Southeastern Rear Edge Populations of Quercus suber L. Showed Two Alternative Strategies to Cope with Water Stress

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Pressure–Volume Relationships
2.3. Proline Concentration
2.4. Statistical Analysis
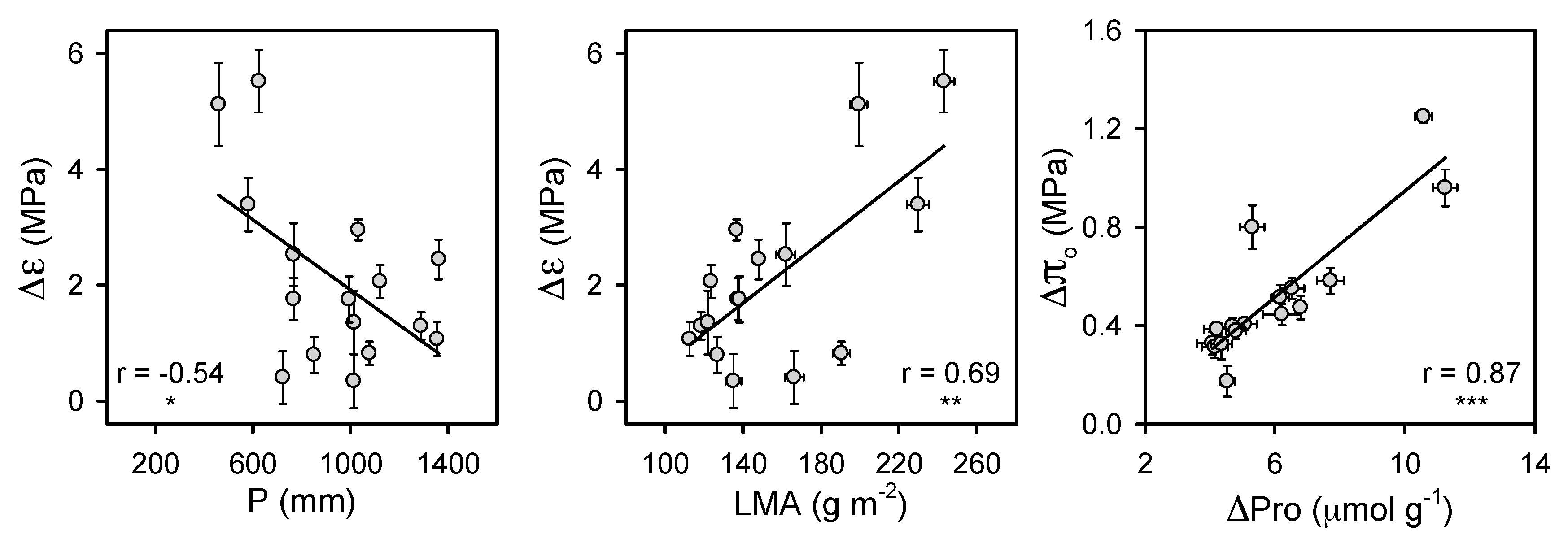
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walter, H. Vegetation of the Earth and Ecological Systems of the Geo-Biosphere, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1985. [Google Scholar]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M.; et al. The Mediterranean climate: An overview of the main characteristics and issues. In Mediterranean Climate Variability; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 1–26. ISBN 978-0-444-52170-5. [Google Scholar]
- Gil-Pelegrín, E.; Saz, M.A.; Cuadrat, J.M.; Peguero-Pina, J.J.; Sancho-Knapik, D. Oaks under Mediterranean-type climates: Functional response to summer aridity. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus quercus L.; Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 137–193. ISBN 978-3-319-69099-5. [Google Scholar]
- Mediavilla, S.; Escudero, A. Stomatal responses to drought at a Mediterranean site: A comparative study of co-occurring woody species differing in leaf longevity. Tree Physiol. 2003, 23, 987–996. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Responses and adaptations. Environ. Exp. Bot. 2014, 103, 68–79. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Keenan, T. Photosynthetic responses to stress in Mediterranean evergreens: Mechanisms and models. Environ. Exp. Bot. 2014, 103, 24–41. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Vilagrosa, A.; Alonso-Forn, D.; Ferrio, J.P.; Sancho-Knapik, D.; Gil-Pelegrín, E. Living in Drylands: Functional Adaptations of Trees and Shrubs to Cope with High Temperatures and Water Scarcity. Forests 2020, 11, 1028. [Google Scholar] [CrossRef]
- Mooney, H.A.; Dunn, E.L. Convergent Evolution of Mediterranean-Climate Evergreen Sclerophyll Shrubs. Evolution 1970, 24, 292–303. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.G.; Fonseca, C.R.; Overton, J.M.; Westoby, M. Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? Funct. Ecol. 2003, 17, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Ackerly, D.D. Adaptation, niche conservatism, and convergence: Comparative studies of leaf evolution in the California chaparral. Am. Nat. 2004, 163, 654–671. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Xu, L. What limits evaporation from Mediterranean oak woodlands—The supply of moisture in the soil, physiological control by plants or the demand by the atmosphere? Adv. Water Resour. 2007, 30, 2113–2122. [Google Scholar] [CrossRef]
- Alonso-Forn, D.; Sancho-Knapik, D.; Ferrio, J.P.; Peguero-Pina, J.J.; Bueno, A.; Onoda, Y.; Cavender-Bares, J.; Niinemets, Ü.; Jansen, S.; Riederer, M.; et al. Revisiting the Functional Basis of Sclerophylly within the Leaf Economics Spectrum of Oaks: Different Roads to Rome. Curr. For. Rep. 2020. [Google Scholar] [CrossRef]
- Bethoux, J.P.; Gentili, B.; Morin, P.; Nicolas, E.; Pierre, C.; Ruiz-Pino, D. The Mediterranean Sea: A miniature ocean for climatic and environmental studies and a key for climatic functioning of the North Atlantic. Prog. Oceanogr. 1999, 44, 131–146. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Villar-Salvador, P.; Pérez-Rontomé, C.; Maestro-Martínez, M.; Montserrat-Martí, G. Leaf morphology and leaf chemical composition in three Quercus (Fagaceae) species along a rainfall gradient in NE Spain. Trees 1997, 11, 127–134. [Google Scholar] [CrossRef]
- Bussotti, F.; Bettini, D.; Grossoni, P.; Mansuino, S.; Nibbi, R.; Soda, C.; Tani, C. Structural and functional traits of Quercus ilex in response to water availability. Environ. Exp. Bot. 2002, 47, 11–23. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Leaf mass per area ratio in Quercus ilex leaves under a wide range of climatic conditions. The importance of low temperatures. Acta Oecol. 2007, 31, 168–173. [Google Scholar] [CrossRef]
- Niinemets, Ü. Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex. New Phytol. 2015, 205, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Barrón, E.; Camarero, J.J.; Vilagrosa, A.; Gil-Pelegrín, E. Morphological and physiological divergences within Quercus ilex support the existence of different ecotypes depending on climatic dryness. Ann. Bot. 2014, 114, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Gil-Pelegrín, E.; Notivol, E. Aridity promotes differences in proline and phytohormone levels in Pinus pinaster populations from contrasting environments. Trees 2012, 26, 799–808. [Google Scholar] [CrossRef]
- Sancho-Knapik, D.; Sanz, M.A.; Peguero-Pina, J.J.; Niinemets, Ü.; Gil-Pelegrín, E. Changes of secondary metabolites in Pinus sylvestris L. needles under increasing soil water deficit. Ann. For. Sci. 2017, 74, 24. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Xiao, X.; Xu, X.; Yang, F. Adaptive responses to progressive drought stress in two Populus cathayana populations. Silva Fenn. 2008, 42, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Vilagrosa, A.; Morales, F.; Abadía, A.; Bellot, J.; Cochard, H.; Gil-Pelegrín, E. Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species. Environ. Exp. Bot. 2010, 69, 233–242. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Ardy, R.; Zhang, Y.; Sun, S.; Cao, K.; Sack, L. Rapid determination of comparative drought tolerance traits: Using an osmometer to predict turgor loss point. Methods Ecol. Evol. 2012, 3, 880–888. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Functional groups in Quercus species derived from the analysis of pressure–volume curves. Trees 2002, 16, 465–472. [Google Scholar] [CrossRef]
- Singh, S.P.; Zobel, D.B.; Garkoti, S.C.; Tewari, A.; Negi, C.M.S. Patterns in water relations of central Himalayan trees. Trop. Ecol. 2006, 47, 159–182. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; García-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Iglesias, A.; Garrote, L.; Flores, F.; Moneo, M. Challenges to manage the risk of water scarcity and climate change in the Mediterranean. Water Resour. Manag. 2007, 21, 775–788. [Google Scholar] [CrossRef]
- García-Mozo, H.; Mestre, A.; Galán, C. Phenological trends in southern Spain: A response to climate change. Agric. For. Meteorol. 2010, 150, 575–580. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Sancho-Knapik, D.; Martin-StPaul, N.K.; Limousin, J.M.; McDowell, N.G.; Gil-Pelegrín, E. Drought-Induced Oak Decline—Factors Involved, Physiological Dysfunctions, and Potential Attenuation by Forestry Practices. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus quercus L.; Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 419–451. ISBN 978-3-319-69099-5. [Google Scholar]
- Sancho-Knapik, D.; Peguero-Pina, J.J.; Cremer, E.; Camarero, J.J.; Fernández-Cancio, Á.; Ibarra, N.; Konnert, M.; Gil-Pelegrín, E. Genetic and environmental characterization of Abies alba Mill. populations at its western rear edge. Pirineos 2014, 169, e007. [Google Scholar] [CrossRef] [Green Version]
- Gil-Pelegrín, E.; Peguero-Pina, J.J.; Sancho-Knapik, D. Oaks and People: A Long Journey Together. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus quercus L.; Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 1–11. ISBN 978-3-319-69099-5. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.N.; Bemmels, J.B. Time to get moving: Assisted gene flow of forest trees. Evol. Appl. 2016, 9, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Soto, A.; Lorenzo, Z.R.; Gil, L. Differences in fine-scale genetic structure and dispersal in Quercus ilex L. and Q. suber L.: Consequences for regeneration of Mediterranean open woods. Heredity 2007, 99, 601–607. [Google Scholar] [CrossRef]
- Mosmann, V.; Castro, A.; Fraile, R.; Dessens, J.; Sánchez, J.L. Detection of statistically significant trends in the summer precipitation of mainland Spain. Atmos. Res. 2004, 70, 43–53. [Google Scholar] [CrossRef]
- Vilagrosa, A.; Bellot, J.; Vallejo, V.R.; Gil-Pelegrín, E. Cavitation, stomatal conductance, and leaf dieback in seedlings of two co-occurring Mediterranean shrubs during an intense drought. J. Exp. Bot. 2003, 54, 2015–2024. [Google Scholar] [CrossRef]
- Bates, S.L.; Waldre, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The world-wide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Wright, I.J.; Evans, J.R. Leaf mesophyll diffusion conductance in 35 Australian sclerophylls covering a broad range of foliage structural and physiological variation. J. Exp. Bot. 2009, 60, 2433–2449. [Google Scholar] [CrossRef] [Green Version]
- Peguero-Pina, J.J.; Sisó, S.; Sancho-Knapik, D.; Díaz-Espejo, A.; Flexas, J.; Galmés, J.; Gil-Pelegrín, E. Leaf morphological and physiological adaptations of a deciduous oak (Quercus faginea Lam.) to the Mediterranean climate: A comparison with a closely related temperate species (Quercus robur L.). Tree Physiol. 2016, 36, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Nabais, C.; Freitas, H.; Hagemeyer, J. Tree-rings to climate relationships of Quercus ilex L. in the NE-Portugal. Dendrochronologia 1998, 16–17, 37–44. [Google Scholar]
- Urli, M.; Porté, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- Pinto, C.A.; David, J.S.; Cochard, H.; Caldeira, M.C.; Henriques, M.O.; Quilhó, T.; Paço, T.A.; Pereira, J.S.; David, T.S. Drought-induced embolism in current-year shoots of two Mediterranean evergreen oaks. For. Ecol. Manag. 2012, 285, 1–10. [Google Scholar] [CrossRef]
- Tyree, M.T.; Cochard, H. Summer and winter embolism in oak: Impact on water relations. Ann. For. Sci. 1996, 53, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Morales, F.; Flexas, J.; Gil-Pelegrín, E. Differential photosynthetic performance and photoprotection mechanisms of three Mediterranean evergreen oaks under severe drought stress. Funct. Plant Biol. 2009, 36, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Gullo, M.A.; Salleo, S. Different strategies of drought resistance in three Mediterranean sclerophyllous trees growing in the same environmental conditions. New Phytol. 1988, 108, 267–276. [Google Scholar] [CrossRef]
- Nadal, M.; Roig-Oliver, M.; Bota, J.; Flexas, J. Leaf age-dependent elastic adjustment and photosynthetic performance under drought stress in Arbutus unedo seedlings. Flora 2020, 272, 151662. [Google Scholar] [CrossRef]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Binks, O.; Meir, P.; Rowland, L.; da Costa, A.C.L.; Vasconcelos, S.S.; de Oliveira, A.A.R.; Ferreira, L.; Christoffersen, B.; Nardini, A.; Mencuccini, M. Plasticity in leaf-level water relations of tropical rainforest trees in response to experimental drought. New Phytol. 2016, 211, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.M.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Diamantoglou, S.; Rhizopoulou, S. Free proline accumulation in sapwood, bark and leaves of three evergreen sclerophylls and a comparison with an evergreen conifer. J. Plant Physiol. 1992, 140, 361–365. [Google Scholar] [CrossRef]
- Xiao, X.; Yang, F.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress. Physiol. Plant. 2009, 136, 150–168. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Code | Latitude | Longitude | Altitude (m a.s.l.) | P (mm) | T (°C) | MAI |
|---|---|---|---|---|---|---|---|
| Dar Fatma | DF | 36.82° N | 8.77° E | 710 | 1361 | 14.4 | 55.9 |
| Echihia (Ain Zana) | E | 36.79° N | 8.74° E | 850 | 1355 | 14.7 | 55.0 |
| El Feija | EF | 36.77° N | 8.65° E | 800 | 1289 | 15.2 | 51.2 |
| Babouch | Bab | 36.80° N | 8.68° E | 277 | 1121 | 15.9 | 43.3 |
| Megen Sef | MS | 36.77° N | 8.78° E | 548 | 1077 | 15.5 | 42.3 |
| Beni Mtir | BM | 36.73° N | 8.73° E | 650 | 1014 | 15.7 | 39.4 |
| Jebel Khroufa (Ouchtata) | O | 36.93° N | 8.943° E | 200 | 1031 | 16.2 | 39.4 |
| Jebel Jouza | JJ | 36.83° N | 8.99° E | 540 | 1013 | 15.8 | 39.2 |
| Aïn Snoussi | AS | 36.82° N | 8.92° E | 522 | 993 | 15.8 | 38.5 |
| Hammam Bourguiba | HB | 36.76° N | 8.59° E | 265 | 850 | 17.4 | 31.0 |
| Bellif | Bl | 37.04° N | 9.07° E | 158 | 767 | 17.6 | 27.8 |
| Ras rajel | Rr | 36.951° N | 8.872° E | 62 | 766 | 17.8 | 27.5 |
| Fernana | Frn | 36.65° N | 8.70° E | 699 | 722 | 17.2 | 26.5 |
| Kef er rennd | Ker | 36.823° N | 10.769° E | 637 | 624 | 16.4 | 23.6 |
| Jebel Chehid, Ramlia | JCH | 36.37° N | 9.31° E | 612 | 580 | 17.2 | 21.4 |
| Hammem Jdidi | HJ | 36.39° N | 10.38° E | 132 | 459 | 18.9 | 15.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghouil, H.; Sancho-Knapik, D.; Ben Mna, A.; Amimi, N.; Ammari, Y.; Escribano, R.; Alonso-Forn, D.; Ferrio, J.P.; Peguero-Pina, J.J.; Gil-Pelegrín, E. Southeastern Rear Edge Populations of Quercus suber L. Showed Two Alternative Strategies to Cope with Water Stress. Forests 2020, 11, 1344. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121344
Ghouil H, Sancho-Knapik D, Ben Mna A, Amimi N, Ammari Y, Escribano R, Alonso-Forn D, Ferrio JP, Peguero-Pina JJ, Gil-Pelegrín E. Southeastern Rear Edge Populations of Quercus suber L. Showed Two Alternative Strategies to Cope with Water Stress. Forests. 2020; 11(12):1344. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121344
Chicago/Turabian StyleGhouil, Hana, Domingo Sancho-Knapik, Amira Ben Mna, Nabil Amimi, Youssef Ammari, Rubén Escribano, David Alonso-Forn, Juan Pedro Ferrio, José Javier Peguero-Pina, and Eustaquio Gil-Pelegrín. 2020. "Southeastern Rear Edge Populations of Quercus suber L. Showed Two Alternative Strategies to Cope with Water Stress" Forests 11, no. 12: 1344. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121344