Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions


1
Department of Forest Botany, Dendrology and Geobiocoenology, Faculty of Forestry and Wood Technology, Mendel University in Brno, Zemědělská 1, 613 00 Brno, Czech Republic
2
Department of Silviculture, Faculty of Forestry and Wood Technology, Mendel University in Brno, Zemědělská 1, 613 00 Brno, Czech Republic
*
Author to whom correspondence should be addressed.
Forests 2020, 11(5), 521; https://0-doi-org.brum.beds.ac.uk/10.3390/f11050521
Submission received: 14 April 2020
/
Revised: 1 May 2020
/
Accepted: 4 May 2020
/
Published: 6 May 2020
(This article belongs to the Special Issue Dragon Trees - Tertiary Relicts in Current Reality)
Abstract
:Research highlights: This study is focused on the germination of Dracaena cinnabari seeds in order to discover the possibility of natural and artificial regeneration of this species. Background and Objectives: This study aimed to determine the optimal temperature for D. cinnabari seed germination, e.g., the temperature at which the germination percentage and germination rate (vitality) are the highest. The objectives of this study are to: (1) determine the optimal temperature for the germination of D. cinnabari seeds, (2) compare the suitability of different seed collection methods, and (3) compare the germination parameters of seeds that were collected from different localities. The results of this study will contribute to obtaining the highest number of seedlings from limited seed material for reforestation of the most endangered localities of D. cinnabari species. Materials and methods: Four seed sections were employed. These sections were directly collected from either the fruits of a cut panicle or the ground and were obtained from different localities that differ in altitude. The seeds were tested in a greenhouse while using Petri dishes at three different temperatures—22, 26, and 30 °C—with four replicates of 25 seeds of each section. ANOVA and the t-test were employed for data analysis. Results: The highest germination percentages (GPs) were achieved at 26 °C and 30 °C, which were 84.6% and 82.5%, respectively. The ANOVA and t-test results showed that the germination index (GI) of the species was relatively higher at a temperature of 30 °C relative to that at other temperatures in the study. Although seeds that were collected from the tree achieved a higher GP, the t-test result showed no significant differences in the GI of D. cinnabari seeds that were collected from the ground and from the tree (p > 0.05). Overall, the findings of this study show that temperature has substantial influence on the germination of seeds of D. cinnabari. Therefore, we recommend a temperature of 30 °C to facilitate the germination of D. cinnabari, as it achieved the highest GI at this temperature relative to that at the other temperatures (22 °C, 26 °C) applied in this study.
1. Introduction
The Monocotyledonous Dracaena species, especially the members of the dragon tree group [1], are rare species with a limited and often scattered distribution, very limited natural regeneration, and usually very small populations. Among the different Dracaena species, Dracaena cinnabari Balf.f., which is endemic to Socotra Island, has a large population with an unbalanced age structure [2]. Eighty-thousand individuals are distributed along the eastern part of the island [3,4]; most of them have over mature individuals [5], according to previous studies.
Some of the localities, such as Qatariyah, with the last few trees (localities number 11 and 16 according to Maděra et al. [4]) require urgent intervention to prevent the loss of genetic variability and unique genotype features, as well as other species that are associated with the trees [6,7]. The trees on the whole island do not have a problem of flowering or producing seeds when compared with, e.g., D. serrulata Baker, which flowers, according to observations, mainly after exceptional rain events [8]. D. cinnabari usually flowers each year, at least for the majority of the individuals. Artificial regeneration would be an important practice for supporting natural regeneration, which is lacking due to overgrazing and possibly climate change [9,10,11,12,13].
For successful regeneration, obtaining the highest number of seeds with the highest possible germination percentage is very important. Unfortunately, information regarding the collection of D. cinnabari or Draceana sp. seeds, their handling and processing, and the suitable conditions for germination are lacking [14]. Some researchers reached low germination percentage (35%) of D. cinnabari seeds [15], others described germinated D. cinnabari seeds only after some pre-sowing treatment [16]. These weak germination results prompt the question of whether seeds are dormant or not. Seed dormancy delays or inhibits germination, and some studies, for example, Marrero et Almeida [17] consider non-germinated seeds of D. tamaranae Marrero Rodr., R.S. Almeira and M. Gonzáles-Martin as probably dormant. However, some findings showed a relatively high germination percentage (77%) of D. draco L. seeds [16] or of D. cinnabari seeds (78% and 90%) [18] without any pre-sowing treatment. Using different growth hormones to facilitate seed germination might not also be successful, and some studies, for example Chan-Chin et Govinden-Soulange [19] did not obtain a higher germination percentage of seeds after gibberellic acid and butenolid application for D. concinna Kunth as compared with non-treated seeds. These findings indicate that any pre-sowing treatment might not be necessary.
Seeds must be exposed to suitable conditions to germinate. Temperature is one of the most important external factors for seed germination under natural and laboratory conditions [20,21], which means that a suitable temperature range has to be identified. Some studies confirmed that temperature highly influences the seed germination of different species, including Dracaena sp. For example, Chan-Chin et Govinden-Soulange [19] compared the germination of D. concinna seeds under different temperature conditions (15–35 °C) and obtained the highest germination percentage for a temperature of 25 °C. A high germination percentage have been reached at 20 °C by Hubálková et al. [18] for D. cinnabari seeds, and by Monteiro et al. [22] and Adolt [16] for D. draco seeds. Conversely, the results of Adolt [16] and Chan-Chin et Govinden-Soulange [19] show a low germination percentage of D. cinnabari and D. concinna at the same temperature of 20 °C. None of the seeds of D. concinna germinated at 15 °C [19], and none of the seeds of D. draco germinated at alternating temperatures of 10 °C and 20 °C [22].
These and other similar studies show that each species requires a different temperature for the seeds to germinate. D. cinnabari belongs to the tropical species, for which 25–35 °C would be the optimum temperature for germination [23]. Therefore, studies on the impact of various temperatures on the germination of D. cinnabari are lacking. The results of previous studies are hardly ever comparable, not only due to the use of other Dracaena species but also to the different origins, times, and methods of collection; other methods of handling, processing, and storage of seeds; and, different media for the germination test.
As a result, this study has focused on determining the optimal temperature for D. cinnabari seed germination, i.e., a temperature at which the germination rate, germination percentage, and germination index are the highest. The objectives of this study are to: (1) determine the optimal temperature for the germination of D. cinnabari seeds, (2) evaluate the effect of seed collection methods on the germination of D. cinnabari, and (3) compare the germination parameters of seeds of different origin. All of the results will contribute to obtaining the most seedlings from limited seed material from the most endangered localities of D. cinnabari.
2. Materials and Methods
2.1. Seed Collection
The seeds were collected in three different localities by two different methods.
(1) SB—the seeds were collected from the ground under 10 nearby trees (area of about 0.3 ha) in the Firmihin locality. Seeds without fruit pulp were collected from the ground in October 2014. The Firmihin locality has an elevation of approximately 650 m a.s.l. and is characterised by a mean annual temperature of 23.6 °C with a maximum daily mean of 27.4 °C at the end of April and beginning of May, as well as a decrease in the minimum daily mean of 21.4 °C at the end of January and beginning of February according to data from the Czech climatic station in the Firmihin locality at an altitude of 440 m a.s.l. between 2000 and 2009.
(2) PL—seeds were directly collected from one tree by cutting one panicle of ripe fruits in Firmihin in the same locality as SB seeds in October 2014. Whole fruits (Figure 1a) were taken to the laboratory, and the seeds (Figure 1b) were removed from the fruit. We also counted the total number of fruits in this panicle and the number of seeds in individual fruits (Figure 2a).
(3) NN—seeds were collected directly from trees by picking individual ripe fruits (area of about 0.7 ha) in the Sirhin locality at an elevation of 850 m a.s.l. in September 2014. The mean annual temperature at the climate station with a similar altitude reaches 22.0 °C with a daily mean maximum temperature of 22.6 °C and a daily mean minimum temperature of 18.9 °C, according to our data. The fruits were carefully dried to make them lose their moisture for the purpose of reducing fungi attack and to increase their storability, and then, the seeds were removed.
(4) PE—seeds were collected directly from trees by cutting more panicles (area of about 0.2 ha) in the Skant locality at an elevation of 1400 m a.s.l. in December 2014. The mean annual temperature from the climate station in Skant at an altitude of 1440 m a.s.l. reaches 17.5 °C with a daily mean maximum temperature of 22.2 °C and a daily mean minimum temperature of 12.6 °C, according to our data. The seeds were removed from the fruits a few days after the collection and then taken to the laboratory.
All of the seeds were placed in plastic bags and stored in the laboratory at ordinary (not controlled) temperatures, that ranged between 20–25 °C, until the experiment was started.
2.2. Germination Experiment
The germination experiment was performed in a university laboratory in air-conditioned cabinet Climacell 707 EVO at stable and controlled temperatures of 22 °C (October–November 2015), 26 °C (January–February 2015), and 30 °C (November–December 2015). Each of the experimental studies was performed at different temperatures for 33 days. The seeds were germinated in Petri dishes while using pieces of cotton swabs as a substrate (Figure 2b), both at the bottom of the dish and the top of the dish. The filter paper seemed to be an insufficient substrate, as it did not provide a sufficient amount of moisture to the seeds (due to the round shape and size of the seeds). Four replications were employed for each treatment (i.e., each seed section and temperature); each replication had 25 seeds (i.e., total of 100 seeds). Unfortunately, for the PE locality, a seed germination study at 26 °C was not performed, as the seeds were collected later than other seeds. A total of 400 seeds were employed for the germination study at temperatures of 22 °C and 30 °C, while 300 seeds were employed for the 26 °C temperature germination study.
The seeds were watered and counted at least three times per week (usually on Monday, Wednesday, and Friday), and germinating seeds (Figure 2b) were removed. Only distilled water was applied to retain moisture in the seeds. After the experiment, the ungerminated seeds at 26 °C underwent a Tetrazolium Chloride test to realise the viability of the seeds.
2.3. Data Analysis
For data analysis, the statistical software program SigmaPlot 13 (Systat Software, Inc., San Jose, CA, USA) was utilised. In the data analysis, a parametric method of analysis was performed, as all the germination data were normally distributed. One-way ANOVA was conducted for a comparison of the final mean seed germination percentage (GP), mean germination time (MGT), and mean germination index (GI) of the seeds collected from the different localities and evaluated for their germination at different temperatures. The t-test was also performed for the data analysis. When a significant difference was observed, mean separation was performed while using Fisher’s least significant difference (LSD) test with a significance level of p = 0.05.
GP, MGT, and GI for the different temperature treatments applied to evaluate the seed germination of D. cinnabari seeds were analysed while using the following formulas:
germination percentage (GP) was calculated using the formula
GP (%) = total number of seeds germinated/total number of sown seeds × 100
The mean germination time (MGT) was calculated using the formula
where—n = no. of newly germinated seeds, T = time from the beginning of the experiment
MGT (days) = ∑(T1 × n1 + T2 × n2 +…+ Tk × nk)/∑(n1 + n2 +…+ nk)
Germination Index (GI) was calculated while using the formula
where—N = no. of newly germinated seeds on the first, second, and subsequent days and T = the weight given to the number of germinated seeds on the first, second, and subsequent days. In the GI, the maximum weight is given to seeds germinated on the first day and less to those that were germinated at subsequent dates [24].
GI = ∑(N1 × T1 + N2 × T2 +…+ Nk × Tk)
3. Results
3.1. Number of Seeds in Individual Fruits
We counted the number of fruits and seeds in individual fruits, as we achieved one whole panicle cut from a tree at Firmihin. The panicle contained 427 fruits: 92 fruits contained one seed, 194 fruits contained two seeds, and 141 fruits contained three seeds (Figure 2a). Therefore, the panicle contained a total of 903 seeds. Thus, approximately 21.5%, 45.5%, and 33% of the fruits had one seed, two seeds and three seeds, respectively.
3.2. Germination
3.2.1. Effect of Temperature on Seed Germination
Figure 3 presents the results for the GP of the D. cinnabari seeds that were collected from the different localities, which were evaluated at different temperatures for their germination. The ANOVA results showed significant differences among the different temperatures in the GP of the seeds that were collected in the Sirhin (NN) locality (Figure 3, p = 0.001). However, the ANOVA results for the GP of the seeds collected in the Firmihin (PL) locality, and then treated for their germination at temperatures of 22 °C, 26 °C, and 30 °C showed no significant differences (p = 0.087, Figure 3). Similarly, the statistical analysis results revealed no significant differences in the GP of the seeds collected in the Skant (PE) locality, which were treated at a temperature of 22 °C, 26 °C, and 30 °C (p = 0.445, Figure 3).
Figure 4 shows the ANOVA results for the effect of temperature (22 °C, 26 °C, and 30 °C) on the MGT of the D. cinnabari seeds that were collected from the different localities. The statistical analysis results revealed that the temperature applied as a treatment for germination has a significant effect on the MGT of the D. cinnabari seeds collected from the NN locality (p ≤ 0.001, Figure 4). Similarly, the ANOVA results revealed significant differences in the MGT for the seeds collected from the PL locality and treated for their germination at temperatures of 22 °C and 30 °C and temperatures between 26 °C and 30 °C (p ≤ 0.001, Figure 4). The statistical analysis results also indicated significant differences in the MGT of the seeds collected in the PE locality and treated for germination in temperatures between 22 °C and 30 °C (p ≤ 0.01, Figure 4).
Figure 5 presents the results for the effect of different temperatures on the GI of D. cinnabari seeds, which were collected from the different localities (NN, PL and PE). The ANOVA results showed significant differences in the GI of the D. cinnabari seeds that were collected from the NN locality, which were treated for their germination at temperatures of 22 °C, 26 °C, and 30 °C (p ≤ 0.001, Figure 5). The ANOVA results revealed no significant differences in their GI, for the seeds collected in the PL locality and treated for their germination at temperatures of 22 °C and 26 °C (Figure 5), and significant differences in the GI for the treatments between 22 °C and 30 °C and between 26 °C and 30 °C (Figure 5). The statistical analyses result further indicated significant differences in the GI of the treatments between 22 °C and 30 °C.
3.2.2. Effect of Seed Collection Locality on Seed Germination
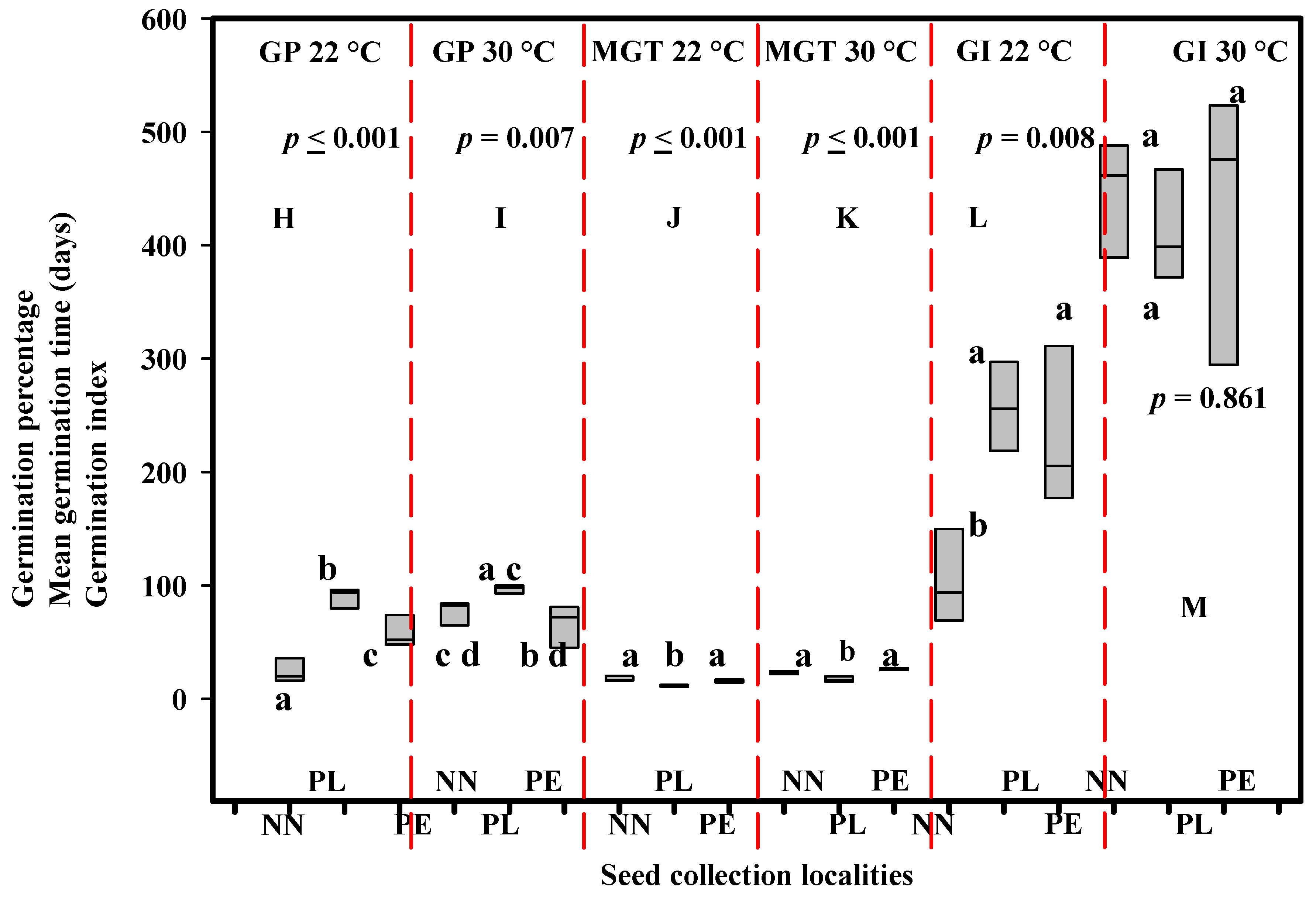
Figure 6 presents the statistical analysis results for the GP, MGT, and GI of D. cinnabari seeds collected from the different localities ((Sirhin (NN), Firmihin (PL), and Skant (PE)), which were evaluated for their germination under a similar temperature condition. The ANOVA results revealed significant differences in the GP of D. cinnabari seeds collected from the NN, PL, and PE localities, which were treated for their germination at temperatures of 22 °C (p ≤ 0.001, Figure 6H) and 30 °C (p = 0.007, Figure 6I). The ANOVA results also indicated significant differences in the MGT of the seeds of D. cinnabari from the localities of NN, PL, and PE, which were treated for their germination at temperatures of 22 °C (p ≤ 0.001, Figure 6J) and 30 °C (p = 0.001, Figure 6K). The ANOVA results showed significant differences in the GI of the D. cinnabari seeds that were collected from different localities and evaluated for their germination at 22 °C (p = 0.008, Figure 6L), while no significant differences in the GI for the seeds of D. cinnabari collected from different localities and assessed for their germination at a temperature of 30 °C (p = 0.861, Figure 6M).
3.2.3. Germination of Seeds Collected from the Ground and Tree Crown
Figure 7, Figure 8, and Figure 9, present the t-test analysis results of the GP, MGT, and GI for seeds of D. cinnabari collected in the Firmihin locality, from the ground (SB) and the tree crown (PL), and subsequently evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C, respectively. The t-test analysis results revealed significant differences in the GP of seeds that were collected from the ground and tree crown and subsequently evaluated for their germination at a temperature of 22 °C (p = 0.04, Figure 7). Similarly, the t-test results showed a significant difference in the GP of seeds collected from the ground and tree crown and then assessed for their germination at temperatures of 26 °C (p = 0.015, Figure 7) and 30 °C (p = 0.020, Figure 7). The results showed no significant differences in the MGT of the seeds that were collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C (p = 0.19, Figure 8), 26 °C (p = 0.996, Figure 8) and 30 °C (p = 0.096, Figure 8). Similarly, the statistical analysis results showed no significant differences in the GI of seeds that were collected from the ground and the tree crown and subsequently evaluated for their germination at temperatures of 22 °C (p = 0.262, Figure 9), 26 °C (p = 0.182, Figure 9), and 30 °C (p = 0.230, Figure 9).
3.3. Viability of the Seeds
The seeds that did not germinate at 26 °C underwent a Tetrazolium Chloride test to determine their viability after finishing the germination experiment. From 300 seeds, 46 seeds did not germinate, of which seven seeds were dead, and the remaining 39 seeds alive. Therefore, the percentage of dead seeds in the whole dataset was only 2.3%, according to this test.
4. Discussion
4.1. Number of Seeds in Individual Fruits
The natural regeneration of Dracaena cinnabari is limited by overgrazing and climate change; therefore, artificial regeneration can help to keep this species in their habitats [13]. Successful regeneration will be influenced by the quantity and quality of seeds and conditions for seed germination and seedlings emergence. The fruit of D. cinnabari is a berry that contains 1–3 seeds. The highest proportion of fruits contained two seeds, while the lowest proportion of fruits contained one seed, based on our results from one panicle cut in Firmihin. This finding contradicts the findings of Adolt [16], who reported the highest proportion of one-seeded fruits and the lowest proportion of three-seeded fruits, and unpublished results obtained by one of the authors in 2003 with the same ratio (refer to Table 1). For a generalisation of results, it would be necessary to analyse more panicles from more trees in time, but these results show the probable activity of pollinators (insect) within different seasons.
A lower amount of seeds in fruits and panicles can be caused by deficient pollination or other factors, such as adverse weather, attacks by insects and other pests, and deficiencies of carbohydrates, mineral nutrients, and hormones [25]. In some species, the fruits from self-pollinated flowers are more likely to be shed prematurely than are fruits from cross-pollinated flowers, according to these authors. The differences of the present results with other findings could also be associated with differences in the age structure of the trees where the seeds were collected. Madera et al. [4], in their study, showed a big variation in the age structure of D. cinnabari throughout Socotra Island.
4.2. Germination Results
For successful regeneration, it is crucial to assure suitable conditions for seed germination. In nature, the germinated seeds of D. cinnabari are often digested by birds, probably endemic Onychognathus frater [26], which can improve the germinating parameters of seeds [27]. Producers of ornamental species recommend soaking D. cinnabari seeds in water for three to five days, which can increase the vitality of the seeds [28]. Furthermore, Adolt [16] worked with D. cinnabari seeds with minimal ability to germinate and increased their GP by soaking in vinegar or hydrogen peroxide. In his experiment, Beyhl [15] achieved a 35% GP, but did not provide any details regarding the conditions, locality, or seed collection methods, and only provided information about the substrate, which was soil. Marrero et Almeida [16] described 10–16% of tested D. tamaranae seeds as dormant. From these publications, we can deduce that D. cinnabari seeds may need some pre-sowing treatment or some pre-sowing treatment could increase their germination parameters. However, as our study and a study by Hubálková et al. [18] indicate, D. cinnabari seeds do not demand any pre-sowing treatment. The differences between the two findings are mainly in MGT, their seeds of D. cinnabari germinated after 2–3 months’ period [18] as compared to 33 days within our experiment. The results of Adolt [16] confirm acceptable results of GP (77%) without pre-sowing treatment and for germination also using whole fruits for D. draco from Tenerife Island.
The substrate properties can also influence the success of germination. E.g., Marrero et Almeida [17] germinated D. tamaranae in a more nutritious soil substrate and achieved GP of 45–96%, while GP in a less nutritious substrate did not exceed 62%. On the other hand, light conditions were not detected to have an influence within Chan-Chin et Govinden-Soulange [19] experiment. They did not observe differences in the germination of D. concinna seeds between dark conditions and diurnal light conditions.
The quality of seeds depends, among others, on the method of seed collection. Our results showed certain differences between seeds that were collected from the ground and seeds removed from the tree (panicle of ripe fruits). Although the seed vitality (MGT and GI) was comparable, the GP of seeds that were collected from trees was higher (92%) than those collected from the ground (79%). This result could be associated with the conditions in the soil and the presence of fungi and microorganisms, which can obviously negatively affect seeds. Collecting seeds from standing lively trees with fruits seems better and more favourable, but the results should be confirmed by some repeated research to analyse seeds from more trees and verify diversity within a population. Moreover, some authors, e.g., Hubálková et al. [18], reached an acceptable germination percentage (84%) for seeds that were collected from the ground and, on the contrary, Adolt [16] germinated seeds from a panicle cut from a tree in Dixam locality reaching zero germination percentage in the case of no pre-sowing treatment. He stated that one of the possible reasons could be a collection of non-ripe fruits, while the collection of seeds from the ground provides a certainty of ripe seeds.
GI appears to be the most comprehensive measurement parameter that combines both the germination percentage and speed (spread, duration and ‘high/low’ events) [29]. The findings of this study revealed that the GI of seeds collected from different localities and the GI of seeds collected from the ground and directly from the tree crown were relatively higher at a temperature of 30 °C than at temperatures of 22 °C and 26 °C in the germination experiment study. This finding shows that temperature has a substantial influence on the germination of seeds of D. cinnabari; the results of a previous study [22] indicate similar findings for Dracaena sp. seeds. Yet, Chan-Chin et Govinden-Soulange [19] obtained the best results for the germination of D. concinna seeds at 25 °C (92% of GP); seeds that germinated at 30 °C and 35 °C had a lower GP (80%). Moreover, Monteiro et al. [22] germinated D. draco seeds in Petri dishes with: (1) a constant temperature of 20 °C and (2) combined temperatures of 10 °C and 20 °C changed every 12 h. Their results show 91% GP for the constant temperature and 0% for the combined temperature. However, GP do not give us a complex idea about seed germination, as written above. The GP of the seeds in our study in two of the tree localities (PL and PE) did not differ at the study temperatures, but the MGT of the seeds for all of the variants was the highest at a temperature of 30 °C. Temperature does not always influence the GP of D. cinnabari seeds, but seeds at different temperatures differ in the speed of germination. This fact agrees with the results of the experiment of Hubálková et al. [18], who reached similar GP but very different MGT. Germination parameters are usually under strong genetic control, which has been confirmed, e.g., at Pseudotsuga menziesii [30], Eucalyptus globulus [31], or other trees species [32], but our study based on seeds from three different localities show no differences in results (i.e., the seeds from all the localities had the best MGT and GI at 30 °C), therefore an influence of maternal effects seems minimal. This is also a reason why we consider the diversity within the populations to be sufficiently covered.
Quite interesting are the results of different germination percentage (GP) and vitality (MGT and GI) at different temperatures of D. cinnabari seeds with different origins. Seeds from the lowest altitude (Firmihin locality, 650 m a.s.l., PL) with the highest mean annual temperature reach the highest GP. Conversely, seeds from the highest altitude (Skant locality, 1400 m a.s.l., PE) with the lowest mean annual temperature reach a lower GP. Hubálková et al. present similar results [18] for the same localities. We discovered that seeds from the lowest altitude (Firmihin locality PL) with the highest mean annual temperature reach the lowest vitality (MGT), and vice versa. A larger number of seeds from mild climates can germinate, but they germinate more slowly than seeds in harsh climates. However, these results should be confirmed by the subsequent research within time to eliminate maternal effects and assure diversity within the population at each locality.
Our results combined with information from previous studies extend knowledge regarding threatened D. cinnabari species; they bring information beneficial for D. cinnabari regeneration. The results can also serve as a basis of further studies that can help to preserve populations of this flagship species.
5. Conclusions
Overall, the present study results could show that, despite different authors who have carried out different germination studies on D. cinnabari and found variable results, the current findings showed that the 30 °C temperature used as a treatment enhanced the seed germination percentage in a shorter period of time.
D. cinnabari seeds can reach a lower GP when the seeds are collected from the ground than from the tree. Therefore, we recommend collecting ripe fruits from standing trees, followed by seed extraction. A pre-sowing treatment to break seed dormancy for the species is not necessary. Rather, temperature highly influences the germination of seeds of D. cinnabari. This study showed that a temperature of 30 °C applied as a treatment for germination achieved a higher GI relative to the other temperatures (22 °C and 26 °C) in the study. Therefore, we recommend applying this temperature to enhance the germination of the species.
Author Contributions
L.B., field data harvesting, data analysis, and writing—original draft preparation; S.A.M., methodology, data analysis, and writing—original draft preparation; K.H., field data harvesting, and writing—original draft preparation; H.H., conceptualisation, methodology, and writing—original draft preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Czech Development Agency, grant number 22/2015/10.
Acknowledgments
The authors are grateful to the Environment Protection Authority, Socotra Branch who arranged us the permission on seed collection. The authors are also grateful to Pavel Hanáček and his team, who supported us in providing an air-conditioned cabinet for the experiment, to Petr Němec, who collected seeds in Skant, and Abdulwahab Saad who collected seeds in Sirhin.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marrero, A.; Almeida, S.R.; Martín-González, M. A new species of the wild Dragon Tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic Implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Adolt, R.; Pavliš, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees-Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svrátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon’s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Čermák, Z.; Pavliš, J. Dragon’s blood tree—Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- Vasconcelos, R.; Pujol-Buxó, E.; Llorente, G.A.; Saeed, A.; Carranza, S. Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests 2020, 11, 353. [Google Scholar] [CrossRef] [Green Version]
- Vahalík, P.; Patočka, Z.; Drápela, K.; Habrová, H.; Ehrenbergerová, L.; Lengálová, K.; Kalivodová, H.; Pompeiano Vaníčková, L.; Al-Shamahi, E.; Lupton, D.; et al. The Conservation Status and Population Mapping of the Endangered Dracaena serrulata in the Dhofar Mountains, Oman. Forests 2020, 11, 322. [Google Scholar] [CrossRef] [Green Version]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Brown, G.; Mies, B.A. Vegetation Ecology of Socotra. Plant and Vegetation 7, 1st ed.; Springer: Berlin, Germany, 2012; p. 379. [Google Scholar]
- Hubálková, I. Prediction of Dragon’s Blood Tree (Dracaena cinnabari Balf.) stand sample density on Soqotra Island. J. Landsc. Ecol. 2011, 4, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Pavliš, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf. f. of Socotra Island suggest essential elements for a conservation strategy. Biol. Bratisl. 2018, 74, 339–349. [Google Scholar] [CrossRef]
- Maděra, P.; Forrest, A.; Hanáček, P.; Vahalík, P.; Gebauer, R.; Plichta, R.; Jupa, R.; Van Rensburg, J.J.; Morris, M.; Nadezhdina, N.; et al. What We Know and What We Do Not Know about Dragon Trees? Forests 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Beyhl, F.E. Attempts at raising the Soqotran Dragon tree, Dracaena cinnabari outside the island. In Proceedings of the First International Symposium on Soqotra Island: Present and Future; Dumont, H.J., Ed.; United Nations Publications: New York, NY, USA, 1996; pp. 125–133. [Google Scholar]
- Adolt, R. Proposal of principles of dragon trees gene resources protection in forests on Socotra and Canary Islands. Master’s Thesis, Mendel University of Agriculture and Forestry, Brno, Czech Republic, 2001; p. 247. [Google Scholar]
- Marrero, A.; Almeida, R.S. Aspectos de la biología reproductiva del drago de Gran Canaria: Dracaena tamaranae, en relación a su conservación. Rev. Acad. Canar. Cienc. 2017, 29, 111–202. [Google Scholar]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth dynamics of Dracaena cinnabari under controlled conditions as the most effective way to protect endangered species. Saudi J. Biol. Sci. 2017, 24, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Chan-Chin, D.; Govinden-Soulange, J. Germination profile of selected plants from Mauritius—Towards a conservation strategy. Seed Sci. Technol. 2015, 43, 536–540. [Google Scholar] [CrossRef]
- Black, M.; Bewley, J.D.; Halmer, P. The Encyclopedia of Seeds: Science, Technology and Uses; CAB International: Wallingford, UK, 2006; p. 828. ISBN 0-85199-723-6. [Google Scholar]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CABI Publishing of CAB International: London, UK, 2000; pp. 261–292. [Google Scholar]
- Monteiro, A.; Vasconcelos, T.; Tapada Bertelli, A. Seed propagation of Dracaena draco L. Garcia Orta Sér. Bot. 1999, 14, 187–189. [Google Scholar]
- FloraBank. Guideline 8: Basic Germination and Viability Tests for Native Plant Seed. Australian Tree Seed Centre and Warren Mortlock. 1999. Available online: https://www.greeningaustralia.org.au/publications/florabank-germination-viability/?download (accessed on 20 March 2020).
- Bench, A.R.; Fenner, M.; Edwards, P. Changes in germinability, ABA content and ABA embryonic sensitivity in developing seeds of Sorghum bicolor (L.) Moench induced by water stress during grain filling. N. Phytol. 1991, 118, 339–347. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G. Physiology of Woody Plants, 2nd ed.; Academic Press: Cambridge, MA, USA, 1997; p. 411. [Google Scholar]
- Al-Saghier, O.; Porter, R.F. The Bird Biodiversity of Soqotra. In Proceedings of the First International Symposium on Soqotra Island: Present and Future; Dumont, H.J., Ed.; United Nations Publications: New York, NY, USA, 1998. [Google Scholar]
- Turček, F.J. The relationships of birds and mammals to tree species, with particular focus to seeds as a food resource. The Work of Czechoslovak forestry research institutes (in Slovak language). Práce výzkumných ústavů lesnických 1952, 1, 125–166. [Google Scholar]
- Dracaena Seed Propagation Guide—How to Plant Dracaena Seeds. Available online: https://www.gardeningknowhow.com/houseplants/dracaena/dracaena-seed-propagation.htm (accessed on 28 March 2020).
- Kader, M.A. A Comparison of Seed Germination Calculation Formulae and the Associated Interpretation of Resulting Data. J. Proc. R. Soc. N. South Wales 2005, 138, 65–75. [Google Scholar]
- El-Kassaby, Y.A.; Edwards, D.G.W.; Taylor, D.W. Genetic control on germination parameters in Douglas-fir and its importance for domestication. Silvae Genet. 1992, 41, 48–54. [Google Scholar]
- Rix, K.D.; Gracie, A.J.; Potts, B.M.; Brown, P.H.; Gore, P.L. Genetic control of Eucalyptus globulus seed germination. Ann. For. Sci. 2015, 72, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.; Wingfield, M.J.; Slippers, B. Maternal effects should be considered in the establishment of forestry plantations. For. Ecol. Manag. 2020, 460, 117909. [Google Scholar] [CrossRef]
Figure 1.
Fruits (a) and seeds (b) of Dracaena cinnabari.

Figure 2.
Example of three-seed fruit (a), and germinating seeds on cotton swabs (b).

Figure 3.
ANOVA results for comparison of the effect of temperature on the final germination percentage (%) of seeds collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 3.
ANOVA results for comparison of the effect of temperature on the final germination percentage (%) of seeds collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).

Figure 4.
ANOVA results for comparison of the effect of temperature on the mean germination time (days) of seeds of D. cinnabari collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 4.
ANOVA results for comparison of the effect of temperature on the mean germination time (days) of seeds of D. cinnabari collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).

Figure 5.
ANOVA results for comparison of the effect of temperature on the germination index of seeds collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 5.
ANOVA results for comparison of the effect of temperature on the germination index of seeds collected in the Sirhin (NN), Firmihin (PL), and Skant (PE) localities. Means with the same letter are not significantly different from each other (p = 0.05).

Figure 6.
ANOVA results for comparison of the effects of seed collection localities/sources (Sirhin (NN), Firmihin (PL), and Skant (PE)) on the germination percentage (GP), mean germination time (MGT), and germination index (GI) of D. cinnabari seeds. Means with the same letter are not significantly different from each other (p = 0.05). Figure 6 H and I represent GP at 22 °C and 30 °C temperature, respectively. Figure 6 J and K represent MGT at 22 °C and 30 °C temperature, respectively. Figure 6 L and M represent GI at 22 °C and 30 °C temperature, respectively.
Figure 6.
ANOVA results for comparison of the effects of seed collection localities/sources (Sirhin (NN), Firmihin (PL), and Skant (PE)) on the germination percentage (GP), mean germination time (MGT), and germination index (GI) of D. cinnabari seeds. Means with the same letter are not significantly different from each other (p = 0.05). Figure 6 H and I represent GP at 22 °C and 30 °C temperature, respectively. Figure 6 J and K represent MGT at 22 °C and 30 °C temperature, respectively. Figure 6 L and M represent GI at 22 °C and 30 °C temperature, respectively.

Figure 7.
t-test results for the germination percentage of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 7.
t-test results for the germination percentage of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).

Figure 8.
t-test results for the mean germination time (MGT) of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 8.
t-test results for the mean germination time (MGT) of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).

Figure 9.
t-test results for the germination index (GI) of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).
Figure 9.
t-test results for the germination index (GI) of D. cinnabari seeds collected from the ground and directly from the crown and evaluated for their germination at temperatures of 22 °C, 26 °C, and 30 °C. Means with the same letter are not significantly different from each other (p = 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of fruits with one, two, and three seeds in one panicle.
| Fruits with: | 1 Seed | 2 Seeds | 3 Seeds | Total Fruits/Seeds |
|---|---|---|---|---|
| No. of fruits/seeds 2014 | 92/92 | 194/388 | 141/423 | 427/903 |
| No. of fruits/seeds (Adolt 2001) | 156/156 | 81/162 | 16/48 | 253/366 |
| No. of fruits/seeds 2003 | 699/699 | 141/282 | 30/90 | 870/1071 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bauerová, L.; Munie, S.A.; Houšková, K.; Habrová, H. Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions. Forests 2020, 11, 521. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050521
AMA Style
Bauerová L, Munie SA, Houšková K, Habrová H. Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions. Forests. 2020; 11(5):521. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050521
Chicago/Turabian StyleBauerová, Lucie, Shiferaw Alem Munie, Kateřina Houšková, and Hana Habrová. 2020. "Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions" Forests 11, no. 5: 521. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050521
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.