1. Introduction
Knowledge on the patterns of biomass partitioning of a variety of species is of high importance for carbon reporting [
1], tree physiology, plant ecology, and process-based modelling [
2], and has also applications for forestry management [
3]. Although generalised biomass models can be used to estimate biomass stock in forests stands [
4], locally fitted models are recommended by the IPCC to minimise the bias of estimations [
5]. In addition, young stands are characterised by fast changes in biomass allocation in individual tree compartments, due to which biomass equations developed for older stands are not appropriate [
6,
7,
8].
Considering a range of forest tree species, a lot of works focused on biomass allocation and foliage traits with regard to a number of different factors, e.g., competition in a stand [
9,
10]. Few authors [
11,
12,
13] used the ratio between foliage dry mass and total dry plant biomass (leaf mass ratio; LMR) or between foliage area and total plant dry biomass (leaf area ratio; LAR) to link them to ecological and production processes. Similarly, specific leaf area (ratio between leaf area and dry leaf mass; SLA) has often been studied in plant ecology as one of the adaptation strategy indicators [
14]. Canopy leaf area of forests serves as a dominant physical control over primary production, transpiration, energy exchange, and other physiological attributes related to a variety of ecosystem processes; hence, it is a substantial element of ecological studies [
15]. Here, the leaf area index (LAI, defined as the amount of leaf area in the canopy per unit ground area) is the main parameter of the canopy [
16]. None of these indicators has been studied in common aspen either at the tree or the stand level.
Common aspen, or also named Eurasian aspen (
Populus tremula L.), is one of the most widely distributed tree species around the world [
17]. It is a light-demanding broadleaved softwood tree species that is native to boreal zones as well as to cooler parts, such as the temperate zones of Europe and Asia [
18]. It is one of the so-called pioneer species that occupy post-disturbance areas after windbreaks, fire events, or clear cutting [
17,
19]. Although in forest stands dominated by other species it is usually scattered, it can create homogeneous stands at post-disturbance areas [
20].
The aspen grows very fast, especially during the first 15–20 years, i.e., in the period when crown competition increases [
20]. Even though the commercial importance of aspen is limited, the species is often considered as a relevant part of forest ecosystems due to its fundamental importance for other plant and animal species [
21,
22]. This species creates conditions for existence of a variety of herbivorous, saprophytic invertebrates, fungi and lichens, birds, etc. [
20]. It is one of the most attractive tree species for forage for large herbivores [
23]. Hence, its presence increases the carrying capacity of red deer hunting grounds and can ensure the biological protection of other economically important tree species [
24].
Moreover, the species is considered to be soil ameliorating since its foliage litter contains high concentrations of calcium, potassium, and magnesium [
19]. It is also an important species for regulating the microclimate and for enhancing the structural and biological diversity of open agricultural landscapes in the temperate zone of Europe [
22,
25].
The share of aspen in Slovak forests is small, since from the point of its contribution to the wood stock and forest area it was ranked 18th and 16th, respectively, out of all tree species (approximately 50) recorded in the national forestry inventory [
26]. According to the data from the last National Forest Inventory of Slovakia, aspen was recorded at 8% of the inventory plots [
26]. Although it occurred at elevations from 200 to 1300 m a.s.l., aspen was most frequent at elevations between 450 and 550 m a.s.l. [
10]. Despite its good ecological values, it is not an important tree species for any biotopes of European and national significance, nor is it a main tree species of any biotope. In Slovakia and Czechia, aspen (similarly to many other softwood broadleaved tree species) occur more frequently in stands growing on former agricultural land than on forest lands [
27].
Since common aspen is fast growing and tolerant to extreme or nutrient-poor ecological conditions, it might be a prospective species to produce biomass for energetic purposes or for pulp and paper production [
28]. Common aspen and its hybrids (especially
P. tremula ×
P. tremuloides) are frequently planted in Nordic and Baltic countries [
29,
30,
31] and used for pulp production. The increasing demand of woody biomass by the energy sector as well as by the forest industry will increase the need for alternative wood sources. Applying “short rotation forestry” based on fast-growing tree species, including those from the
Populus genus, is a promising alternative [
32]. While a number of papers have focused on the biomass of a variety of tree species, nearly no information on the biomass characteristics for common aspen is available and absolutely no works studied its biomass allocation patterns or component traits in terms of ecological conditions or biological aspects. Our review of the European literature focusing on common aspen indicated neglected interest in this species.
Therefore, the main aim of our paper was to quantify total tree biomass and its allocation to components in common aspen at both the tree and stand levels. A further aim of the paper was to quantify foliage traits (especially SLA) and stand canopy status (LAI) with respect to tree or stand size, specifically stem base diameter.
2. Material and Methods
2.1. Site and Stand Description
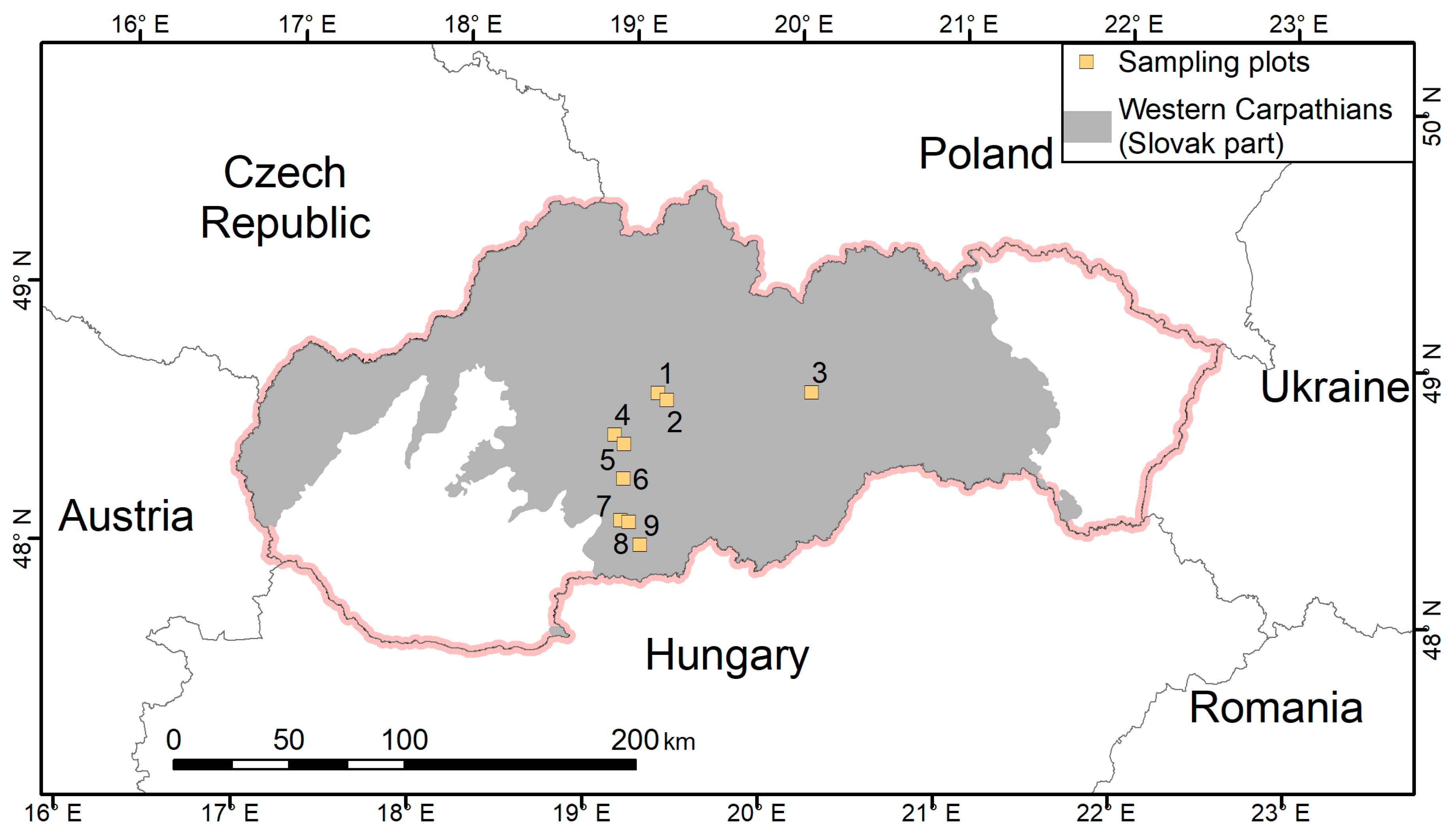
Our research focused on the forested mountainous area of central Slovakia belonging to the Western Carpathians. In general, the forest composition of the Western Carpathians, which is a part of the Carpathian Mountain Range, changes with altitude. At the lowest altitudes, oaks (mainly Quercus robur L. and Q. petraeae (Matt.) Liebl.) dominate, while at the middle altitudes European beech (Fagus sylvatica) is the dominant species. At higher altitudes (over approx. 900 m a.s.l.), coniferous species such as Norway spruce (Picea abies (L.) H. Karst.), Silver fir (Abies alba Mill.), Scots pine (Pinus sylvetris L.), and European larch (Larix decidua Mill.) prevail.
A preliminary selection of forest stands containing common aspen was conducted using a database of Programs of Forest Management by Stand Units in Slovakia (available on:
http://gis.nlcsk.org/lgis/) based on specific information on tree species composition and stand age. The main criteria for forest selection were (1) the share of the target tree species, i.e., aspen, had to be equal to or greater than 90%, and (2) a stand age of maximum 20 years. Afterwards, we examined preselected forest stands in the field, where we checked the origin and the actual contribution of aspen to tree species composition in stands. The final selection of nine forest stands (
Figure 1) was performed considering exclusively natural regeneration and nearly a 100% share of aspen. The youngest selected stand was 2 years old and the oldest stand was 15 years old. All stands were dense with no large gaps in the forest canopy.
The altitudinal range of the selected stands was between 335 m and 610 m a.s.l. (
Table 1, i.e., they occurred in the forest vegetation zone with natural prevalence of oak and beech). In the selected region and altitudes, annual mean temperatures are between 5.0 °C and 8.5 °C, annual precipitation totals fluctuate between 600 and 900 mm, and the growing season usually lasts from 130 to 175 days. Soils are represented by Cambisols, Luvisols, and Rendzina (
Table 1).
2.2. Field and Laboratory Works
Field measurements and tree samplings were performed in the second half of the growing season of 2018. In each selected stand, three circular plots were established with a distance between the plots of at least 10 m. The radius of the plots varied between 1.5 m and 3.0 m, as it depended on stand density and was chosen to cover at least 30 trees. Diameters at stem base (hereinafter as
d0) and tree heights were measured for all trees at the plots. In total, 27 circular plots and 971 trees were measured. Mean tree and stand characteristics for the nine forest stands are presented in
Table 2. Plot-level data were used for subsequent calculations at a stand level using input data recorded at the tree level (destructive tree sampling).
Afterwards, 20 aspen trees were selected from each stand (altogether 180 individuals based on stratified random sampling) to cover all bio-sociological classes (i.e., dominant, co-dominant, sub-dominant, and suppressed). The root system of each tree was excavated to include all roots with a diameter of at least 1 mm. In addition, 5 leaves were sampled from the lower, middle, and upper thirds of the tree crowns, i.e., 15 leaves were sampled from each tree. Leaf subsamples were packed in paper envelopes identified with plot and tree codes, and the position of the leaves in the crown.
Excavated trees were cross-cut at a stem base to separate the belowground (root system) and aboveground parts. Subsequently, branches with foliage were cut off from the main stem. Stem length (tree height) and diameter
d0 were measured. Each tree component, i.e., root system, stems, and branches with foliage, were packed in paper bags identified with plot and tree codes. All samples were transported to the laboratory. Subsamples of leaves were scanned and their one-side leaf surface area (LA hereinafter) was calculated using the Easy Leaf Area software [
33]. Each leaf was oven-dried (under 95 °C for 24 h) and weighed with a precision of 0.001 g. Individual leaf mass and area were used for the calculation of SLA at a specific crown part (upper, middle, and lower).
Leaves were manually trimmed back from the branches. Consequently, roots, stems, branches, and foliage were stored in a warm (about 28 °C), dry, and well-ventilated room. After several weeks, individual tree components were placed in the oven and dried under a temperature of 95 °C for a determined period; specifically, foliage for 48 h and woody parts (i.e., roots, stems, and branches) for 72 h to obtain dry matter. After drying, all components were weighed with a precision of 0.1 g.
2.3. Calculations and Modelling
From the data we derived models that quantified the biomass in the foliage and woody parts of aspen trees, as well as other variables representing leaf growth (LA, SLA, LAI, LAR, LMR). Models were derived for several levels:
- -
The level of individual leaves (leaf mass wf, LA and SLA)
- -
The tree level (foliage mass wf, mass of woody parts wwp, LA, LAR, LMR)
- -
The stand level (foliage mass wf, mass of woody parts wwp, LAI).
An allometric function with two regression coefficients (
b1 and
b2),
was used to describe the relationship of the particular dependent variable (represented by Y) to the
d0 stem base diameter. In the models for the leaf and tree levels, the tree stem base diameter
d0 was used, while in the stand level models a mean stand diameter at the stem base
d0g was used. The frequently used diameter at breast height was not applicable in our analyses, because some of the selected trees did not reach a height of 1.3 m. Almost all constructed equations were derived in a basic power form. The equations of foliage and wood biomass were derived as linearised logarithmic allometric equations that were subsequently transformed back to the exponential form. This approach was applied to ensure a methodological link to previous works that quantified the biomass of components in young trees [
34]. The shape of the allometric function after the logarithmic transformation and its back transformation is
where
λ is a correction factor.
A logarithmic version of the equation is frequently used to exclude heteroscedasticity of residues, which is always present in the calculation of biomass components. Using a logarithmic equation enables the estimation of parameters with linear regressions, which fulfils the assumption of constant variance of residues. Although in the last years non-linear regression methods have been developing, the opinions on these two methodological approaches differ between papers, e.g., [
35,
36,
37].
Models at a level of individual leaves were derived for leaves at different crown parts (upper, middle, and lower) using measured data of
d0 (mm),
wf (g), and
LA.
SLA was calculated using the following formula
Since only
d0 diameter and
wf and
wwp mass were measured for a tree, leaf area
LA of the whole tree had to be calculated otherwise. In our work we derived a new model from the measured foliage mass of a particular tree and a tree
LA calculated as an arithmetical average from the LA values of nine leaves taken from three crown parts of the particular tree using the equation
where:
LA—leaf area of a tree (cm2);
LAi—leaf area of ith sampled leaf (cm2);
wf—mass of all tree leaves (g);
—mass of ith sampled leaf (g);
n—number of leaves sampled from one tree (i.e., 15).
For the models at a stand level, mean stand diameter
d0g (mm) was calculated as a quadratic mean of all tree diameters using the formula
Leaf area index
LAI was calculated as the
LAI under the assumed full canopy coverage (
LAI100%) using the relationship
where:
LAi—leaf area of ith tree at a plot (m2 m−2);
S—plot area in m2;
C—crown canopy coverage at a plot in %;
n—number of trees at a plot.
Similarly, variables in other models (volume of woody parts, and foliage volume) were also converted to 100% crown canopy.
Finally, a generalised linear mixed model (GLMM) was created to evaluate the combined influence of
d0 and position of the leaves in the crown on leaf traits. The model has the following form:
where:
y—a dependent variable, namely leaf mass, leaf area, or specific leaf area;
d0—diameter at stem base;
M—a dummy variable that represents the middle crown part, i.e., if the leaf was taken from the middle part, the value is 1, otherwise it is 0;
U—a dummy variable that represents the upper crown part;
α, β, γ1 and γ2—the regression coefficients of the model.
All statistical analyses were performed in Statistica 10.0 and R software version 3.5.1 [
38].
3. Results
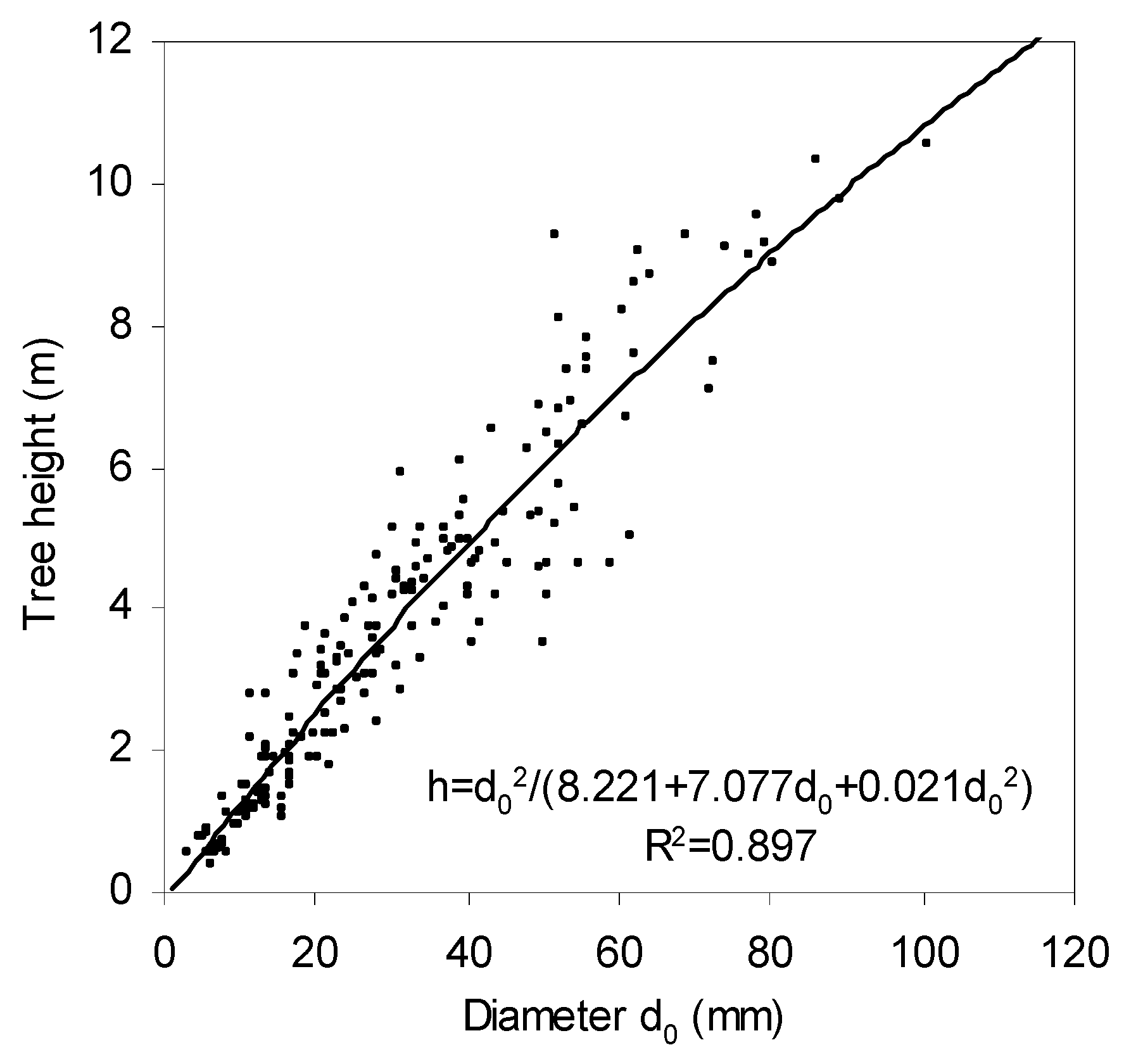
Heights and stem base diameters of the sampled trees varied between 0.4 m and 10.5 m and from 3.3 mm to 100.9 mm, respectively (
Figure 2;
Table 3). The relationship between
d0 diameter and tree height was best described by a fractional power relationship, specifically,
, which explained almost 90% of the variability. This relationship may help users who prefer using tree height as an independent variable in biomass models, to convert our models that are based on
d0 diameter.
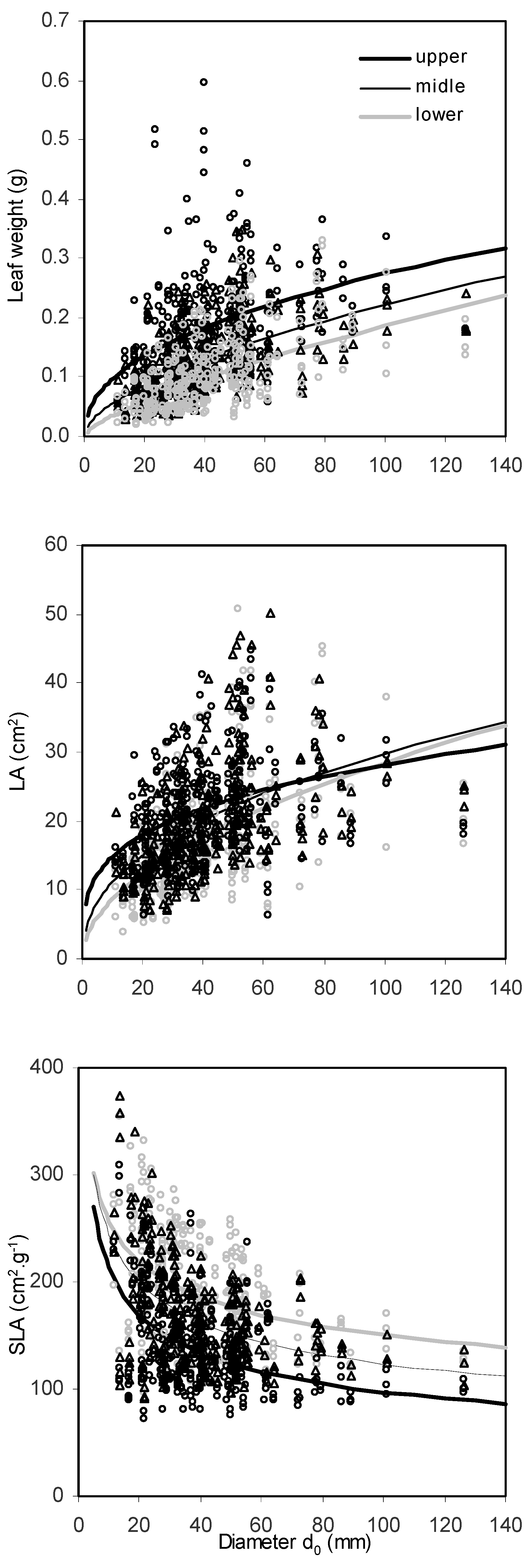
The values of the measured leaves revealed high variation in all the assessed leaf characteristics, i.e., leaf mass (from 0.019 to 0.513 g), leaf area (between 3.79 and 50.81 cm
2) as well as SLA (71.33 to 374.24 cm
2 g
−1;
Table 4). Leaf mass of individual leaves differed along the vertical crown profile, with the heaviest leaves in the upper crown part and the lightest ones in the bottom part of the crown (
Figure 3a). At the same time, leaf area increased with tree size as represented by
d0 diameter (
Figure 3b), although differences between the crown parts were less evident than in leaf mass. The influence of leaf position in the crown and tree size on SLA was very evident (
Figure 3c). The largest SLA values were found for small trees and lower crown parts. Moreover, differences in SLA between the crown parts increased with tree size.
Our mixed models (
Table 5) also revealed a significant influence of the crown part from which the leaves were sampled. The models confirmed that both leaf mass and leaf area increase from the bottom to the top of the crown, i.e., leaves in the upper crown part have larger values of area and mass than in the middle part, which has larger and heavier leaves than the lower part. In the case of specific leaf area, we found an opposite order, i.e., lower > middle > upper, which means that the specific leaf area of the bottom leaves is greater than the SLA of the middle and upper ones (leaves on the top of the crown can be considered as those with minimum SLA, given the diameter at the base is equal for the samples).
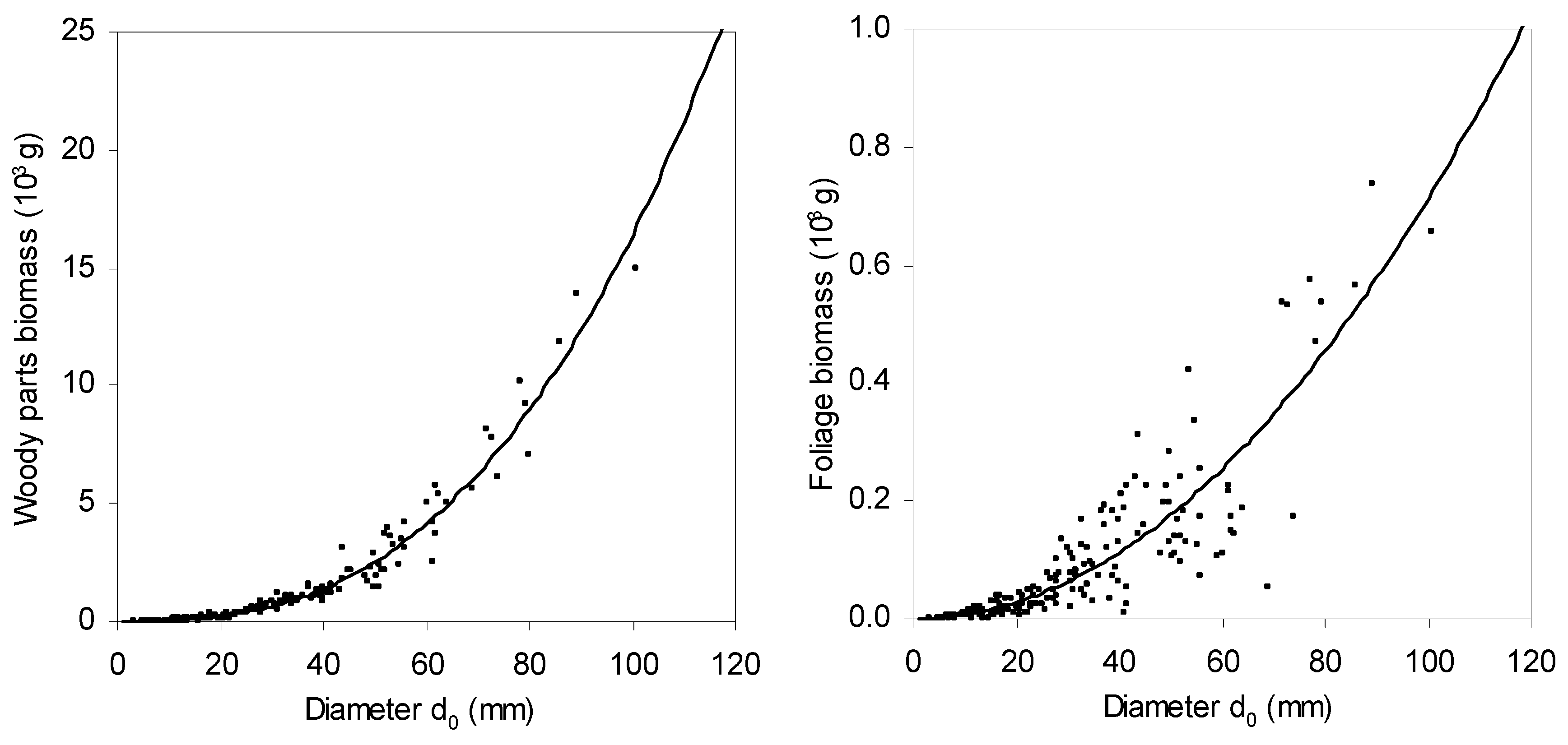
Allometric biomass models for individual tree components as well as for whole-tree biomass showed their close dependences on
d0 diameter (in all cases
p < 0.001; see
Table 6). Fitted values showed that while aspen trees with a diameter of 50 mm had about 2.5 kg of woody biomass and 0.2 kg of foliage biomass, individuals with a diameter of 100 mm had 16.4 kg of biomass in woody parts and 0.7 kg in foliage (
Figure 4). This means that biomass in woody parts increased with tree size faster (in the
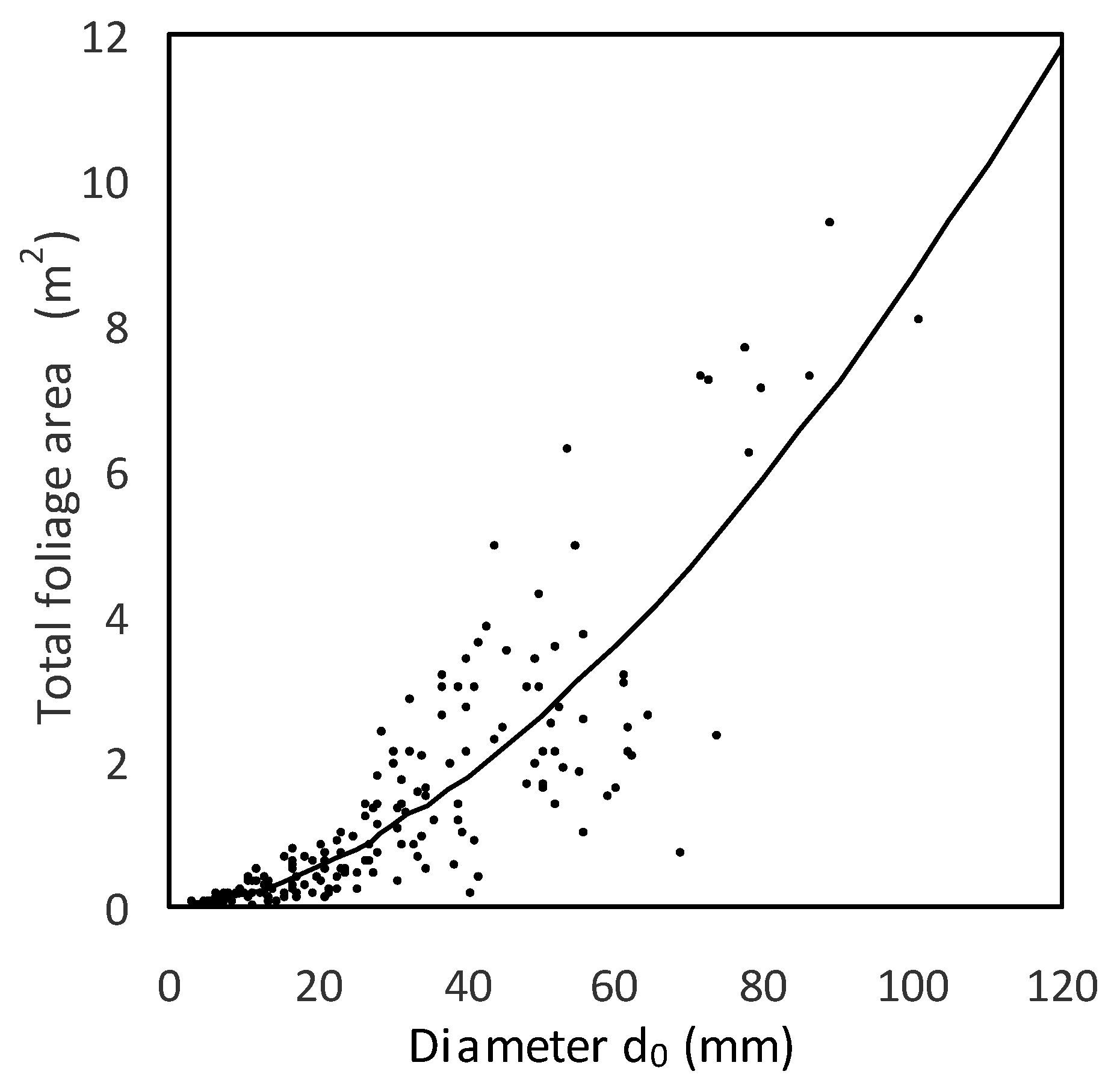
d0 interval 50–100 mm as much as 6.6 times) than in foliage (in the diameter interval 50–100 mm only by 3.5 times). If foliage area at a tree level is considered, increase in the
d0 interval from 50 mm to 100 mm was from 2.6 m
2 to 8.7 m
2 (tripled value;
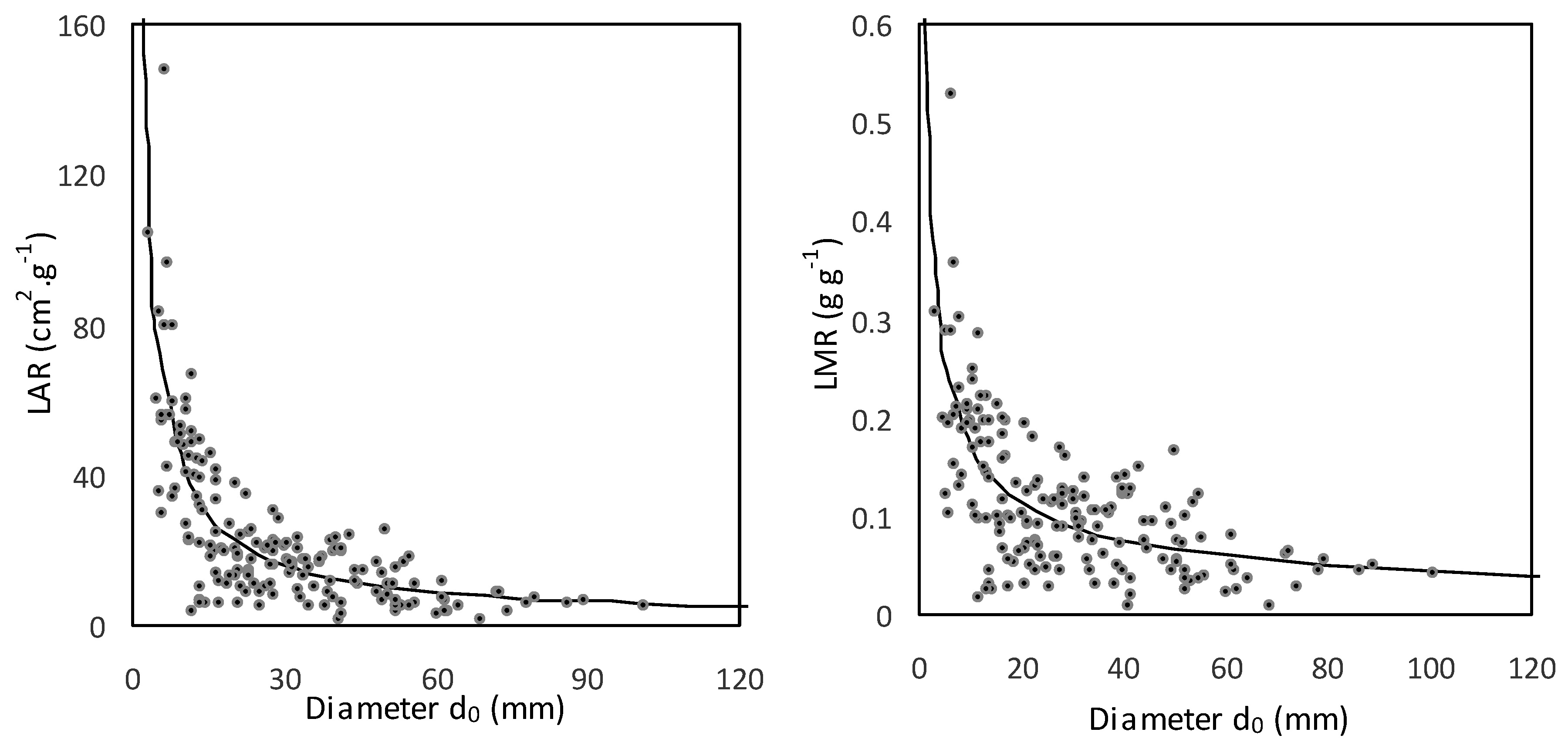
Figure 5). As for LAR and LMR, both indicators decreased with tree size (
Figure 6;
Table 7). This fast decrease was typical in very small trees (with
d0 below approximately 20 mm); then the trend became milder.
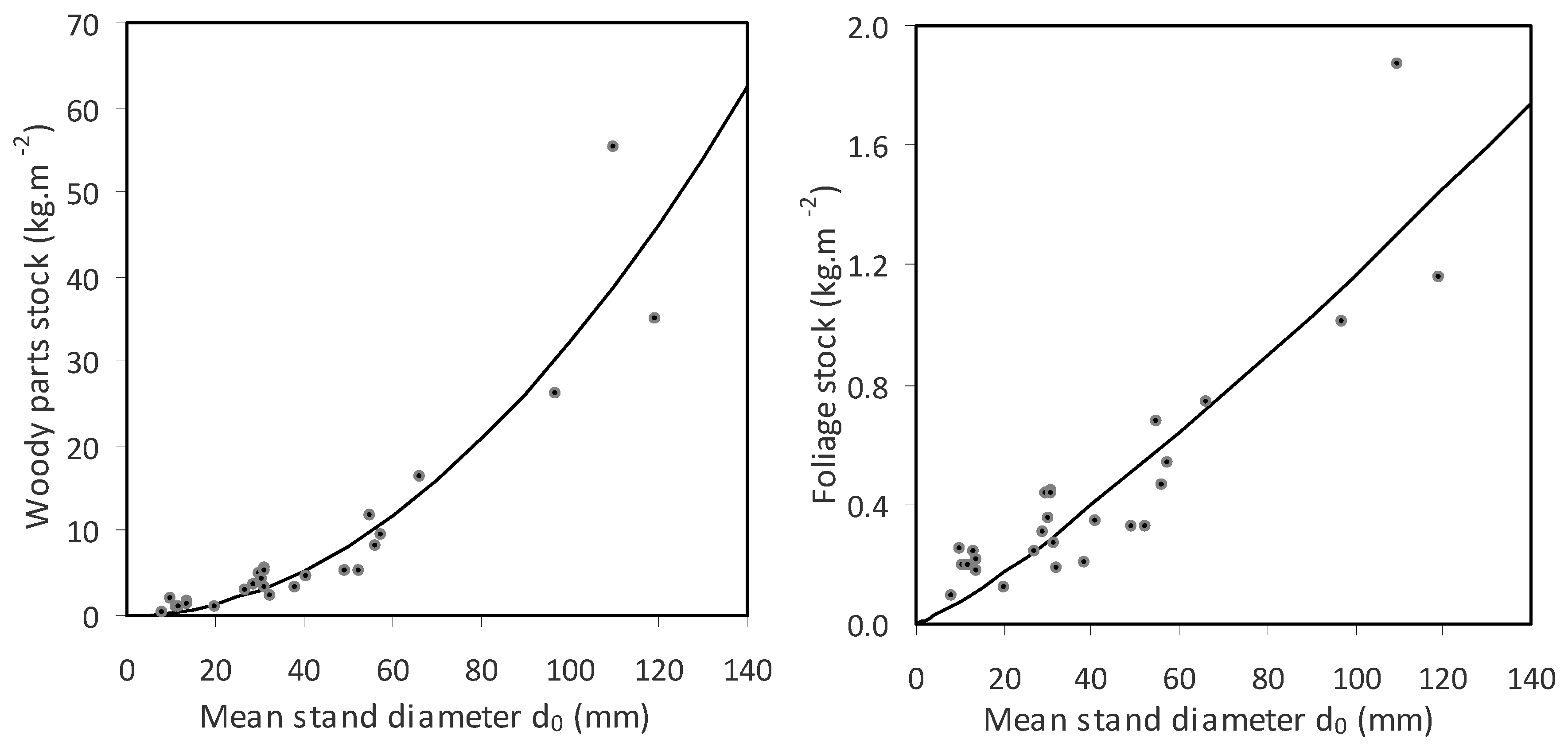
Finally, quantities of woody parts and foliage were estimated at the stand level. The fitted curves (
Figure 7 and
Table 8) showed that the increase in stand biomass in woody parts with an increasing
d0 was greater than the increase in foliage biomass. For instance, while an aspen stand with a mean diameter
d0 of 50 mm had about 8 kg of biomass in woody parts per m
2, a stand with a
d0 of 100 mm contained about 32 kg of woody biomass per m
2. This represents a fourfold increase. On the other hand, the difference between the aspen stands with mean diameters
d0 of 50 mm and 100 mm was only doubled (0.5 kg m
−2 versus 1.2 kg m
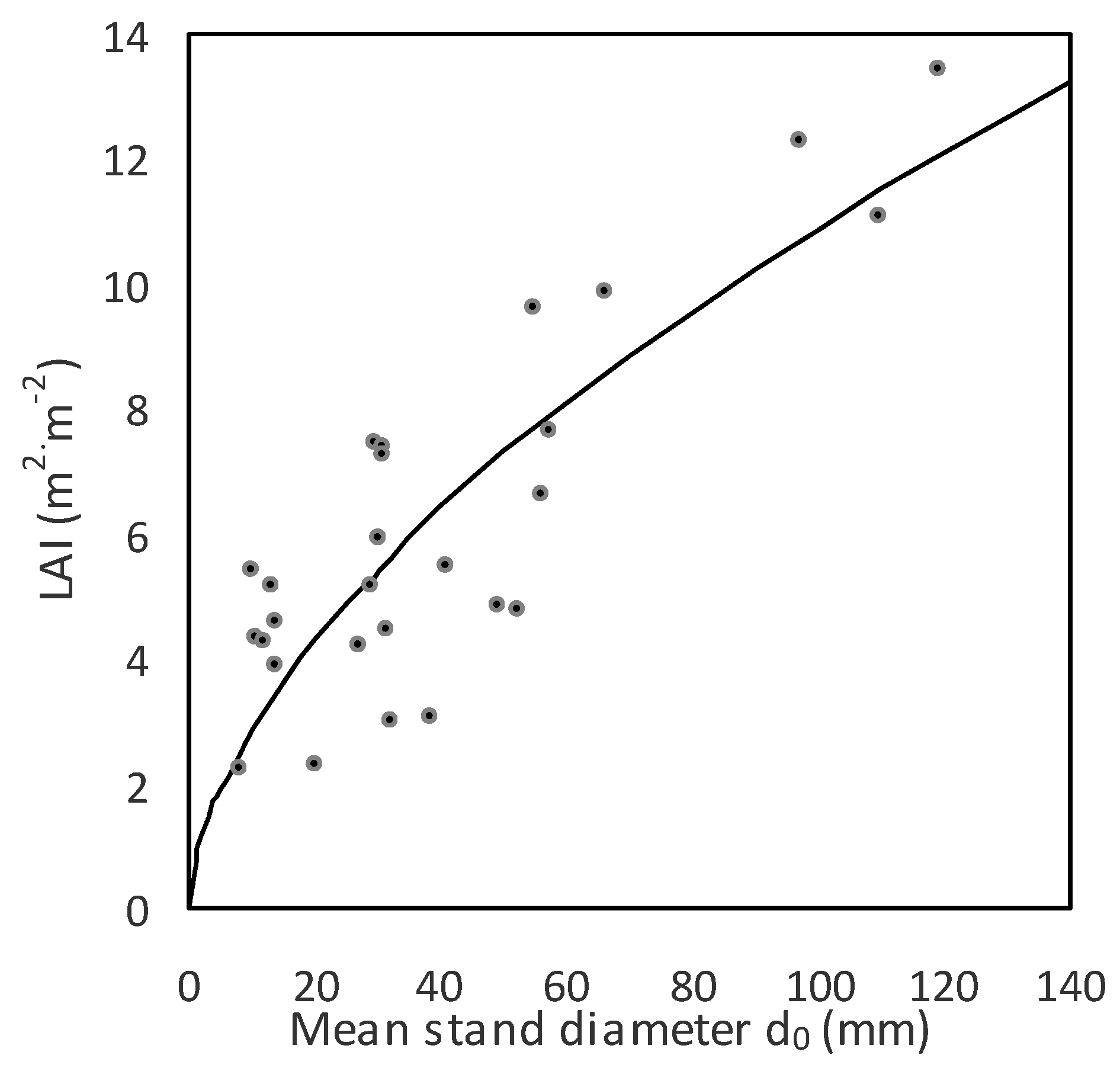
−2). Values of LAI increased with
d0 first fast, and then more slowly (
Figure 8,
Table 8). For instance, while a stand with
d0 equal to 50 mm had an LAI of 7.3 m
2 m
−2, the LAI of a stand with a
d0 of 100 mm was 10.9 m
2 m
−2. The greatest value of LAI equal to 13.5 was found in the aspen forest with a mean stand diameter
d0 of 120 mm.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}