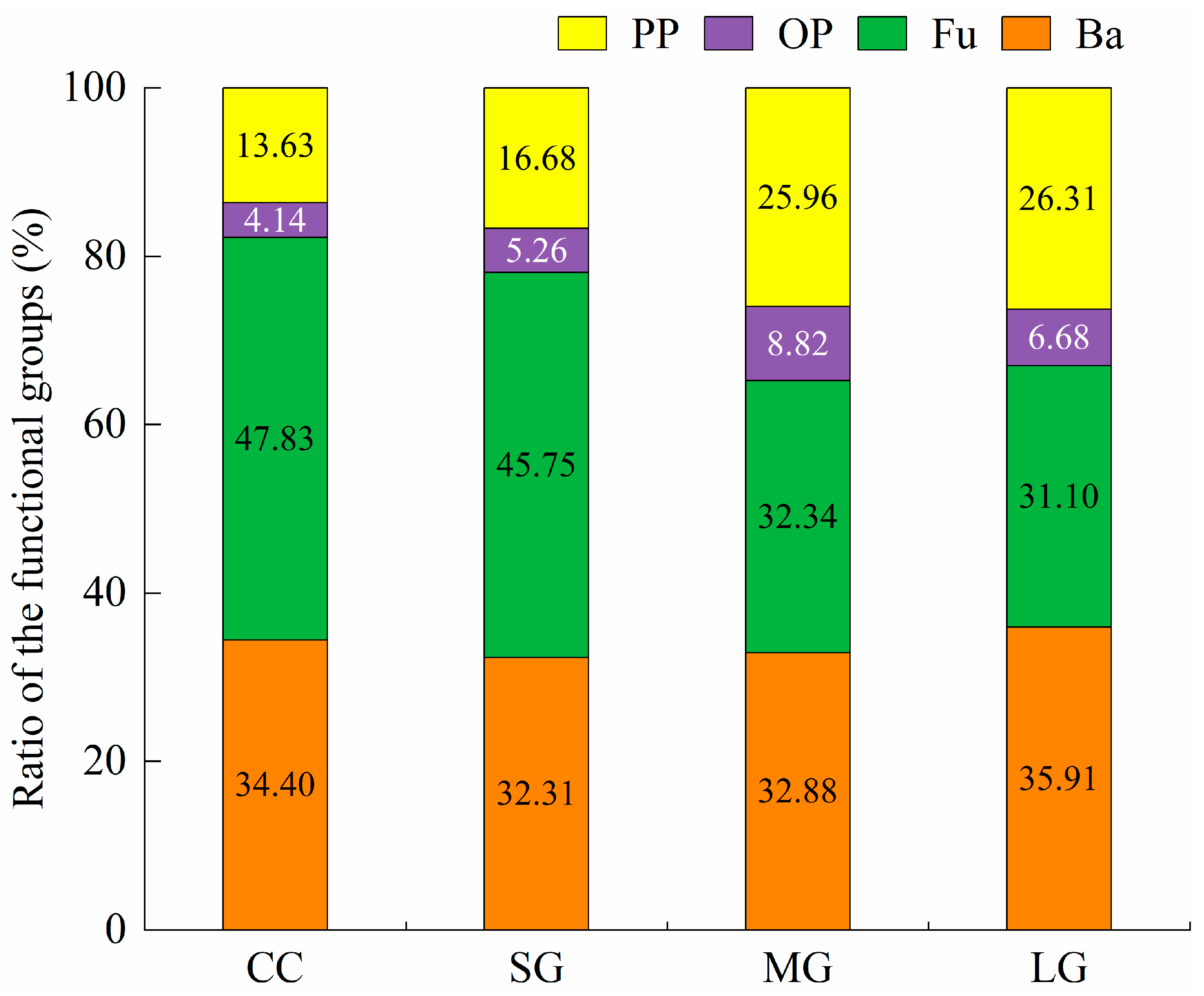
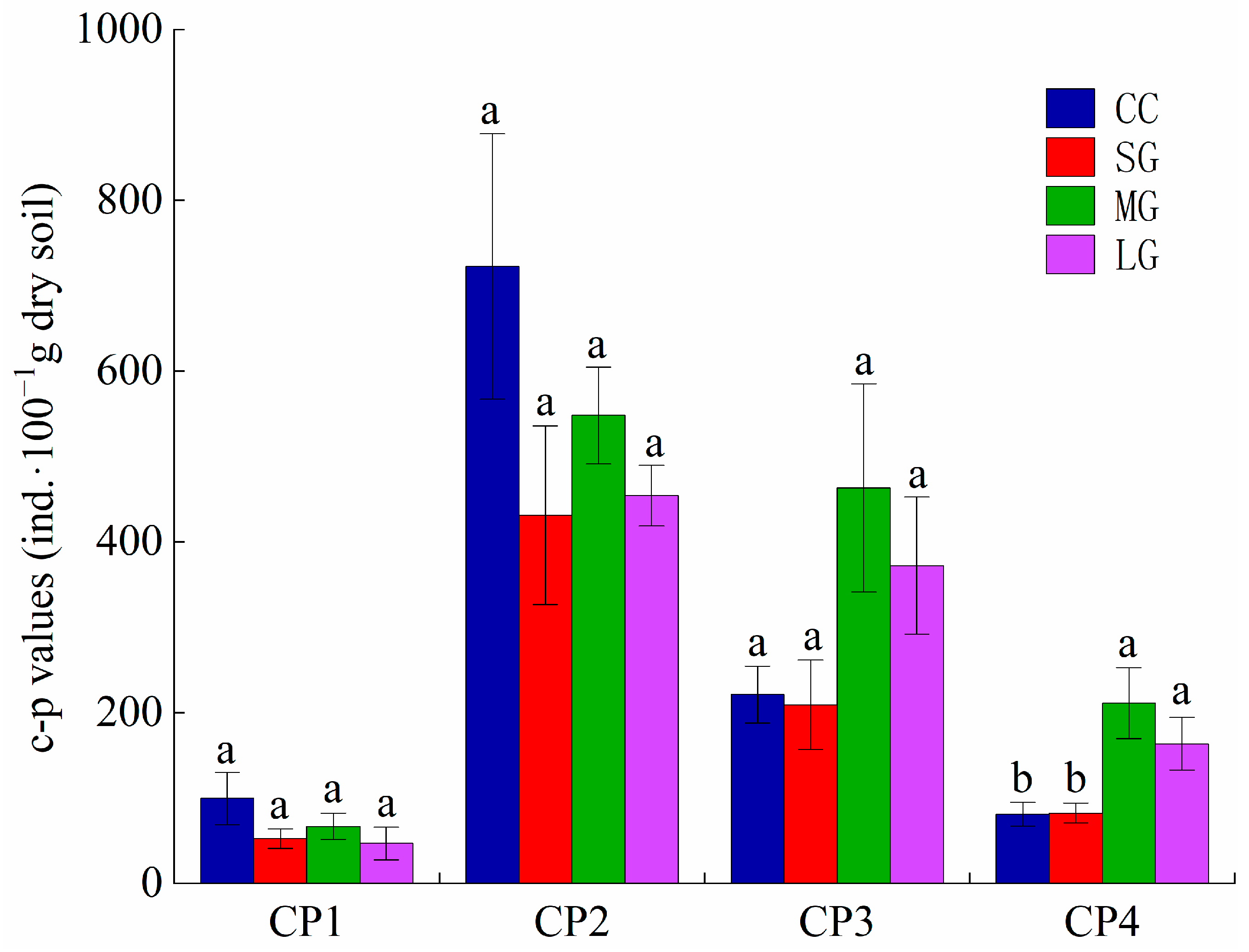
3.2. Soil Nematode Functional (Trophic, c-p) Groups
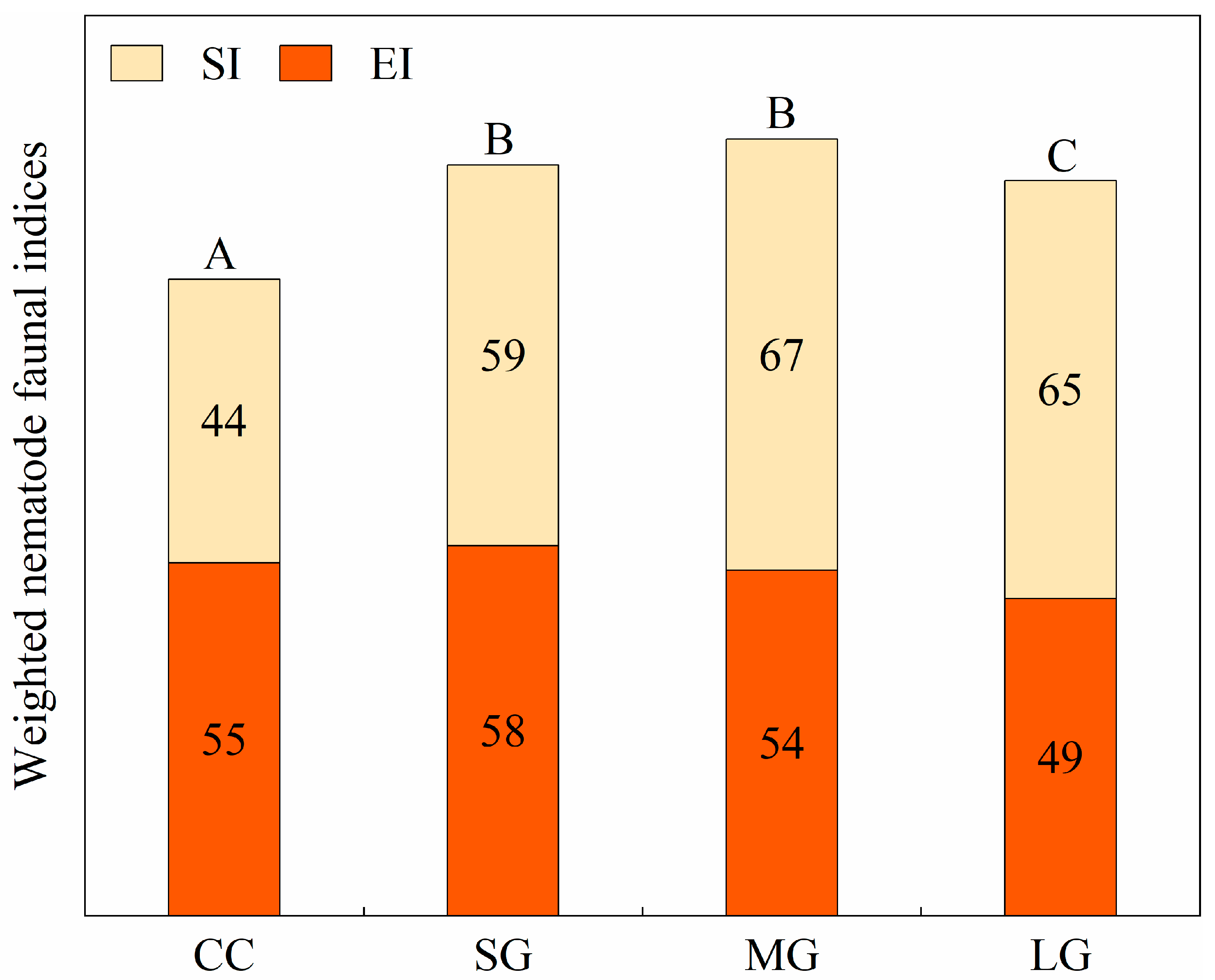
3.3. Soil Nematode Community Indices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yamamoto, S.I. Forest gap dynamics and tree regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Ritter, E.; Dalsgaard, L.; Einhorn, K.S. Light, temperature and soil moisture regimes following gap formation in a semi-natural beech-dominated forest in Denmark. For. Ecol. Manag. 2005, 206, 15–33. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Bockheim, J.G. Impacts of forest gaps on soil properties and processes in old growth northern hardwood-hemlock forests. Plant Soil 2007, 294, 219–233. [Google Scholar] [CrossRef]
- Sariyildiz, T. Effects of Gap-Size Classes on Long-Term Litter Decomposition Rates of Beech, Oak and Chestnut Species at High Elevations in Northeast Turkey. Ecosystems 2008, 11, 841–853. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Dalling, J.W.; Carson, W.P. The impact of lianas on tree regeneration in tropical forest canopy gaps: Evidence for an alternative pathway of gap-phase regeneration. J. Ecol. 2000, 88, 655–666. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Mercurio, R. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinus laricio, Poiret) stands. For. Ecol. Manag. 2007, 242, 412–418. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Butenko, K.O.; Gongalsky, K.B.; Korobushkin, D.I.; Ekschmitt, K.; Zaitsev, A.S. Forest fires alter the trophic structure of soil nematode communities. Soil Biol. Biochem. 2017, 109, 107–117. [Google Scholar] [CrossRef]
- Kitagami, Y.; Kanzaki, N.; Matsuda, Y. Distribution and community structure of soil nematodes in coastal Japanese pine forests were shaped by harsh environmental conditions. Appl. Soil Ecol. 2017, 119, 91–98. [Google Scholar] [CrossRef]
- Salamun, P.; Hanzelova, V.; Miklisova, D.; Sestinova, O.; Findorakova, L.; Kovacik, P. The effects of vegetation cover on soil nematode communities in various biotopes disturbed by industrial emissions. Sci. Total Environ. 2017, 592, 106–114. [Google Scholar] [CrossRef]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Gutierrez, C.; Fernandez, C.; Escuer, M.; Campos-Herrera, R.; Beltran Rodriguez, M.E.; Carbonell, G.; Rodriguez Martin, J.A. Effect of soil properties, heavy metals and emerging contaminants in the soil nematodes diversity. Environ. Pollut. 2016, 213, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.E.; Blumenthal, D.M.; Carrillo, Y.; Cesarz, S.; Ciobanu, M.; Hines, J.; Pabst, S.; Pendall, E.; de Tomasel, C.M.; Wall, D.H.; et al. Elevated CO2 and warming shift the functional composition of soil nematode communities in a semiarid grassland. Soil Biol. Biochem. 2016, 103, 46–51. [Google Scholar] [CrossRef]
- Olatunji, O.A.; Gong, S.; Tariq, A.; Pan, K.; Sun, X.; Chen, W.; Zhang, L.; Dakhil, M.A.; Huang, D.; Tan, X. The effect of phosphorus addition, soil moisture, and plant type on soil nematode abundance and community composition. J. Soil. Sediment. 2019, 19, 1139–1150. [Google Scholar] [CrossRef]
- Vandegehuchte, M.L.; Sylvain, Z.A.; Reichmann, L.G.; de Tomasel, C.M.; Nielsen, U.N.; Wall, D.H.; Sala, O.E. Responses of a desert nematode community to changes in water availability. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Prescott, C.E.; Blevins, L.L.; Staley, C.L. Effects of clear-cutting on decomposition rates of litter and forest floor in forests of British Columbia. Can. J. For. Res. 2000, 30, 1751–1757. [Google Scholar] [CrossRef]
- He, W.; Wu, F.; Yang, W.; Zhang, D.; Xu, Z.; Tan, B.; Zhao, Y.; Justine, M.F. Gap locations influence the release of carbon, nitrogen and phosphorus in two shrub foliar litter in an alpine fir forest. Sci. Rep. 2016, 6, 22014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, X.; Berg, B.; Yang, W.; Li, H.; Liao, S.; Tan, B.; Yue, K.; Xu, Z.; Zhang, L.; Wu, F. Formation of forest gaps accelerates C, N and P release from foliar litter during 4 years of decomposition in an alpine forest. Biogeochemistry 2018, 139, 321–335. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Wu, F.; Yang, W.; Tan, B.; Yang, Y.; Ni, X.; He, J. Characteristics of Gaps and Disturbance Regimes of the Alpine Fir Forest in Western Sichuan. Chin. J. Appl. Environ. Biol. 2013, 19, 922–928. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, F.; Tan, B.; Yang, W.; He, W.; Ni, X. Effects of gap sizes on foliar litter decomposition in alpine forests. Acta Ecol. Sin. 2016, 36, 3537–3545. [Google Scholar] [CrossRef]
- Tan, B.; Wu, F.; Yang, W.; Yu, S.; Liu, L.; Wang, A.; Yang, Y. Seasonal dynamics of soil fauna in the subalpine and alpine forests of west Sichuan at different altitudes. Acta Ecol. Sin. 2013, 33, 12–22. [Google Scholar] [CrossRef]
- Schliemann, S.A.; Bockheim, J.G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. The definition of treefall gap and its effect on measures of forest dynamics. Biotropica 1982, 14, 158–160. [Google Scholar] [CrossRef]
- Runkle, J.R. Gap regeneration in some old-growth forests of the Eastern United States. Ecology 1981, 62, 1041–1051. [Google Scholar] [CrossRef]
- Liu, Q.H.; Hytteborn, H. Gap structure, disturbance and regeneration in a primeval Picea abies forest. J. Veg. Sci. 1991, 2, 391–402. [Google Scholar] [CrossRef]
- Liang, W.; Li, Q.; Jiang, Y.; Neher, D.A. Nematode faunal analysis in an aquic brown soil fertilised with slow-release urea, Northeast China. Appl. Soil Ecol. 2005, 29, 185–192. [Google Scholar] [CrossRef]
- Yin, W.Y. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 1998. [Google Scholar]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Zhang, X.K.; Liang, W.J.; Li, Q. Forest Soil Nematodes in Changbai Mountain: Morphology and Distribution; China Agriculture Press: Beijing, China, 2013. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collection. J. Theoret. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T. Nematode diversity in agroecosystems. Agr. Ecosyst. Environ. 1999, 74, 113–135. [Google Scholar] [CrossRef]
- Yeates, G.W. Variation in soil nematode diversity under pasture with soil and year. Soil Biol. Biochem. 1984, 16, 95–102. [Google Scholar] [CrossRef]
- Bongers, T.; van der Meulen, H.; Korthals, G. Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Appl. Soil Ecol. 1997, 6, 195–199. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J.; Yang, W.; Wu, F.; Xu, Z.; Tan, B.; Zhang, L.; He, X.; Guo, L. Canopy gaps accelerate soil organic carbon retention by soil microbial biomass in the organic horizon in a subalpine fir forest. Appl. Soil Ecol. 2018, 125, 169–176. [Google Scholar] [CrossRef]
- Prescott, C.E.; Hope, G.D.; Blevins, L.L. Effect of gap size on litter decomposition and soil nitrate concentrations in a high-elevation spruce–fir forest. Can. J. For. Res. 2003, 33, 2210–2220. [Google Scholar] [CrossRef]
- Xue, H.; Luo, D. Characteristics of soil nematode community in clear cutting slash of Fir forests in southeast Tibet. Sci. Silvae Sin. 2013, 49, 107–114. [Google Scholar] [CrossRef]
- Xue, H.Y.; Luo, D.Q. Characteristics of soil nematode community in Abies georgei var. smithii forest gaps in southeast Tibet, China. Chin. J. Appl. Ecol. 2013, 24, 2494–2502. [Google Scholar]
- Zhang, R.; Liu, X.; Zhong, H.; Wu, Q.; Wu, P. Distribution pattern of soil nematode communities along the vertical climate zones on the eastern slope of Gongga Mountain. Chin. J. Appl. Environ. Biol. 2016, 22, 959–971. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, W.; Wu, F.; Yang, F.; Lan, L.; Liu, Y.; Guo, C.; Tan, B. Diversity of soil nematode communities in the subalpine and alpine forests of western Sichuan, China. Chin. J. Appl. Ecol. 2017, 28, 3360–3368. [Google Scholar] [CrossRef]
- Walker, B.H. Biodiversity and Ecological Redundancy. Conserv. Biol. 1992, 6, 18–23. [Google Scholar] [CrossRef]
- Lin, N.; Bartsch, N.; Heinrichs, S.; Vor, T. Long-term effects of canopy opening and liming on leaf litter production, and on leaf litter and fine-root decomposition in a European beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2015, 338, 183–190. [Google Scholar] [CrossRef]
- He, W.; Wu, F.; Yang, W.; Tan, B.; Zhao, Y.; Wu, Q.; He, M. Lignin Degradation in Foliar Litter of Two Shrub Species from the Gap Center to the Closed Canopy in an Alpine Fir Forest. Ecosystems 2015, 19, 115–128. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y. Effects of Forest Gaps on Litter Lignin and Cellulose Dynamics Vary Seasonally in an Alpine Forest. Forests 2016, 7, 27. [Google Scholar] [CrossRef]
- Feng, R.; Yang, W.; Zhang, J. Review on biochemical property in forest soil organic layer and its responses to climate change. Chin. J. Appl. Environ. Biol. 2006, 12, 734–739. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef]
- Jones, D.; Farmer, V.C. The ecology and physiology of soil fungi involved in the degradation of lignin and related aromatic compounds. Eur. J. Soil Sci. 1967, 18, 74–84. [Google Scholar] [CrossRef]
- Lin, D.; Pang, M.; Fanin, N.; Wang, H.; Qian, S.; Zhao, L.; Yang, Y.; Mi, X.; Ma, K. Fungi participate in driving home-field advantage of litter decomposition in a subtropical forest. Plant Soil 2019, 434, 467–480. [Google Scholar] [CrossRef]
- Bakonyi, G.; Nagy, P. Temperature- and moisture-induced changes in the structure of the nematode fauna of a semiarid grassland-patterns and mechanisms. Glob. Chang. Biol. 2000, 6, 697–707. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Gap | Gap Size (m2) | Cause of Gap | Gap Age (Year) | Gap Makers | Soil Temperature (°C) | Soil Moisture (%) |
|---|---|---|---|---|---|---|
| Large gap | 281.7 ± 4.4 | Breakage at trunk | 31.4 ± 2.7 | Fir (78%) + spruce (22%) | 9.23 ± 0.22 a | 40.72 ± 2.33 ab |
| Medium gap | 165.0 ± 5.8 | Breakage at trunk | 30.1 ± 2.8 | Fir (71%) + spruce (29%) | 7.81 ± 0.15 b | 37.54 ± 2.69 ab |
| Small gap | 43.3 ± 3.3 | Standing death | 26.7 ± 1.9 | Fir | 7.48 ± 0.14 c | 36.67 ± 1.94 b |
| Closed canopy | - | - | - | - | 7.42 ± 0.16 c | 43.71 ± 1.72 a |
| TG/Genera | FG | Closed Canopy | Small Gaps | Medium Gaps | Large Gaps | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ind. | Dominance | Ind. | Dominance | Ind. | Dominance | Ind. | Dominance | Ind. | Dominance | ||
| Ba | 386.5 | 250.2 | 423.7 | 370.1 | 1430.4 | ||||||
| Rhabditis | Ba1 | 92.1 | ++ | 45.4 | ++ | 61.4 | ++ | 40.6 | ++ | 239.5 | ++ |
| Bunonema | Ba1 | 7.1 | + | 6.7 | + | 5.0 | + | 0.9 | + | 19.7 | + |
| Eucephalobus | Ba2 | 35.3 | ++ | 21.2 | ++ | 26.6 | ++ | 17.8 | ++ | 100.9 | ++ |
| Heterocephalobus | Ba2 | 6.5 | + | 0.9 | + | 1.3 | + | 2.0 | + | 10.6 | + |
| Acrobeloides | Ba2 | 62.4 | ++ | 17.6 | ++ | 35.1 | ++ | 29.9 | ++ | 145.0 | ++ |
| Plectus | Ba2 | 36.4 | ++ | 27.8 | ++ | 75.1 | ++ | 82.4 | ++ | 221.6 | ++ |
| Wilsonema | Ba2 | 31.9 | ++ | 22.9 | ++ | 26.4 | ++ | 23.5 | ++ | 104.7 | ++ |
| Microlaimus | Ba2 | 1.7 | + | 0.9 | + | 10.7 | + | 5.2 | + | 18.5 | + |
| Teratocephalus | Ba3 | 43.9 | ++ | 55.8 | ++ | 70.4 | ++ | 66.3 | ++ | 236.4 | ++ |
| Metateratocephalus | Ba3 | 10.2 | + | 5.2 | + | 0 | - | 0.7 | + | 16.1 | + |
| Rhabdolaimus | Ba3 | 3.8 | + | 0 | - | 4.2 | + | 1.9 | + | 9.9 | + |
| Prismatolaimus | Ba3 | 30.5 | ++ | 17.2 | ++ | 53.7 | ++ | 25.9 | ++ | 127.4 | ++ |
| Alaimus | Ba4 | 11.3 | ++ | 16.1 | ++ | 38.6 | ++ | 63.2 | ++ | 129.2 | ++ |
| Paramphidelus | Ba4 | 13.4 | ++ | 12.4 | ++ | 15.2 | ++ | 9.8 | + | 50.9 | ++ |
| Fu | 537.3 | 354.2 | 416.7 | 320.5 | 1628.8 | ||||||
| Filenchus | Fu2 | 421.3 | +++ | 278.7 | +++ | 312.8 | +++ | 261.6 | +++ | 1274.4 | +++ |
| Aphelenchoides | Fu2 | 89.3 | ++ | 48.1 | ++ | 23.8 | ++ | 10.4 | ++ | 171.6 | ++ |
| Diphtherophora | Fu3 | 17.1 | ++ | 14.6 | ++ | 36.5 | ++ | 27.0 | ++ | 95.2 | ++ |
| Tylencholaimus | Fu4 | 9.5 | + | 12.9 | ++ | 43.6 | ++ | 21.5 | ++ | 87.5 | ++ |
| OP | 46.5 | 40.7 | 113.7 | 68.8 | 269.7 | ||||||
| Epidorylaimus | OP4 | 5.9 | + | 9.9 | ++ | 47.0 | ++ | 20.3 | ++ | 83.0 | ++ |
| Dorydorella | OP4 | 40.6 | ++ | 30.8 | ++ | 66.7 | ++ | 48.6 | ++ | 186.6 | ++ |
| PP | 153.1 | 129.1 | 334.5 | 271.2 | 887.8 | ||||||
| Coslenchus | PP2 | 2.1 | + | 0 | - | 0 | - | 0 | - | 2.1 | + |
| Basiria | PP2 | 24.7 | ++ | 4.1 | + | 26.2 | ++ | 6.7 | + | 61.6 | ++ |
| Paratylenchus | PP2 | 10.9 | + | 8.7 | ++ | 10.1 | + | 14.4 | ++ | 44.1 | ++ |
| Nagelus | PP3 | 15.4 | ++ | 6.9 | + | 33.8 | ++ | 4.5 | + | 60.6 | ++ |
| Pararotylenchus | PP3 | 90.1 | ++ | 102.0 | +++ | 259.8 | +++ | 237.5 | +++ | 689.3 | +++ |
| Pratylenchus | PP3 | 3.4 | + | 3.4 | + | 1.1 | + | 0.7 | + | 8.6 | + |
| Macroposthonia | PP3 | 6.5 | + | 3.9 | + | 3.6 | + | 7.5 | + | 21.5 | + |
| Total individuals | 1123.3 | 774.3 | 1288.5 | 1030.6 | 4216.7 | ||||||
| Total genera | 27 | 25 | 25 | 26 | 27 | ||||||
| Indices | Closed Canopy | Small Gaps | Medium Gaps | Large Gaps |
|---|---|---|---|---|
| H’ | 2.09 ± 0.24a | 2.09 ± 0.26a | 2.31 ± 0.24a | 2.17 ± 0.19a |
| J’ | 0.78 ± 0.08a | 0.76 ± 0.07a | 0.82 ± 0.06a | 0.80 ± 0.05a |
| λ | 0.19 ± 0.07a | 0.21 ± 0.08a | 0.16 ± 0.05a | 0.17 ± 0.04a |
| SR | 2.01 ± 0.32a | 2.22 ± 0.38a | 2.23 ± 0.41a | 2.08 ± 0.47a |
| MI | 2.19 ± 0.13c | 2.38 ± 0.18b | 2.53 ± 0.14a | 2.52 ± 0.14ab |
| ∑MI | 2.29 ± 0.19b | 2.47 ± 0.20a | 2.59 ± 0.17a | 2.60 ± 0.16a |
| MI2-5 | 2.32 ± 0.10b | 2.52 ± 0.21a | 2.65 ± 0.17a | 2.61 ± 0.12a |
| PPI | 2.68 ± 0.29a | 2.88 ± 0.12a | 2.73 ± 0.33a | 2.80 ± 0.28a |
| PPI/MI | 1.22 ± 0.10a | 1.22 ± 0.10a | 1.08 ± 0.11b | 1.11 ± 0.10b |
| EI | 54.92 ± 8.50a | 57.53 ± 8.13a | 53.74 ± 7.60a | 49.31 ± 10.45a |
| SI | 44.07 ± 8.84b | 59.21 ± 13.48a | 67.01 ± 8.47a | 64.99 ± 6.88a |
| CI | 59.42 ± 24.57a | 57.59 ± 17.12a | 58.90 ± 18.25a | 69.02 ± 24.18a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Yang, W.; Zhang, J.; Xu, Z.; Zhang, L.; Liu, Y.; Li, H.; You, C.; Tan, B. Forest Gap Size Alters the Functional Diversity of Soil Nematode Communities in Alpine Forest Ecosystems. Forests 2019, 10, 806. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090806
Shen Y, Yang W, Zhang J, Xu Z, Zhang L, Liu Y, Li H, You C, Tan B. Forest Gap Size Alters the Functional Diversity of Soil Nematode Communities in Alpine Forest Ecosystems. Forests. 2019; 10(9):806. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090806
Chicago/Turabian StyleShen, Ya, Wanqin Yang, Jian Zhang, Zhenfeng Xu, Li Zhang, Yang Liu, Han Li, Chengming You, and Bo Tan. 2019. "Forest Gap Size Alters the Functional Diversity of Soil Nematode Communities in Alpine Forest Ecosystems" Forests 10, no. 9: 806. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090806